
Abstract
This study was designed to investigate whether the extract from Boswellia serrata oleo-gum resin (BSE) can protect against glutamate-induced oxidative damage and cytotoxicity in PC12 and N2a cell lines. Using a simple and reliable reverse-phase high-performance liquid chromatography (HPLC), the amount of 3-acetyl-11-keto-β-boswellic acid (AKBA) in the BSE was found to be 18.5% w/w. The results confirmed that BSE and AKBA, at concentrations as high as 100 μg/mL or 10 μM, respectively, caused no significant cytotoxicity or apoptotic cell death. Co- and pretreatment with BSE (25–100 μg/mL) or AKBA (5 μM) restored the viability of PC12 and N2a cells under glutamate toxicity (8 mM). Treatment with BSE and AKBA also attenuated the toxic effects of glutamate on intracellular reactive oxygen species, lipid peroxidation, superoxide dismutase activity, and oxidative DNA damage compared with the untreated glutamate-injured cells. Furthermore, BSE and AKBA decreased the apoptotic cell population in the sub-G1 region and the rate of both early and late-stage apoptosis induced by glutamate in the cells. Our data suggest that the protective effects of Boswellia extract and AKBA against glutamate toxicity in PC12 and N2a cells may be mediated through the amelioration of the oxidative stress and the resultant apoptosis.
Introduction
G
An oleo-gum resin, also known as Olibanum or Frankincense, is naturally obtained from the Boswellia serrata plant belonging to Burseraceae family. Boswellia species are mainly native to India, Northern Africa and Middle East (Archier and Vieillescazes, 2000). Generally, B. serrata oleo-gum resin contains 5–9% essential oil, 20–23% gum, and about 60% resin (Basch et al., 2004).
B. serrata oleo-gum resin has been used in traditional medicine in Asia for treatment of ulcers, dysentery, vomiting, tumors, and chronic inflammatory diseases (Ernst, 2008). Also, the resin has been traditionally used as an herbal product for enhancing memory and learning function (Hosseini et al., 2010). Some recent investigations reported the beneficial properties of B. serrata resin on memory performance (Hosseini et al., 2010; Mahmoudi et al., 2011). A mixture of bioactive components including monoterpenes (α-thujone), diterpenes (such as incensole and its derivatives), triterpenes (such as α- and β-amyrins), pentacyclic triterpenic acids (boswellic acids), and tetracyclic triterpenic acids (tirucall-8, 24-dien-21-oic acids) have been also isolated and identified from the resinous part of B. serrata (Siddiqui, 2011).
Boswellia resin and its isolated compounds have shown various biological activities such as anti-inflammatory (Banno et al., 2006), antitumor (Syrovets et al., 2000; Moussaieff and Mechoulam, 2009), and immunomodulatory effects (Ammon, 2010; Kokkiripati et al., 2011).
Anti-inflammatory activity is mediated through 5-lipoxygenase (5-LOX), human leukocyte elastase inhibition, and the suppression of the nuclear factor kappa-beta (NF-κB) pathway (Anthoni et al., 2006; Poeckel and Werz, 2006; Cuaz-Pérolin et al., 2008, Ammon, 2010). The anticancer activity of boswellic acids (BAs) has been attributed to the triggering of apoptosis via caspase-8 pathway-dependent activation (Liu et al., 2002) and also topoisomerases-I and II-alpha inhibition (Syrovets et al., 2000).
Boswellia resin and incensole acetate were found to attenuate hippocampal neurodegeneration and improve cognitive functions (Moussaieff et al., 2008; Hosseini-Sharifabad and Esfandiari, 2015). In vitro studies reported the enhancement effect of BAs on hippocampal neurites outgrowth and branching (Karima et al., 2010). Additionally, BAs induced neuroprotection against oxidative stress-induced ischemic injury through antioxidant and anti-inflammatory pathways (Ding et al., 2014, 2015).
PC12 (Wang et al., 2010; Kawakami et al., 2011) and N2a (Bezvenyuk et al., 2000) cell lines have been widely used for studying the mechanism of glutamate-induced neurotoxicity. The aim of this study was to investigate whether Boswellia resin extract (BSE) protect the cells against glutamate-mediated oxidative injury and apoptosis.
Materials and Methods
Reagents and chemicals
Annexin V-fluorescein isothiocyanate (FITC) was purchased from Abcam. Fetal bovine serum (FBS), high glucose Dulbecco's modified Eagles medium (DMEM, 4.5 g/L), penicillin, and streptomycin were obtained from Gibco. 3-(4,5-Dimethylthiazol-2-yl)-2,5-diphenyl tetrazolium (MTT), propidium iodide (PI), pyrogalol, 2′,7′-dichlorofluorescin diacetate (DCFH2-DA), bicinchoninic acid (BCA) protein assay detection kit were purchased from Sigma. 3-Acetyl-11-keto-β-boswellic acid (AKBA; Calbiochem),
Preparation of extract of B. serrata oleo-gum resin
The oleo-gum resin of B. serrata obtained from Medicinal Plants Division of Imam Reza Pharmacy (herbarium No. 92083) was washed, dried, and crushed to a powder using an electric microniser. For preparation of ethanolic extract, 10 g of the powder was soaked and gently agitated in 200 mL of ethanol (96%) at 40°C for 48 h. The extract was then filtered and concentrated to remove the solvent using a rotary evaporator at 40°C. The resulting extract was dried and kept at −20°C (Chevrier et al., 2005).
HPLC analytical procedure
Quantification analysis of 3-acetyl-11-keto-β-boswellic acid (AKBA) in ethanolic extract of B. serrata resin was characterized by HPLC-UV method. The chromatographic separation was performed with a reverse-phase waters C18 analytical column (250 × 4.6 mm, 5 μm particle size) and isocratic elution using the mobile phase composed of acetonitrile–water (90:10, % v/v) adjusted to pH 4 with glacial acetic acid at the flow rate of 2 mL/min.
The injection volume of samples was 20 μL and samples were run for a total of 15 min. UV detection wavelength was 420 nm (Shah et al., 2008; Sadeghnia et al., 2014). The sample solutions were filtered through 0.45 μm membrane filter before HPLC analysis.
Cell culture and treatment
PC12 (pheochromocytoma-derived) and N2a (neuroblastoma) cell lines were purchased from Pasteur Institute. The cells were grown in high glucose DMEM (4.5 g/L) supplemented with 10% FBS, 100 IU/mL penicillin, and 100 μg/mL streptomycin, in a humidified atmosphere with carbon dioxide (5% v/v) at 37°C. For measuring the cell viability and intracellular reactive oxygen species (ROS) level, the cells (104 cells/well) were seeded in 96-well culture plate and for other assays, the cells (106 cells/well) were seeded in six well culture plate. First, treatment with different concentrations of glutamate (4–32 mM), BSE (1.5–100 μg/mL), and AKBA (1–10 μM) were conducted.
For evaluating the possible protective effects of Boswellia extract and AKBA against glutamate-neurotoxicity, the cells were pretreated with various concentrations of Boswellia extract (25–100 μg/mL) and AKBA (5 μM) for 2 and 24 h and then exposed to glutamate (8 mM) for 24 h. Also, the cells were co-treated with Boswellia extract at different concentrations (25–100 μg/mL), AKBA (5 μM), and glutamate for 24 h. All tests were performed in triplicate. These concentrations and times were selected based on our preliminary experiments and previous studies (Kokkiripati et al., 2011).
Cell viability assay
PC12 and N2a cells viability was assessed using MTT colorimetric assay. After the treatments, MTT solution (10 μL, at 5 mg/mL) was added to each well of culture plates and incubated for 3 h and then the medium was removed and 100 μL of DMSO was added to solubilize the resulting formazan. MTT reduction by living cells was measured at 570 and 620 nm (background) using an ELISA plate reader (Sadeghnia et al., 2014; Gao et al., 2015). Results were expressed as a percentage of control.
Measurement of intracellular ROS
Intracellular ROS level was detected by H2DCF-DA (2′,7′-dichlorodihydrofluorescein diacetate), which oxidized by ROS and converted to the highly fluorescent compound (DCF).
Before the treatment, H2DCF-DA (10 μL, 200 μM) was added to each well and incubated at 37°C in the dark for 30 min. The cells were then washed with PBS buffer. Afterward, the cultured cells were pre-(2 h), and co-treated with fresh culture medium containing various concentration of Boswellia extract (25–100 μg/mL) and AKBA (5 μM), followed by exposure to glutamate (8 mM). After 2 h incubation, the fluorescence intensity of DCF was measured using a fluorescence plate reader (excitation at 485 nm, emission at 530 nm) (Wu and Yotnda, 2011).
Assessment of lipid peroxidation
Lipid peroxidation (LPO) was measured by thiobarbituric acid reactive substance (TBARS) assay. MDA (malondialdehyde) as a main product of LPO reacts with TBA to generate fluorescence adduct. After the treatments, the cells were scraped into trichloroacetic acid (2.5%, 1 mL) and centrifuged at 13,000 g at 4°C for 2 min. Then, the lysate supernatant (500 μL) was mixed with TBA (0.67%, 800 μL) and TCA (15%, 400 μL). This mixture was heated for 20 min in a water bath (95°C) and cooled in an ice water bath. After centrifugation at 2500 rpm for 10 min at 4°C, fluorescence intensity of each supernatant was determined in excitation and emission wavelengths 530 and 550 nm, respectively. The amount of LPO was estimated by TBARS fluorescence intensity (Ghorbani et al., 2015).
Measurement of superoxide dismutase activity
At the end of the treatment period, the cells were harvested from the plates and washed with ice-cold PBS. The cells were then resuspended and incubated with lysis buffer containing 50 mM Tris-HCl (pH 7.4), 150 mM NaCl, 1% Triton X-100, 1 mM EDTA, and 0.2% sodium dodecyl sulfate (SDS) at 2°C for 30 min. Cell lysates were prepared after centrifugation at 10,000 rpm for 20 min at 4°C and assayed for soluble protein content using BCA kit (Tayarani-Najaran et al., 2013).
Superoxide dismutase (SOD) activity in the cell lysates was evaluated using the procedure previously described based on the ability of the enzyme to inhibit the autoxidation of pyrogallol.
The amount of the SOD inhibiting the rate of pyrogallol auto-oxidation by 50% was defined as one enzyme unit. The percentage of inhibition of pyrogallol autoxidation was determined using the following equation:
where ΔA = absorbance change due to pyrogallol autoxidation in the sample reaction system, ΔAmax = absorbance change due to pyrogallol autoxidation in the control (without cell lysate).
Briefly, 120 μL of cell lysate was added the reaction mixture containing 0.2 mM pyrogallol in Tris-Cl buffer (pH 8.2, 50 mM) and EDTA (1 mM) to initiate the reaction and absorbance decrease of pyrogallol was spectrophotometrically recorded at 420 nm (Garrel et al., 2010).
Comet assay (single-cell gel electrophoresis assay)
The alkaline version of the single-cell gel electrophoresis assay was used to assess DNA damage. The treated cells were suspended in 1% low-melting point (LMP) agarose in PBS (pH 7.4) and spread on a microscope slide (coated with a 1% normal melting agarose [NMP] in PBS layer). Another layer of LMP agarose was also added on top and then, the slides were immediately immersed in cold lysis buffer (2.5 M NaCl, 100 mM Na2EDTA, 10 mM trizma, 1% sarkosyl, 10% dimethyl sulfoxide, and 1% Triton X-100, pH 10) at 4°C in the dark overnight.
The slides were then transferred to a horizontal gel electrophoresis tank containing an alkaline solution (300 mM NaOH, 1 mM Na2EDTA, pH >13) for 40 min to allow DNA unwinding. After that, electrophoresis was conducted at a constant voltage of 23 V and 300 mA at 4°C for 30 min. The slides were then neutralized by tris buffer (0.4 M, pH 7.5). After staining with ethidium bromide (20 μg/mL), the slides were examined using fluorescence microscope (excitation: 520–550 nm, barrier filter: 580 nm). The experiment was done in triplicate and a total of 50 randomly selected cells from each slide were analyzed. The DNA damage was expressed as the percent of DNA in the comet tail (tail DNA%) and calculated using computerized image analysis software (CASP software) (Hosseinzadeh and Sadeghnia, 2007).
Cell cycle analysis
Evaluating the alterations induced in cell cycle progression was performed by flow cytometric analysis of PI-stained nuclei. Cell populations with reduced DNA content was identified by a sub-Gl peak localized below the Go/Gl peak. After treatment, the floating and adherent cells were collected and exposed to the hypotonic buffer (50 μg/mL PI in 0.1% sodium citrate plus 0.1% Triton X-100) and then, incubated at 4°C overnight in the dark. Sub-G1 analysis was performed by BD FACS Canto flow cytometer (BD Biosciences). For each sample 10,000 events were recorded. Analysis of data was performed by Win MDI (version 2.8) software (Sadeghnia et al., 2014).
Flow cytometric analysis of apoptosis by annexin V/PI staining
Early and late apoptotic changes in PC12 and N2a cells were determined using annexin V/PI staining. After the treatments, the cells were collected and washed with PBS, then incubated at room temperature for 5 min in the dark with 500 μL binding buffer containing 1 μL annexin V and 1 μL PI as described by the manufacturer. The flow cytometric analysis was performed by BD FACS Canto flow cytometer (BD Biosciences) and data were analyzed with Win MDI software version 2.9 (Yu et al., 2013).
Statistical analysis
All results were expressed as mean ± SEM from three independent experiments. All statistical analyses were made using Graph Pad Prism version 6. Statistical evaluation of the data was performed for comparison between groups using two-way analysis of variance, followed by Tukey post hoc test. The minimum level of significance was set at p < 0.05.
Results
Quantification of AKBA in BSE
A simple and reliable HPLC method was performed for the standardization of Boswellia resin extract (Fig. 1A, B). Specific retention time identified for AKBA was 6.9 min (Fig. 1A). Standard curve of AKBA were obtained by plotting the peak areas versus corresponding concentrations (Y = 7.7 × −34.6). According to the results obtained from HPLC analysis, the amount of AKBA in the extract estimated to be 18.5%.
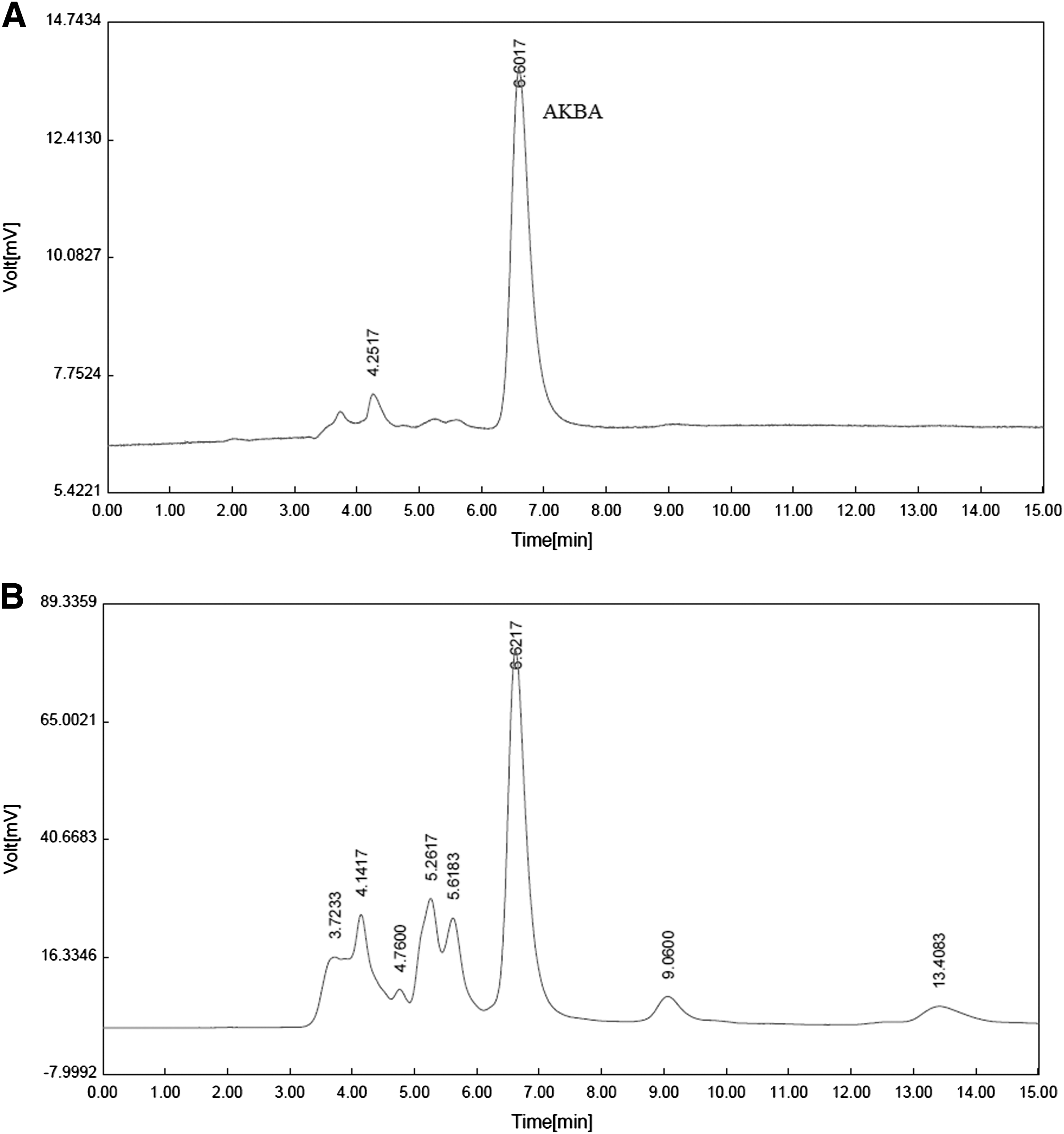
High-performance liquid chromatography chromatograms of 3-acetyl-11-keto-β-boswellic acid (AKBA,
Nontoxicity of BSE and AKBA in PC12 and N2a cells
MTT assay and flow cytometric analysis of the cells in the sub-G1 region were used to identify nontoxicity of BSE and AKBA. The cells were incubated with increasing concentrations of BSE (1.5–100 μg/mL) and AKBA (1–10 μM) for 24 h. As illustrated in Figure 2A and B, BSE and AKBA did not significantly affect the viability of PC12 and N2a cells (p > 0.05). In the same way, no significant differences in the sub-G1 cells population were observed, which reflected that no apoptotic cell death was found after the treatment of PC12 and N2a cells with BSE or AKBA at the used concentrations (Fig. 3A–C).
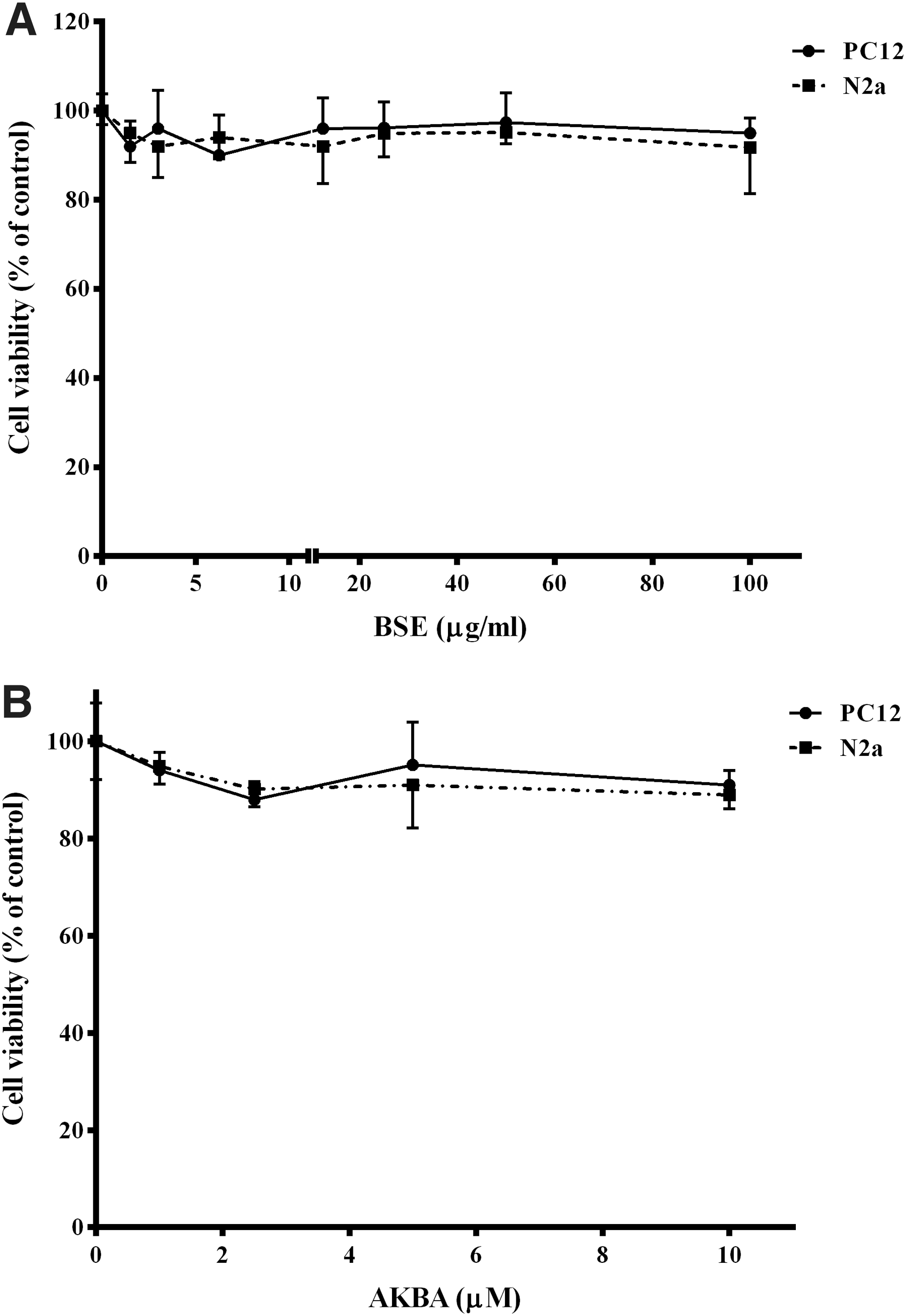
Effect of Boswellia extract (BSE,
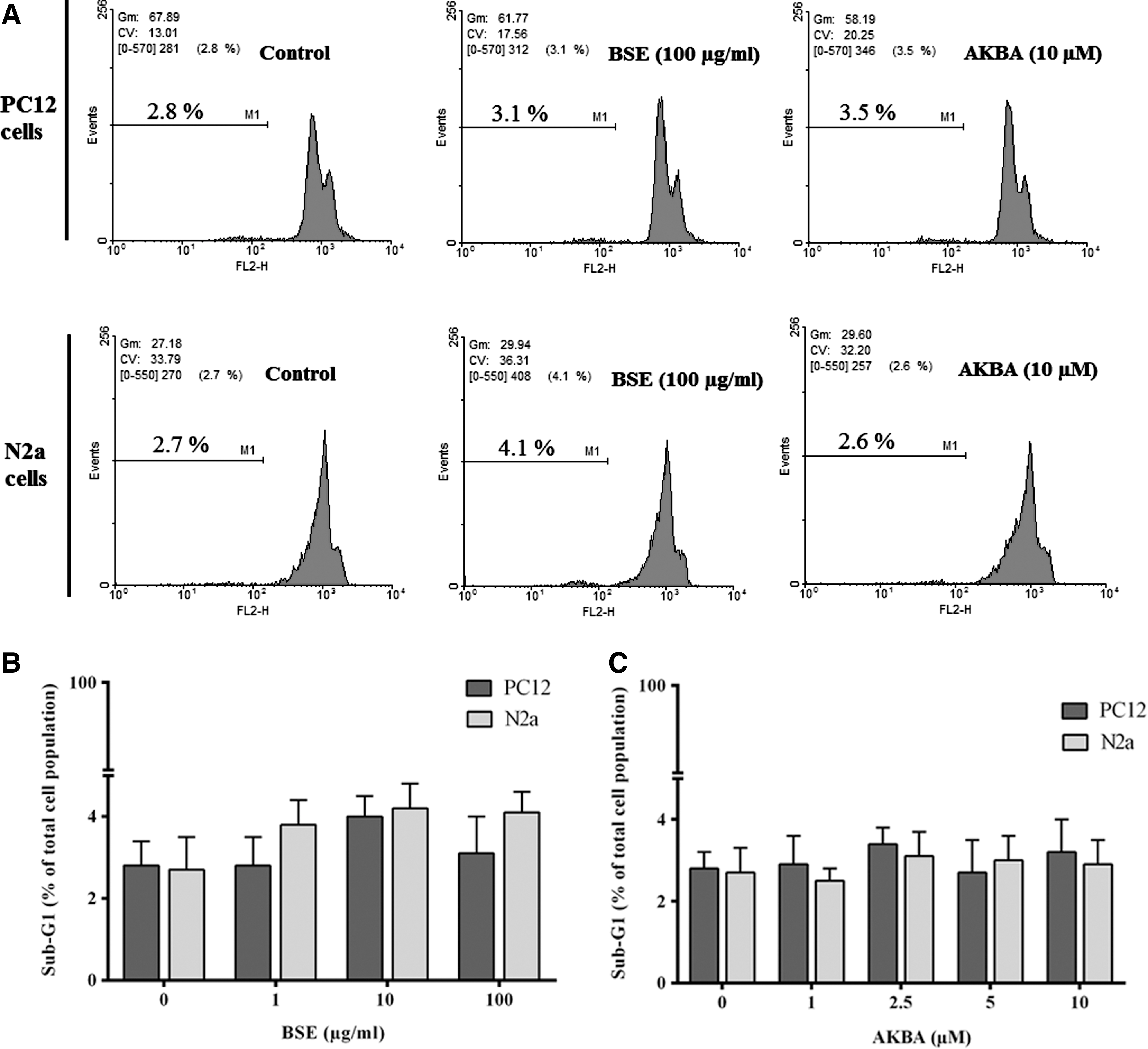
Effect of Boswellia extract (BSE,
Boswellia extract increased cell viability after glutamate-induced cytotoxicity
Assessment of glutamate cytotoxicity through the MTT assay exhibited the dose-dependent toxic effects on viability of the two types of neuronal cell lines in a concentration range of 4–32 mM. Treatment with 8 mM glutamate led to significant decrease in viability of PC12 and N2a cells to 42.5% ± 1.6% and 39.7% ± 1.0% of untreated cells. Therefore, the concentration of 8 mM was chosen for the following experiments (Fig. 4). However, the cytotoxic effects of glutamate were attenuated by co-treatment along with 2 and 24 h pretreatment with BSE at different concentrations (25–100 μg/mL), as revealed by significant increase in viability of PC12 and N2a cells. The viability of PC12 and N2a cells 24 h pretreated with BSE at the highest concentration (100 μg/mL) were increased to 95.0% ± 2.3% and 87.34% ± 6.7% compared to the cells treated with glutamate alone, respectively (p < 0.001). There was no significant difference in the viability of the cells co-treated with 25 μg/mL BSE compared to the cells treated with glutamate alone (Fig. 5).

Effect of glutamate on viability of PC12 and N2a cells. The data were expressed as percentage viability of control cells and presented as mean ± SEM from three independent experiments. **p < 0.01, ***p < 0.001 compared with untreated PC12 cells. c p < 0.001 compared with untreated N2a cells.
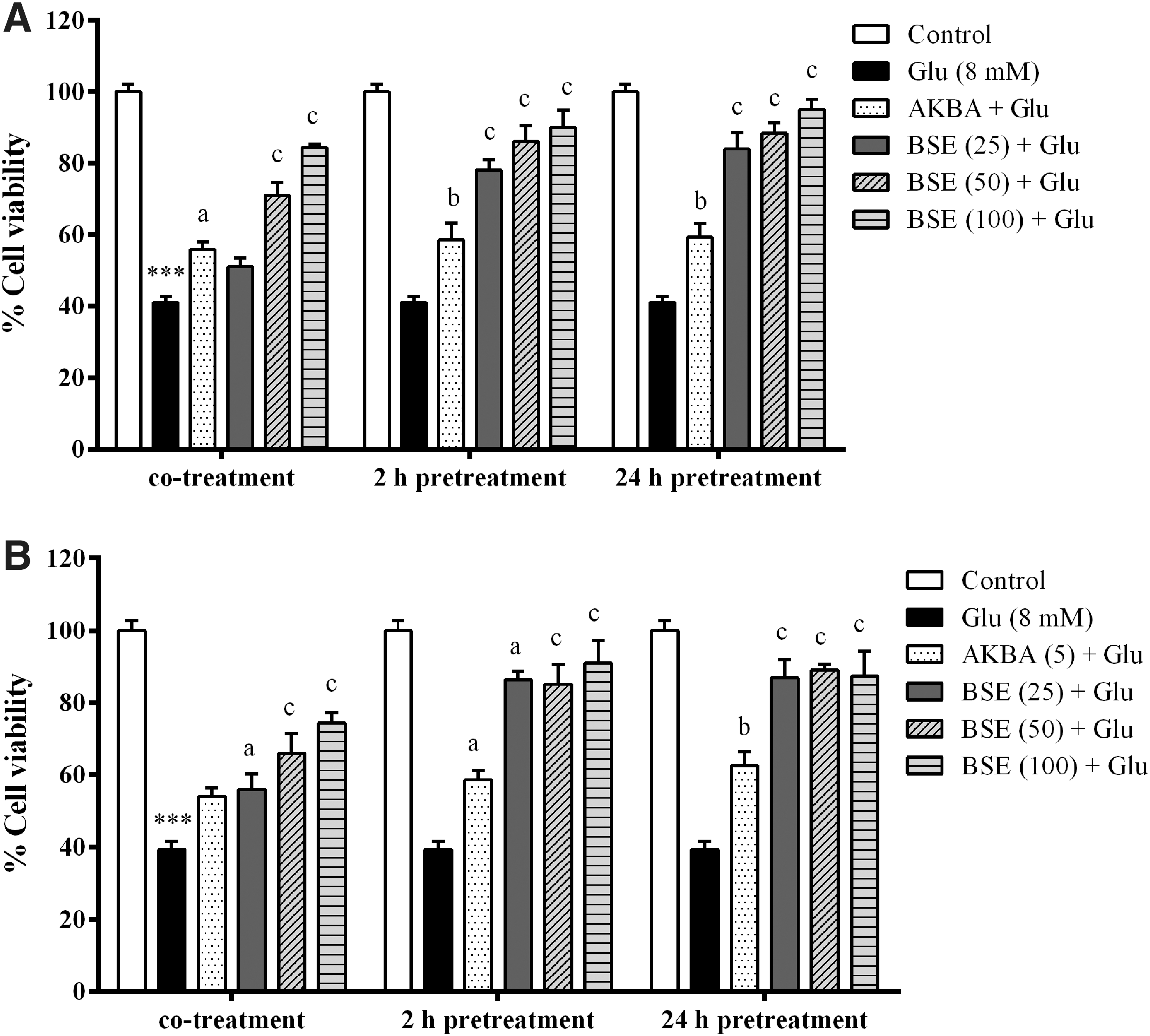
Effect of Boswellia extract (BSE) and 3-acetyl-11-keto-β-boswellic acid (AKBA) on glutamate (Glu)-induced toxicity in PC12
Since no significant difference was observed between 2 and 24 h pretreatment groups, the cells were only pretreated with the extract 2 h before glutamate exposure for subsequent experiments.
Boswellia extract attenuated glutamate-induced generation of ROS in PC12 and N2a cells
ROS generation was significantly elevated in PC12 and N2a cells after exposure to 8 mM glutamate for 2 h, compared to the control group (p < 0.001). However, co-treatment and pretreatment with the extract at different concentrations (25–100 μg/mL) caused a significant decrease in the generation of ROS compared to the cells treated with glutamate alone (p < 0.001). Intracellular ROS content was decreased in PC12 and N2a cells pretreated with BSE at the highest concentration (100 μg/mL) to 138.7% ± 3.5% and 151.3% ± 4.4% as compared to glutamate-treated cells (232.4% ± 7.5% and 234.6% ± 10.4% of control), respectively (Fig. 6).
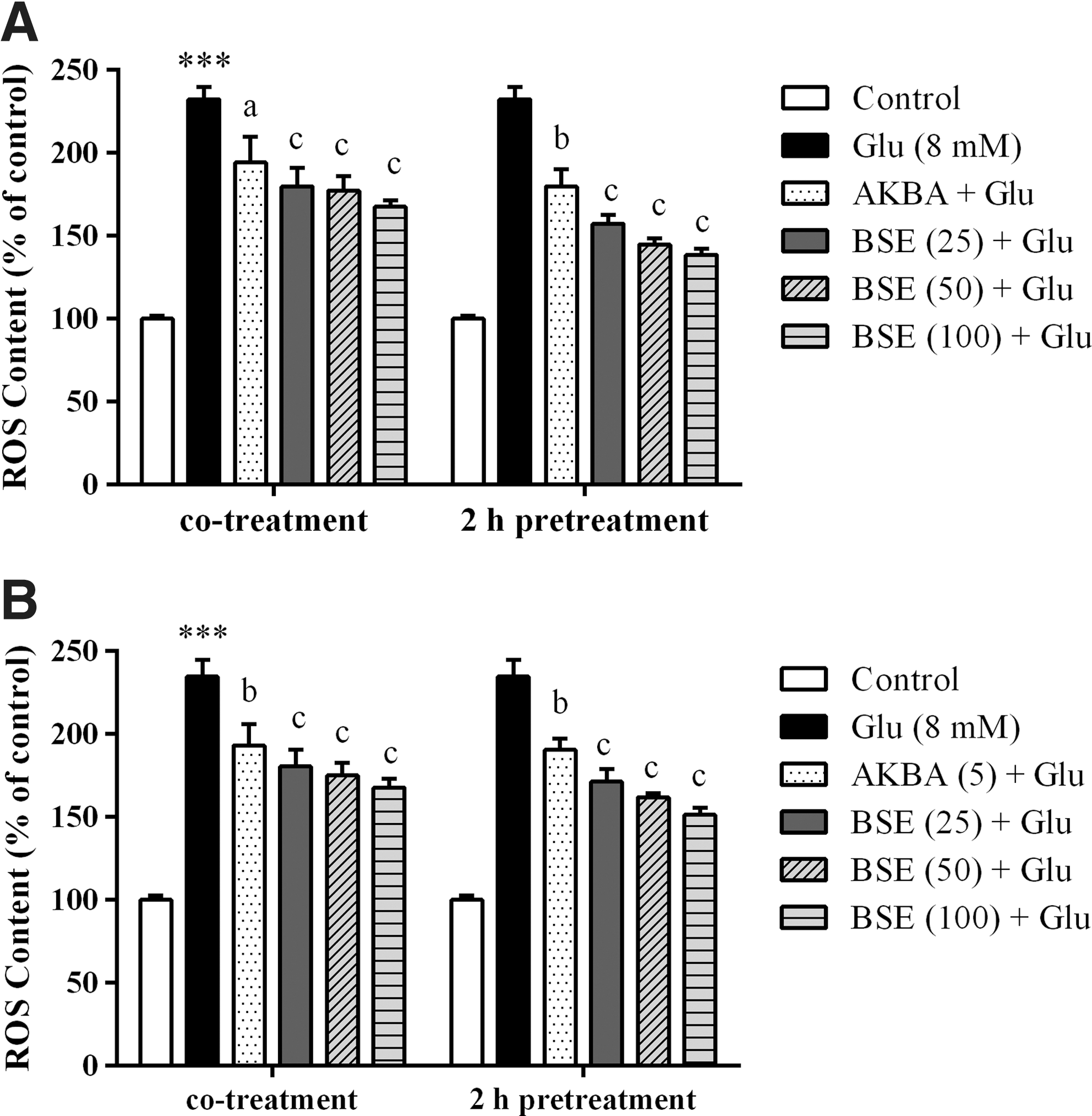
Effect of Boswellia extract (BSE) and 3-acetyl-11-keto-β-boswellic acid (AKBA) on ROS generation in PC12
Boswellia extract reduced the LPO induced by glutamate in PC12 and N2a cells
To investigate whether Boswellia decrease LPO induced by glutamate, TBARS method was performed. As shown in Figure 7, PC12 and N2a cells treated with 8 mM glutamate showed a significant increase in the LPO levels when compared with the untreated cells (p < 0.001). In both cell lines, pretreatment with Boswellia extract (25–100) for 2 h before glutamate exposure reduced LPO contents. Similarly, co-treatment with the extract showed significant decrease in LPO contents. However, co-treatment of PC12 cells with 25 μg/mL BSE did not significantly reduce the LPO content increased by glutamate. LPO level was decreased in PC12 and N2a cells pretreated with BSE at the highest concentration (100 μg/mL) to 124.3% ± 6.5% and 132.8% ± 7.4%, as compared to the cells treated with glutamate alone (183.9% ± 1.4% and 188.1% ± 1.00% of control), respectively (Fig. 7).
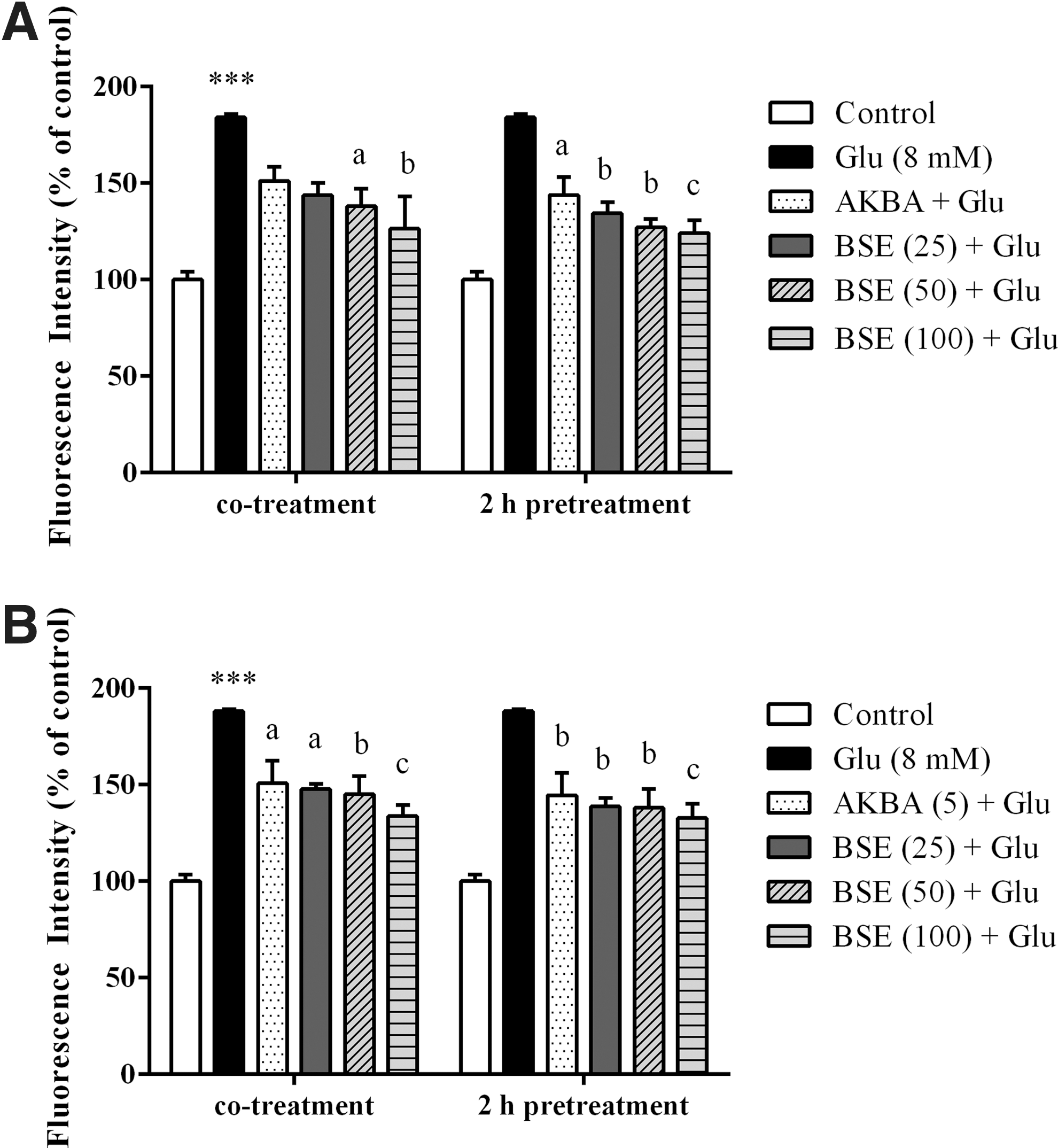
Effect of Boswellia extract (BSE) and 3-acetyl-11-keto-β-boswellic acid (AKBA) on glutamate (Glu)-induced lipid peroxidation in PC12
Boswellia extract increased SOD activity in glutamate-exposed PC12 and N2a cells
As shown in Figure 8, PC12 and N2a cells exposed to 8 mM glutamate for 24 h showed a significant decrease in SOD activity compared to untreated cells (p < 0.001). The SOD activity was increased significantly when the cells were co- and pretreated with Boswellia extract compared to glutamate-treated cells. However, co-treatment of PC12 cells with 25 μg/mL BSE resulted in a nonsignificant decrease in SOD activity. SOD activity was increased in PC12 and N2a cells pretreated with BSE at the highest concentration (100 μg/mL) to 1.1 ± 0.02 and 1.0 ± 0.05 compared to the cells treated with glutamate alone (0.8 ± 0.06 and 0.7 ± 0.03 [unit/min/mg protein], respectively).
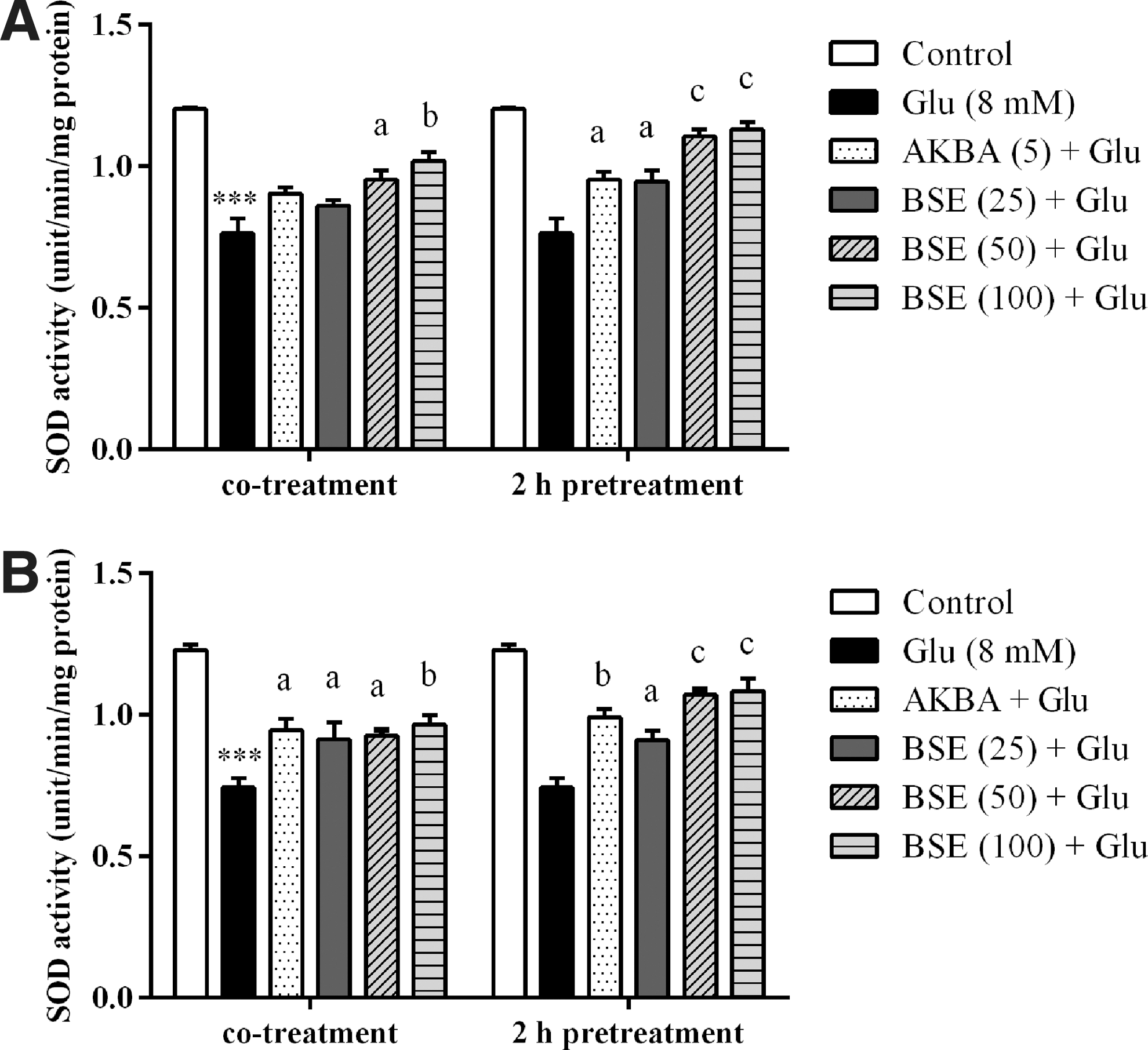
Effect of Boswellia extract (BSE) and 3-acetyl-11-keto-β-boswellic acid (AKBA) on SOD activity in PC12
Boswellia extract attenuated glutamate-induced oxidative DNA damage in PC12 and N2a cells
DNA tail percentage was measured by comet assay as an indicator of DNA damage and fragmentation. The results showed that exposure of PC12 and N2a cells to 8 mM glutamate increased the percentage of DNA in the comet tails compared with control groups (p < 0.001). The DNA fragmentation in cells treated with the extract (25–100 μg/mL) in the presence of 8 mM glutamate for 24 h was significantly decreased when compared to cells treated with glutamate alone. The percentage of DNA tail was decreased in PC12 and N2a cells when exposed with BSE at the highest concentration to 4.6% ± 1.3% and 9.4% ± 3.2% compared to cells treated with glutamate alone (34.0% ± 7.0% and 66.0% ± 13.2%), respectively (Fig. 9).
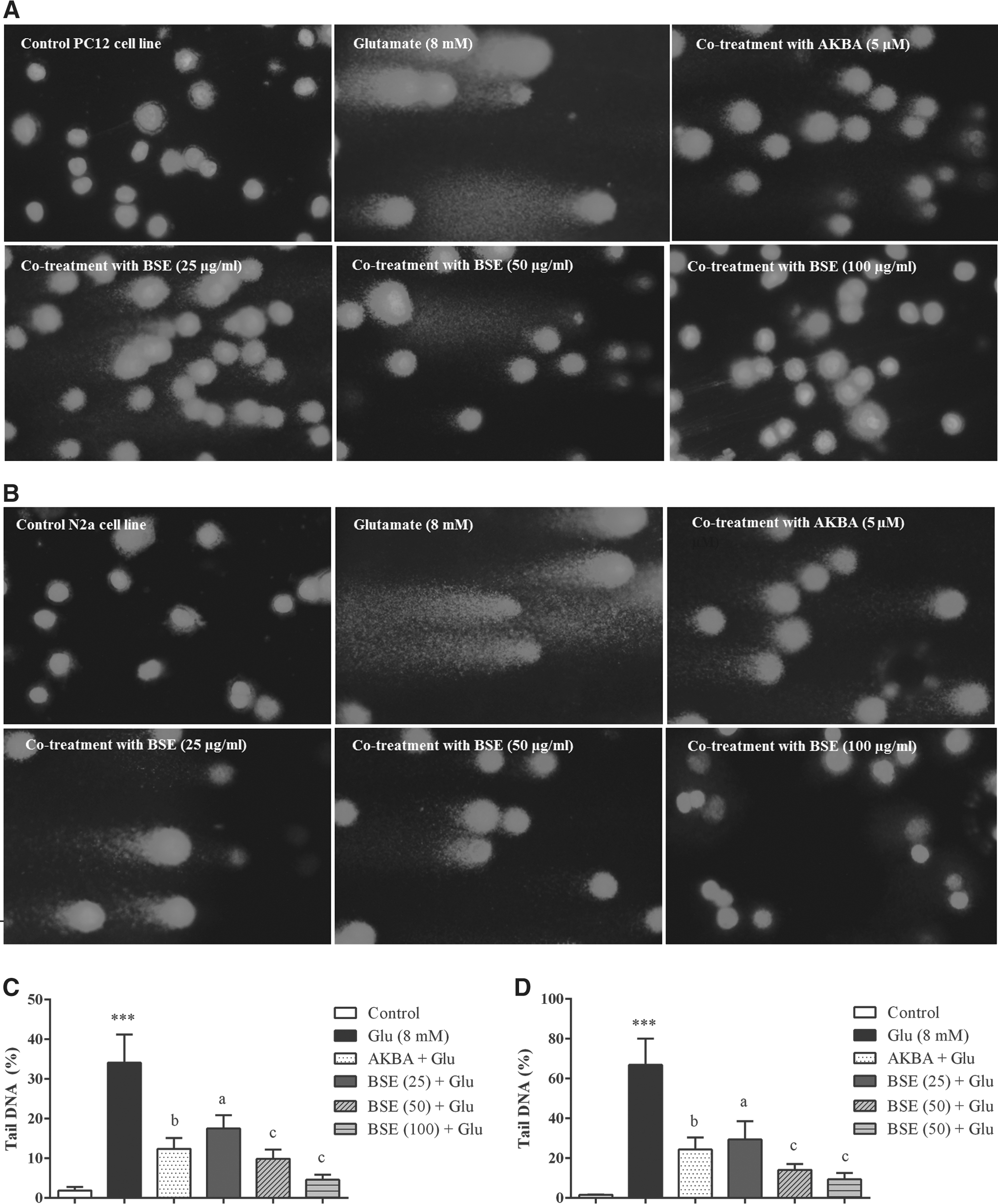
Effect of Boswellia extract (BSE) and 3-acetyl-11-keto-β-boswellic acid (AKBA) on glutamate (Glu)-induced DNA damage in PC12 and N2a cells. The cells were treated with BSE (25–100 μg/mL) and AKBA (5 μM) in the presence of 8 mM glutamate for 24 h.
Boswellia extract reduced sub-G1 cell population in glutamate-treated PC12 and N2a cells
For investigation of possible apoptogenic properties of BSE and AKBA, the cells were treated with different concentrations of BSE (25–100 μg/mL) and AKBA (5 μM) for 24 h.
Glutamate exposure significantly increased the number of cells with reduced DNA content, accumulated in the sub-G1 region, when compared with the control group (p < 0.001). However, no significant changes in cell cycle distribution were observed. Co- and pretreatment and with Boswellia extract significantly decreased sub-G1 cell population compared to cells treated with glutamate alone. The percentage of sub-G1 cell population was decreased in PC12 and N2a cells pretreated with BSE at the highest concentration (100 μg/mL) to 3.4% ± 0.9% and 15.7% ± 1.2%, compared to cells treated with glutamate alone (40.0% ± 1.8% and 54.2% ± 2.7%), respectively (Fig. 10).
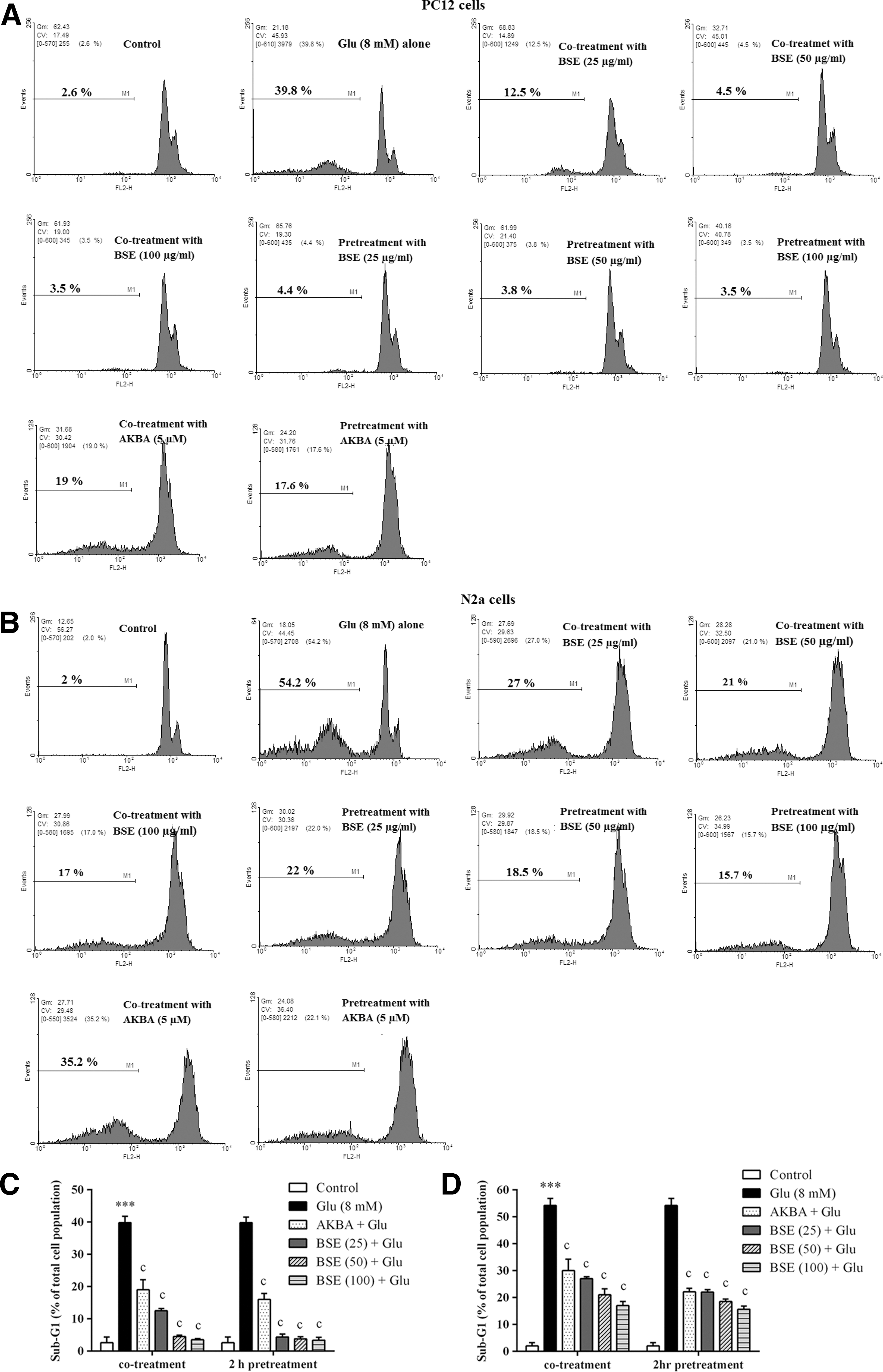
Effect of Boswellia extract (BSE) and 3-acetyl-11-keto-β-boswellic acid (AKBA) on sub-G1 cell populations in glutamate (Glu)-treated PC12 and N2a cells. The cells were co- and 2 h pretreated with various concentrations of the extract (25–100 μg/mL) and AKBA (5 μM), then exposed to glutamate (8 mM) for 24 h, in which the same treatments were applied.
Boswellia extract protected against glutamate-induced apoptosis in PC12 and N2a cells
To evaluate the apoptotic cell death, annexin V-FITC/PI double staining method was conducted. No significant difference was observed in the percentage of early and late apoptotic cells treated with BSE (100 μg/mL) or AKBA (5 μM) compared to untreated cells (p > 0.05). The annexin V+/PI− (lower right quadrant) were considered to represent early apoptotic cells, the annexin V+/PI+ (upper right quadrant), and annexin V−/PI− (lower left quadrant) represented late apoptotic and nonapoptotic cells, respectively. Treatment with glutamate caused a significant increase in the number of apoptotic cells (p < 0.001).
After treatment with Boswellia extract, the number of viable cells was significantly increased, whereas the numbers of both early and late apoptotic cells were significantly decreased (p < 0.001). BSE at 100 μg/mL significantly prevented apoptosis, decreasing the percentage of both early and late apoptotic cell death to 12.2% ± 3.2% and 12.0% ± 2.6% when compared to cells treated with glutamate alone (40.1% ± 2.7% and 44.0% ± 3.2%), respectively (Fig. 11).

Effect of Boswellia extract (BSE) and 3-acetyl-11-keto-β-boswellic acid (AKBA) on glutamate (Glu)-induced apoptosis in PC12 and N2a cells. The cells were treated with BSE (25–100 μg/mL) and AKBA (5 μM) in the presence of 8 mM glutamate for 24 h.
Discussion
Neuroprotective properties of Boswellia oleo-gum resin and its isolated compounds have been reported in previous studies. However, there have been no reports regarding the protective effects of Boswellia against oxidative cell injury and apoptotic cell death in glutamate toxicity model. Among the four major boswellic acids from Boswellia resin, AKBA was identified as the most potent anti-inflammatory and anticancer compound (Liu et al., 2002; Poeckel and Werz, 2006). In this study standardization and identification of the constituent in BSE was performed. Also, for the first time, protective effects of BSE against glutamate cytotoxicity was evaluated.
HPLC analysis of BSE confirmed the presence of AKBA (18.5% w/w) in the extract. The amount found in the extract was greater than those reported in the previous researches (Umar et al., 2014). The data of this study confirmed that BSE and AKBA, at concentrations as high as 100 μg/mL or 10 μM, respectively, caused no significant cytotoxicity or apoptotic cell death. The study also showed that treatment with glutamate decreased the viability of PC12 and N2a cells, in a concentration-dependent manner, with significant cytotoxicity being observed at concentrations >4 mM.
The results have shown that both 2 and 24 h pretreatment regimens of BSE and its constituent, AKBA, were significantly restored cell death induced by glutamate; therefore, the 2 h pretreatment regimen was used for subsequent experiments. Furthermore, BSE and AKBA significantly attenuated the rate of both early-and late-stage apoptosis induced by glutamate in PC12 and N2a cells. The antiapoptotic effect was further validated by the significant decrease of the apoptotic cell population in the sub-G1 region.
In this study, excessive amounts of glutamate appeared to mediate oxidative toxicity in PC12 and N2a cells. Glutamate was found to disrupt the glutathione-cystine antiporter system and cause depletion of glutathione and ROS accumulation in the cell (Penugonda et al., 2005; Farombi and Onyema, 2006). Glutamate-induced ROS generation could trigger proapoptotic cell signaling leading to neural cell death (Li et al., 2016). Inhibition of ROS is a promising strategy for blocking glutamate-triggered apoptosis in the cells (Ma et al., 2012).
Interestingly, our data revealed that BSE and AKBA significantly impaired glutamate-induced excessive ROS formation. Consistent with suppression of ROS production, BSE and AKBA treatment resulted in a significant reduction in LPO, oxidative DNA damage, and restoring the SOD activity. LPO, caused by free radicals such as ROS, has been involved in glutamate-induced cytotoxicity (Ma et al., 2012). Glutamate-induced ROS generation particularly triggers DNA damage by attacking deoxyribose that results in single and double strand breaks (Yang et al., 2010).
Since neuronal cells possess low levels of endogenous antioxidant enzymes, the cells are particularly susceptible to ROS-induced neuronal cell death or apoptosis. Oxidative stress can contribute to apoptosis involved in the pathogenesis of neurodegenerative diseases (Dong et al., 2009). These findings suggest that antiapoptotic activity of BSE and AKBA was associated with suppression of oxidative injury in cells. Boswellia resin extract was found to reduce LPO and induce cardioprotective effects in a model of myocardial infarction induced by isoprenaline (Zaki et al., 2014).
Bhushan et al. (2007) reported that a natural triterpenediol from Boswellia induced apoptosis through ROS and NO generation in human promyelocytic leukemia cell line, HL-60. Also, BAs considerably stimulated ROS formation in HL-60 (Altmann et al., 2004). Therefore, BSE and its components may act as both pro-oxidant and antioxidant, depending on the redox state of the biological environment.
Catanzaro et al. (2015) demonstrated that BSE and AKBA preserved intestinal epithelial barrier from oxidative and inflammatory damages at low concentrations (1 and 0.027 μg/mL, respectively) in an experimental model of intestinal inflammation, in vitro. The concentrations (25–100 μg/mL) used in this study is greater than those reported by Catanzaro et al. (2015). One possible explanation is the role of 5-lipoxygenase (5-LOX) enzyme in the intestinal inflammatory conditions. 5-LOX expression has been reported to be significantly upregulated in human colorectal cancer (Soumaoro et al., 2006). In addition, the efficacy of 5-LOX inhibitors in the inhibition of cancer cell growth has been well documented (Melstrom et al., 2008). It has been shown that Boswellia inhibits 5-LOX in concentrations greater than 10–15 μg/mL, in vitro. AKBA is also the most potent (among BAs) inhibitor of 5-LOX with IC50 value in the range of 1.5 to 50 μM, depending on the cell type or assay conditions (Safayhi et al. 1997; Werz et al. 1997).
Moreover, consistent with our results, Kokkiripati et al. (2011) also showed that B. serrata at relatively high concentrations (300–400 and 30–40 μg/mL for aqueous and ethanoic extracts, respectively) markedly attenuated H2O2-mediated ROS generation and toxicity in human monocytic (THP-1) cells.
B. serrata extract and its active constituents were found to act as anti-inflammatory through inhibiting 5-lipoxygenase (Pockel and Werz, 2006), antioxidant (Farshchi et al., 2010), and free radical scavenger agent (Krüger et al., 2008). It was found that anti-inflammatory properties of incensole acetate contributed to the protective effects of B. serrata against hippocampal neurodegeneration (Moussaieff et al., 2008).
Karima et al. (2010) reported the enhancement effect of β-boswellic acid on neurite outgrowth and branching and on polymerization dynamics of tubulin, in vitro. B. serrata resin and BAs also inhibited the oxygen radicals and proinflammatory enzymes released by leukocytes and macrophages in inflammatory conditions (Ammon, 2010). In previous researches, aqueous and alcohol extracts of Boswellia exhibited high free radicals (diphenyl picryl hydrazyl, nitric oxide, hydroxide, and super oxide) scavenging activity, and reducing ability, in a concentration-dependent manner (Sharma et al., 2011; Azemi et al., 2012). Also, antioxidant activity of BAs has been demonstrated in many in vivo conditions such as pulmonary fibrosis and oxidative stress induced by bleomycin (Ali and Mansour, 2011).
Conclusion
BSE and AKBA significantly protected the cells in both co- and 2 h pretreatment regimens against glutamate toxicity. These results suggest that direct scavenger activity and enhancement of antioxidant capacity of cells could be involved in the protective effect of BSE and AKBA against damage induced by ROS. These pharmacological properties of BSE are closely related to the presence of active ingredients, probably by modulating oxidative stress and apoptosis. These results suggest that BSE is a potential neuroprotective agent that acts by inhibiting apoptosis. However, future investigations are needed to precisely understand the cellular mechanisms involved in the neuroprotective effects of Boswellia for the treatment of neurodegenerative disorders.
Footnotes
Acknowledgment
The project was financially supported by Vice Chancellor for Research, Mashhad University of Medical Sciences, Mashhad, Iran.
Disclosure Statement
No competing financial interests exist.