
Abstract
As an inhibitor of TGF-β signaling, SMAD7 was reported to play dual roles in breast cancer development and progression. It inhibited the cancer metastasis by blocking epithelial–mesenchymal transition, however, litter studies focused on its role in other cancer processes. In this study, miR-497 expression was found inversely correlated with SMAD7 expression in breast cancer tissues. Bioinformatics analyses defined a potential miR-497 response element within 3′ untranslated region of SMAD7 that was validated in reporter gene experiments. Enforced miR-497 expression, accompanied with SMAD7 reduction, suppressed MDA-MB-231 and MCF-7 breast cancer cell growth by MTT and invasion assay, and, induced the S phase arrest detected by flow cytometry. Furthermore, upregulated miR-497 expression by mimics treatment significantly suppressed the tumor growth in the orthotopic nude mouse models. Finally, high expression of miR-497 conferred a better prognosis, indicated by Kaplan–Meier test, especially in HER2 overexpression and triple-negative breast cancer (TNBC). Taken together, our results identified the proliferation promoting role of SMAD7 in breast cancer and therefore established the regulations of SMAD7 in breast cancer by miR-497 through a posttranscriptional mechanism. Moreover, miR-497 might be deemed as a novel potential therapeutic target for the HER2 positive and TNBC in future.
Introduction
C
microRNAs, as a kind of small noncoding RNAs, come down to the adjustment of gene expression through inducing mRNA degradation or repressing mRNA translation (Ke et al., 2003; He and Hannon, 2004). The region of the mature miRNA binding to locates at the mRNA 3′-untranslated region (UTR) of specific protein-coding genes (Di Leva et al., 2014). Complete complementarity between miRNAs and the 3′-UTR of the target gene results in mRNA degradation, and partial complementarity usually leads to the target protein's translation inhibition, for human being usually the latter happens (Volinia et al., 2006). More and more evidence showed that miRNAs played key roles in some human biological and pathological processes, such as cell growth, development, and tumorigenesis (Farazi et al., 2013; Listowski et al., 2013). Previous studies have determined that, in breast cancer, the expression level of miR-497 was higher in normal breast tissues compared with the breast cancer tissues, and forced expression of miR-497 suppressed breast cancer cell proliferation and invasion (Li et al., 2011). Moreover, Shen et al. showed that miR-497 could induce apoptosis of by targeting Bcl-w in MCF-7 breast cancer cells (Shen et al., 2012). Luo et al. indicated that miRNA-497 regulated cell growth and invasion by targeting cyclin E1 in MDA-MB-231 breast cancer cells (Luo et al., 2013b).
SMAD family is the important mediator of TGF-β signaling pathway, which includes three functional classes: common mediator SMADs, receptor-regulated SMADs (R-SMADs), and inhibitory SMADs (I-SMADs) (Reguly and Wrana, 2003). SMAD7, as a member of I-SMADs, was reported to be overexpressed in anaplastic thyroid carcinoma, melanoma, and prostate cancer, and its abundance was positively associated with the tumor malignancy (Luo et al., 2014). As a response to TGF-β, SMAD7 physically interacts with TβRI and inhibits R-SMAD-mediated TGF-β activation. Therefore, SMAD7 functions as an inhibitor of TGF-β signaling (Javelaud and Mauviel, 2004).
Taken together, based on previous studies, we speculated that miR-497 might function as a tumor suppressor in breast cancer. Furthermore, we happened to find a correlation between miR-497 and SMAD7 expressions in breast cancer tissues, and subsequent bioinformatics analysis and luciferase assay confirmed the potential binding sites of miR-497 on SMAD7 3′UTR. Therefore, a regulatory relationship may exist between miR-497 and SMAD7, which will expand the mechanisms of miR-497-mediated breast cancer cell growth suppression and provide more sufficient evidence for selecting miR-497 as a potent therapeutic target in breast cancer.
Materials and Methods
Tissue samples
This project was supported by Tianjin municipal Major Scientific and Technological Special Project for Significant Anticancer Development (No. 12ZCDZSY15400). A total of 45 primary breast cancer tissues and corresponding adjacent normal tissues from patients were collected from the Third Department of Breast Cancer, Tianjin Medical University Cancer Institute and Hospital during the period from 2013 to 2014. Another 240 primary breast cancer tissues were collected during the period from 2008 to 2009. Samples were immediately snap frozen in liquid nitrogen and stored at −80°C until use. Informed consent was obtained from all patients, and the research protocols were approved by the Ethics Committee of Tianjin Medical University Cancer Institute and Hospital.
Cell culture and transfection
Human MDA-MB-231 and MCF-7 breast cancer cells were obtained from the Chinese Science Institute. The cells were cultured in RPMI 1640 (RPMI 1640; CORNING) supplemented with 10% fetal bovine serum (Gibco). Cells were cultivated at 37°C in a humidified chamber supplemented with 5% CO2. Cells were transfected with miR-497 mimics or control by use of Lipofectamine 2000 (Invitrogen). miR-497 mimics and negative control were purchased from RiboBio, and sequences are as follows: miR-497 mimics, 5′-CAGCAGCACACUGUGGUUUGU-3′, negative control, 5′-UUGUACUACACAAAAGUACUG-3′.
Real-time polymerase chain reaction assay
Total RNA extraction was carried out using the Total RNA isolation kit (Bioteke Corporation). For microRNA, we used miRcute miRNA first-strand cDNA synthesis kit and miRcute miRNA quantitative real-time polymerase chain reaction (qPCR) detection kit (SYBR Green) (Tiangen) to detect the levels of miR-497 according to the manufacturer's protocol. U6 was as an internal control. Real-time polymerase chain reaction (PCR) was performed on a CFX96 Real-Time System (Bio-Rad) using the following primers: miR-497 forward: 5′-GTGCAGGGTCCGAGGT-3′, U6 forward, 5′-GCTTCGGCAGCACATATACTAAAAT-3′. For mRNA, we put 2 μg of total RNA reverse transcribed with the BioTeke super RT Kit (Bioteke Corporation) to synthesize cDNA samples. Two milliliter of cDNA product were used for PCR amplification with GoTag qPCR Master Mix (Promega) on a thermal cycler using the following primers. SMAD7 mRNA forward: 5′-ATGCTGTGCCTTCCTCCGCTG-3′. Reverse: 5′-CCACGCACCAGTGTGACCGA-3′. RPS14 mRNA forward: 5′-TCACCGCCCTACACATCAAACT-3′. Reverse: 5′-CTGCGAGTGCTGTCAGAGG-3′. RPS14 was used as an internal control.
Western blot analysis
Cells were harvested and washed in cold PBS twice and then were treated with the lysis buffer on ice for 30 min. The lysate was collected into microtubes and centrifuged for 15 min at 12,000 rpm at 4°C. Protein samples (20 μg) were denatured with the 5 × SDS loading buffer at 100°C for 5 min, then were segregated on a 10% SDS polyacrylamide gel electrophoresis, and transferred onto 0.45 μm nitrocellulose membranes. After 60 min of blocking with 5% fat-free milk, membranes were incubated with SMAD7 antibody (1:2000; Sigma) and β-actin antibody (1:2000; Abmart) overnight at 4°C. Blots were washed for 1 h with the anti-rabbit secondary antibody (1:2000; Cell Signaling Technology). After washing with TBST thrice, immunoreactive protein bands were detected.
Dual luciferase activity assay
The 3′-UTR vector of SMAD7 containing an intact miR-497 recognition sequence was purchased from Kangbio. Forty-eight hours after transfection, the luciferase activity was measured using the Dual Luciferase Reporter assay kit (Promega). Cells were washed twice with cold PBS, and 100 μL passive lysis buffer were added and incubated at room temperature for 30 min. Then, the firefly luminescence (FL) reporter was measured immediately after adding 100 μL luciferase assay reagent II. After that, 100 μL Stop&Glo reagent was added to each well to initiate the renilla luminescence (RL) reporter. The ratio of FL/RL was calculated to indicate the activity of the reporter.
MTT assay
The cells were seeded in a 96-well plate at the density of 4 × 103 cells. Then cells were transfected with 0, 25, 50, 75, and 100 nM miR-497 mimics and cultivated for 24, 48, and 72 h, separately. After that, we put 20 μL MTT (5.0 mg/mL) in each well and incubated for 4 h. The culture media were removed, 200 mL DMSO were added. Optical densities were determined at 490 nm.
Tumor cell invasion assay
After 48 h of transfection with control, mimics, and mimics+ SMAD7 expression plasmid, we plated MDA-MB-231 and MCF-7 cells in a six-well plate, respectively. After incubation for 24 h, we removed the medium and washed cells with PBS twice. The cells that had passed through the Matrigel™ membrane were stained with Crystal Violet solution for 25 min and then counted in eight high-power fields. The results represent the average of three independent experiments.
Cell cycle analysis
Cells transfected with miR-497 mimics or negative control were centrifuged at 1200 rpm for 10 min and washed thrice with cold PBS. Then cells were fixed in 70% ethanol and were washed thrice with cold PBS. After 30-min digestion by RNase (10 mg/mL) at 37°C, 15 μL propidium iodide staining solution (1 mg/mL) was added to each sample and analyzed by a flow cytometer.
Orthotopic mouse models establishment and miR-497 treatment
All mouse experiments were approved by the general principles contained in the Guide for the Care and Use of Laboratory Animals published by Tianjin Medical University Cancer Institute and Hospital. BALB/c-A nude mice at 4 weeks of age were purchased from the Animal Center at the Cancer Institute at Chinese Academy of Medical Science (Beijing, PR China). MDA-MB-231 cells were subcutaneously injected into mammary fat pads of each nude mouse. When the tumor volume reached 150 mm3, mice were randomly divided into two groups. Each group (n = 8) was treated with miR-497 mimics or negative control in 10 μL of lipofectamine through local injection of the xenograft tumor at multiple sites. The treatment was performed once in every 2 days for 20 days. The tumor volume was measured with a caliper, using the following formula: volume = length × width2/2. At the end of the experiment on day 20, the mice were killed, tumor weight was measured, and mice tissues were used for miR-497 and SMAD7 expression assay.
Statistical analysis
The statistical analysis was performed with the use of the software package SPSS version 15.0. Two-tailed Student's t-test was used to assess differences in values between each group. Survival curves were plotted using the Kaplan–Meier method and were evaluated for the statistical significance using a log-rank test. p-value of 0.05 was considered to be statistically significant.
Results
miR-497 is reduced in breast cancer tissues, which has a negative correlation with SMAD7 expression
To detect the expression of miR-497 and SMAD in breast cancer tissues, forty-five pairs of breast cancer tissues and adjacent normal tissues were detected by real-time PCR. As the results, the expression levels of miR-497 in breast cancer tissues, were significantly lower than in corresponding adjacent normal tissues (p = 0.001; Fig. 1A), SMAD mRNAs overexpressed in breast cancer tissues (p = 0.008; Fig. 1B). Furthermore, the expression of miR-497 was significantly inversely correlated with the SMAD7 expression in tissues (R = −0.891, p = 0.01, Fig. 1C).
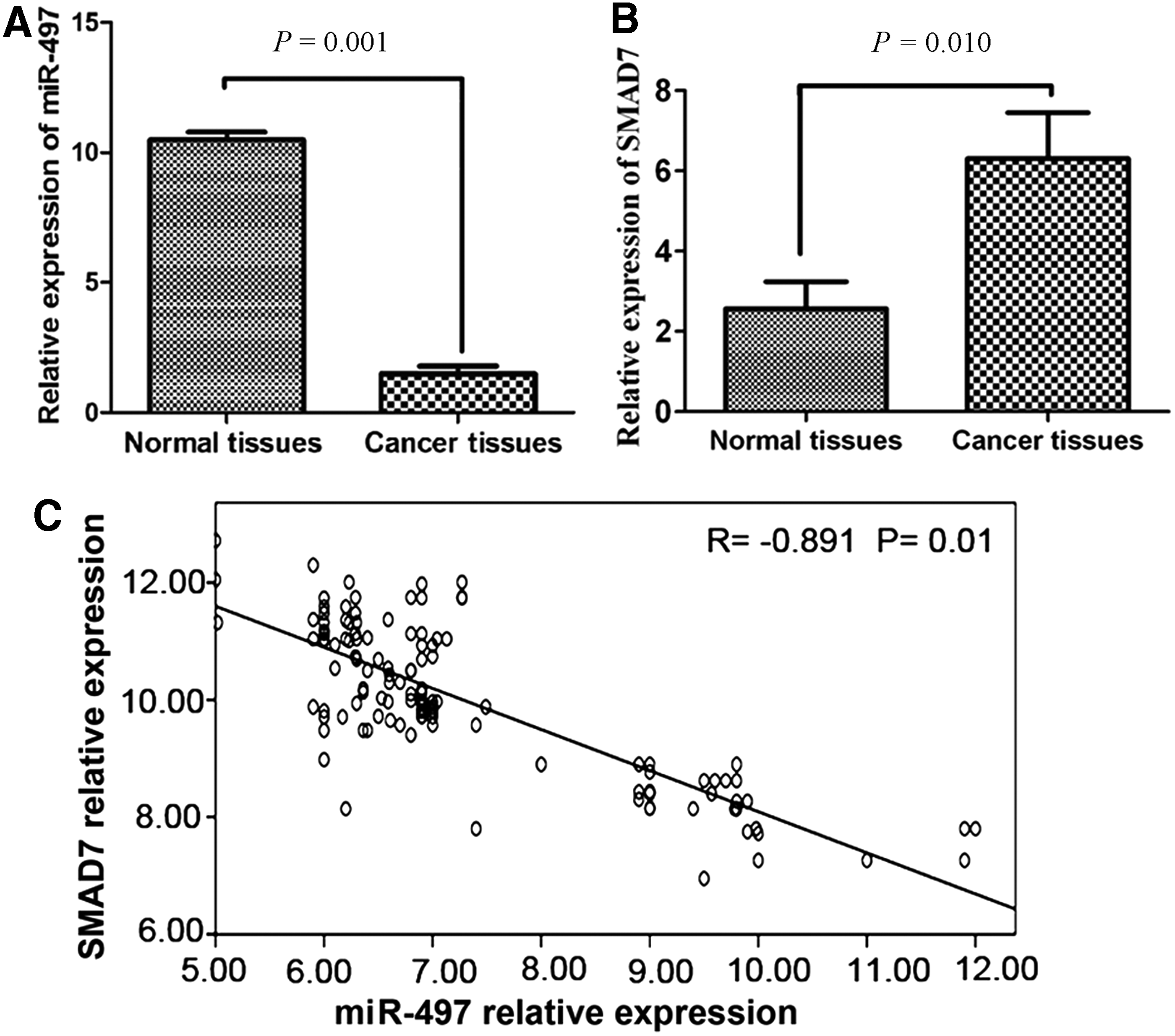
Differential expression of miR-497 and SMAD7 assayed by real-time polymerase chain reaction (PCR) in cancer tissues and adjacent normal tissues of 45 breast cancer samples.
Computational prediction of miR-497 as a potential regulator of SMAD7
miR-497 and SMAD7 showed a negative correlation in expression in breast cancer as described above, however, a regulatory mechanism existed between them that remained uncertain. Following the microRNA target analysis results predicted by targetscan, miRanda, miRGen, and miRBase, miR-497 was identified as a potential regulator of SMAD7 in 3′UTR. Guided by the prediction algorithms, we hypothesized that the “seed sequence” of miR-497 might match the 3′UTR of SMAD7 mRNA (Fig. 2A) and therefore constructed the wild or mutant SMAD7 3′UTR containing luciferase reporter constructs to determine whether SMAD7 expression was indeed regulated by miR-497 (Fig. 2B). Real-time PCR and western blot analysis showed that SMAD7 expression was reduced in MDA-MB-231 and MCF-7 cells upon miR-497 overexpression (Fig. 2C, D). To further confirm SMAD7 was a putative target for miR-497, luciferase reporter construct carrying the 3′UTR miR-497 potential-binding sites or miR-497 mutant-binding sites of SMAD7 was cotransfected with either miR-497 mimics or negative control into the MDA-MB-231 and MCF-7 breast cancer cells. Compared with the control, transfection with miR-497 mimics reduced the luciferase activities significantly. However, the miR-497 mimics did not affect the luciferase activity in the mutant construct when compared with the control (Fig. 2E). These data demonstrated that miR-497 directly modulated SMAD7 expression by binding to the 3′UTR.

SMAD7 is a direct target of miR-497 in breast cancer cells.
miR-497 suppresses breast cancer cell proliferation and invasion in vitro by targeting SMAD7
To explore the tumor suppressive role of miR-497, we reexpressed it in two breast cancer cell lines (MDA-MB-231 and MCF-7) followed by functional assays. Compared with the control group, the cell activity was repressed in a time-dependent manner after transfection with miR-497 mimics. After SMAD7 overexpressed, the proliferation of BC cells recovered (Fig. 3A). To further identify the causes for the proliferation suppression, we performed cell cycle assay by flow cytometry. As shown in Figure 3B, the percentage remaining in S phase was lower in negative control group compared with miR-497 mimics transfected group in MDA-MB-231 cells and MCF-7 cells (28.76% ± 0.11% vs. 17.57% ± 0.14%, 28.14% ± 0.08% vs. 12.49% ± 0.10%, respectively, p < 0.05). The results demonstrated that overexpression of miR-497 in MDA-MB-231 cells and MCF-7 cells induced S phase arrest. In Figure 3C, the invasion assay indicated that the number of cells that passed the Matrigel membrane was significantly decreased in the group transfected with miR-497 mimics in comparison with the negative control group. Also, this phenomenon was weakened after being transfected with SMAD7 expression plasmid. On the basis of the above findings, we concluded that miR-497 was potent at suppressing tumor cell proliferation and invasion by regulating SMAD7 expression.

Enforced expression of miR-497 induces growth inhibition in breast cancer cells in vitro.
miR-497 modulates antitumor efficacy in orthotopic models
Our in vitro study suggested that miR-497 repressed tumor cell growth by targeting SMAD7. To further confirm this, a proof-of-principle experiment was used, using an orthotopic breast cancer model, and a miR-497-mediated antitumor therapeutic approach. On day 10, tumor size of the mimics treatment group reached statistical significance compared with the control group (Fig. 4A). At termination of the study, tumor weight was significantly different between the two groups (Fig. 4B). At 20 days, tumor samples were dissected from mice and prepared for real-time PCR and western blot examination. Similar to results obtained from in vitro, miR-497 expression was induced in tumor specimens from the mimics treatment group (Fig. 4C), whereas SMAD7 expressions were reduced (Fig. 4D). In summary, our in vivo preclinical efficacy trials showed a remarkable antitumor effect of miR-497 by downregulation of SMAD7 in a breast cancer orthotopic mouse model.
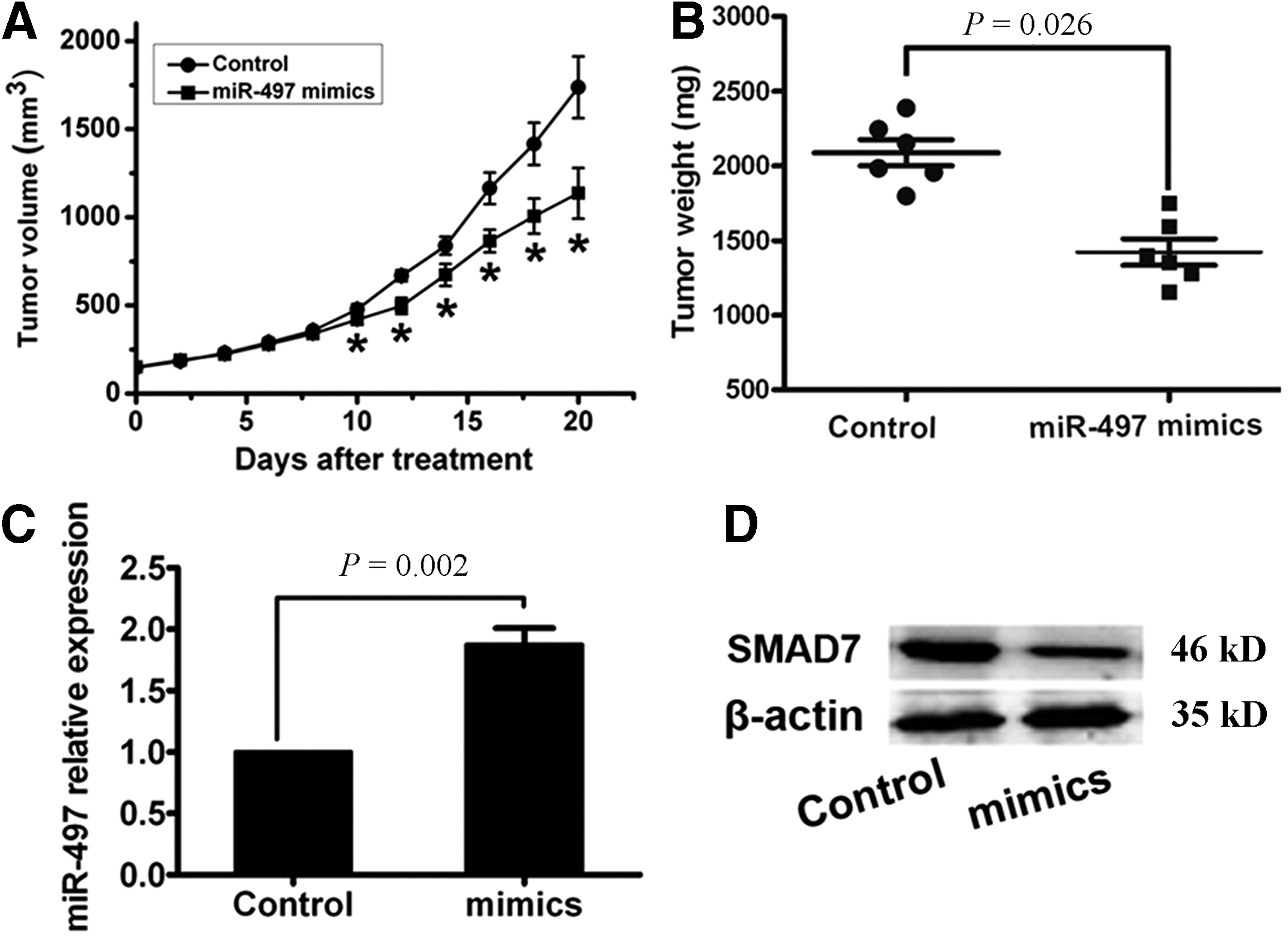
miR-497 suppressed tumor growth in orthotopic breast cancer mouse models.
Correlations of miR-497 expression with the subtypes of breast cancer patients
To investigate the clinical significance of miR-497 expression, the relative expression of miR-497 was determined in 240 cases of breast cancer tissue samples. The clinical characteristics are shown in Table 1. The median value of miR-497 expression was determined as 1.60, therefore, the patients were divided into two groups: high-miR-497 expression group (≥ 1.6; n = 132) and low-miR-497 expression group (< 1.6; n = 108). Then, the correlations of miR-497 expression with the subtype of patients were statistically analyzed. As shown in Table 2, miR-497 expression levels were significantly lower in HER2-positive and triple-negative breast cancer (TNBC) than in luminal subtypes (p = 0.036).
Boldface means statistical significance as P-value < 0.05.
MST, median survival time; HR, hazard ratio; CI, confidence interval.
To further investigate the correlation of miR-497 expression with patients' survival, Kaplan–Meier analyses were performed. As shown in Figure 5A, the 5-year survival of low miR-497 expression group was significantly shorter compared with the high miR-497 expression group (p = 0.0027). These results indicated that miR-497 downregulation might be correlated with the poor survival of breast cancer patients. In the HER2 + subtype, high miR-497 expression also had better 5-year survival compared with the low miR-497 group (p = 0.0142, Fig. 5B). Furthermore, we found that the TNBC subtype with high miR-497 expression had the better 5-year survival compared with the low miR-497 group (p = 0.0391, Fig. 5C). The following Cox regression analysis was performed as Table 3 demonstrates; only grade of BC showed a significant hazard for BC survival (p = 0.008, HR = 4.577, 95% CI = 1.497–13.876). However, low expression of miR-497 also displayed a hazardous trend for BC survival (p = 0.061, HR = 3.299).
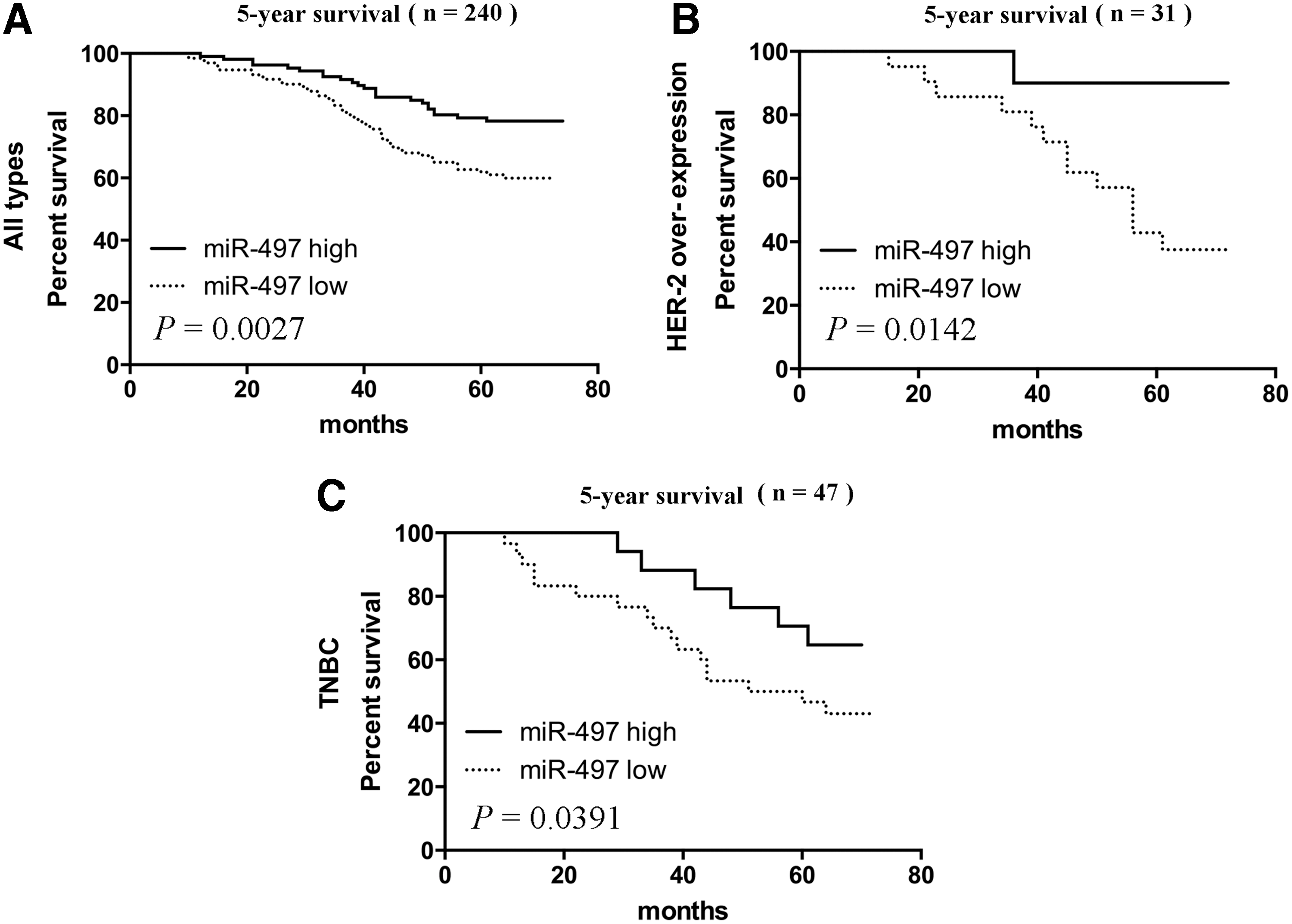
Clinical significance of miR-497 in breast cancer cases.
Boldface means statistical significance as P-value < 0.05.
SE, standard error.
Discussion
Our results display that the low levels of miR-497 are associated with BC progression and therefore could be considered a tumor suppressor. Our data also showed that miR-497 could target the 3′UTR of SMAD7 mRNA, which led to the downregulation of SMAD7 expression. In recovery test, the overexpression of SMAD7 could efficiently reverse the effects of miR-497 in both MDA-MB-231 and MCF-7 cell lines. In survival analysis, the high level of miR-497 was also significantly associated with better survival.
Nowadays, lots of aberrant expressed miRNAs have been reported in breast cancer, which could exert positive or negative function to control the expression of both oncogenes and tumor-suppressor genes to affect carcinogenesis. Leung et al. indicated miR-17, miR-18a, miR-19a/b, and miR-20a were notably elevated, but miR-92a was downregulated in breast cancer cells; they also found that miRNAs could act as biomarkers for radiotherapy effect. In addition, miR-138, miR-106a, and miR-191 were significantly overexpressed in breast cancer compared with normal breast tissue (Leung et al., 2014). Some studies found that miR-497 was downregulated in cervical cancer, colorectal cancer, and breast cancer. A new published data also indicated that miR-497–195 was downregulated in triple-negative breast cancer by deep sequencing (Chang et al., 2015). The expression levels of some miRNAs usually predict the prognostic of tumors. These miRNAs could play a potential role as prognostic biomarkers of cancers (Nana-Sinkam and Croce, 2010). Studies suggested that patients with high-level expression of miR-497 have better prognosis (Guo et al., 2013; Luo et al., 2013a) in cervical and colorectal cancers. However, the clinicopathologic value of miR-497 in human breast cancer was not certain yet.
In this study, we tested the expression level of miR-497 in breast cancer tissues and corresponding adjacent normal tissues. The results showed that miR-497 was significantly downregulated in breast cancer tissues compared with normal tissues, which was consistent with the Dan Li's findings (Li et al., 2011). As an inhibitor of TGF-β signaling, SMAD7 was overexpressed in numerous cancers and its abundance was positively associated with the cancer malignancy (Ekman et al., 2012; Stolfi et al., 2013). Nevertheless, in breast cancer, studies revealed that SMAD7 could exert both anti- and protumor effects. Ectopic SMAD7 sensitized MCF-7 cancer cells to TNF-induced cell death (Hong et al., 2007), abrogated EGF-induced MMP9-9 expression in SKBR3 cells (Kim et al., 2012), and suppressed migration and invasion of mesenchymal-like MCF10CA1h cells (Papageorgis et al., 2010). However, Theohari et al. found that SMAD7 expression was positively correlated with breast cancer tumor size, stage, and MMP9 and MMP14 expressions through the assessment of 150 invasive breast carcinomas by immunohistochemistry (Theohari et al., 2012). The conflicting results of such studies were probably due to its effect under differential conditions, including different cell types and different microenvironments of the cancer tissue. In our study, we found SMAD7 was overexpressed in breast cancer tissues and negatively correlated to the miR-497 expression. And we also confirmed that SMAD7 was a direct target of miR-497 in both MDA-MB-231 and MCF-7 breast cancer cells, which expanded the potential target network of miRNA-497. It would be interesting to investigate whether it is a new regulation mechanism of miR-497 in breast cancer.
As a multifactorial process, survival of BC was enhanced by various intracellular pathways, which usually promote the expression of some imperative proteins involved in cell survival and cell cycle progression. In this study, we analyzed the relationship between miR-497 expression and prognosis of breast cancer patients and found that patients with high miR-497 expression presented better survival. Combined with the relationship between miR-497 and SMAD7 in tissues, we suggested the miR-497 could also influence the BC survival by regulating SMAD7. In our in vivo study, the SMAD7 could cause cell cycle S arrest, which was coinciding to the phenomenon reported by C Stolfi et al. (2014). In the same article, CDC25A was identified as a specific downstream protein of SMAD7, leading to dephosphorylation of CDK2 and inhibition of CDK2 cyclin complex activity (Stolfi et al., 2014). Furthermore, in Cox regression analysis, the miR-497 low expression displayed a hazardous tend for BC survival (p = 0.061), which meant miR-497 could not stand for an independent marker for BC survival in our study (Table 3). However, considering the poor sample size in our study, we suggested that further research could testify the utility of miR-497 in BC survival with a huge sample size. In addition, miR-497 expression levels were significantly lower in HER2-positive and TNBC patients, whereas patients with a higher miR-497 expression had a relatively better 5-year survival rate. These results could also afford a noble therapy strategy for HER2-positive BC and TNBC clinical treatment.
Conclusion
In general, our findings suggested that miR-497 might act as a tumor suppressor in breast cancer, which inhibits breast cancer cell growth and invasion by targeting SMAD7. Thus, this clarification might lead to the development of miR-497-mediated target therapy as a possible treatment of breast cancer and therefore might improve patients' outcome in future.
Footnotes
Acknowledgments
This project was supported by National Science and Technology Support program (2015BAI12B15) and Tianjin municipal Major Scientific and Technological Special Project for Significant Anticancer Development (12ZCDZSY15700). We also sincerely appreciate the Biolight Tec. (Nanjing, China) for their technical help in this article.
Disclosure Statement
No competing financial interests exist.