
Abstract
The critical roles of IGFBP3 in regulating the osteogenic and adipogenic differentiation of bone marrow mesenchymal stem cells strongly indicate its potential effects on the pathogenesis of osteonecrosis of the femoral head (ONFH). In this study, we investigated the association of IGFBP3 gene polymorphism and its protein expression with the development of ONFH to further explore its molecular pathogenesis. Ligase detection reactions and enzyme-linked immunosorbent assay were used to detect the polymorphisms of rs2453839[C/T] and rs3110697[A/G] and the serum protein expression of IGFBP3 gene in 182 cases and 179 controls, respectively. The serum lipids level was also measured by automatic biochemistry analyzer. The results revealed that the recessive model of rs3110697 and the dominant model of rs2453839 were significantly associated with the increased risk of ONFH (p = 0.048, p = 0.047, respectively). The genotypes of rs2453839 were also significantly related to the clinical stages of ONFH (p = 0.017). More importantly, the serum protein expression of IGFBP3 and insulin-like growth factor 1 (IGF1) in the ONFH group were statistically increased compared with the control group (p = 0.044, p = 0.007). The serum triglyceride and low-density lipoprotein cholesterol level in the ONFH group were significantly higher than the control group (p = 0.01, p = 0.005, respectively), but the serum high-density lipoprotein cholesterol level of the ONFH group was dramatically lower than the control group (p = 0.0001). Our results showed that both the gene polymorphisms of IGFBP3 and the abnormal protein expression of serum IGFBP3 and IGF1 closely associated with the development of ONFH.
Introduction
T
The IGFBP3 gene is located on chromosome 7p12.3 and is a member of the insulin-like growth factor binding proteins (IGFBPs) family. IGFBP3 is a well-documented cell growth inhibitor and/or apoptosis promoter that competitively binds to insulin-like growth factor 1 (IGF1) to inhibit its activity (Lee et al., 2014). IGFBP3 forms a ternary complex with insulin-like growth factor acid-labile subunit and either IGF1 or IGF2. In this form, it circulates in plasma, prolonging the half-life of IGFs and altering their interaction with cell surface receptors (Dar et al., 2010; Langkamp et al., 2010; Moya et al., 2015). IGFBP3 gene polymorphism has more been reported to correlate with tumor risk, but has rarely been involved in ONFH risk. In 2010, Hong first demonstrated that the rs2453839 mutation of the IGFBP3 gene and high serum IGFBP3 levels were closely correlated to ONFH risk (Hong et al., 2010). Subsequently, in 2012, our sequencing results also confirmed that IGFBP3 rs2453839 genotypes were significantly related to the development of ONFH (Song et al., 2012). Furthermore, it has been demonstrated that IGFBP3 plays key roles in regulating bone marrow mesenchymal stem cells into osteogenic and adipogenic differentiation in recent years, indicating the potential effects of IGFBP3 on the fat accumulation in the damaged bone marrow cavity of ONFH. Thus, we investigated the genotypes, haplotypes of two tag single-nucleotide polymorphisms (TagSNPs), rs2453839[C/T], and rs3110697[A/G] of IGFBP3 gene, serum protein expression of IGFBP3 gene, and their association with the lipid metabolism disorder and the clinical phenotypes of ONFH in 182 cases and 179 controls to explore the role of IGFBP3 gene polymorphism and its expression in the development ONFH.
Materials and Methods
Patients
A total of 182 unrelated patients with ONFH (126 males, 56 females; age: 53.50 ± 12.63 years) were consecutively enrolled at the Department of Orthopedics, the Second Clinical College of Jilin University (Changchun, China), from March 2014 to June 2015. Patients with ONFH that was caused by direct trauma and patients with ONFH concurrent with cardiovascular diseases (arterial thrombosis, atherosclerosis, and so on), congenital diseases, human immunodeficiency virus infection, diabetes mellitus, or renal dysfunction were excluded. ONFH diagnoses were established by evidence of osteonecrosis using plain radiographs in stages II, III, and IV of the Ficat Classification system (Ficat, 1985). According to the inquiry of detailed disease history and etiological factors, ONFH cases were classified into one of the following subgroups: alcohol-induced [68 cases (38.42%)], idiopathic [67 cases (37.85%)], and steroid-induced osteonecrosis [45 cases (25.42%)]. Steroid-induced osteonecrosis was defined by a history of taking prednisolone cumulative 2000 mg or an equivalent over 21 days. Alcohol-induced osteonecrosis was defined by the consumption of more than 900 mL of pure ethanol per week.
The disease course of ONFH ranged from 0.5 to 240 months, with an average of 72.45 months, and the clinical stages of ONFH consisted of 13 cases of stage II (7.34%), 52 cases of stage III (29.38%), and 112 cases (63.28%) of stage IV. There were six cases of ONFH patients who failed to undergo the clinical stages or etiological classification due to the defect of plain radiographs or unclear etiological factors. Moreover, 179 unrelated control subjects (114 males, 65 females; age: 52.58 ± 11.08 years) who were age- and sex-matched for the ONFH group were consecutively enrolled at the Health Examination Center of Second Clinical College of Jilin University (Changchun, China) from October 2014 to December 2014. A comparison of age and gender between ONFH and Control groups is seen in Table 1. Control subjects were defined in the following manner: they had no hip pain, their fasting blood glucose, triglyceride (TG), and total cholesterol (TC) levels in serum were within the normal reference value range, their abdominal ultrasound examination and chest X-ray radiography were normal, and they did not have a presence of cardiovascular or cerebrovascular diseases. All participants were Han Chinese from northeast China. The study was approved by the ethics committee of the Second Clinical College of Jilin University (Changchun, China) and conformed to the current ethical principles of the Declaration of Helsinki. All individuals provided informed consent for their participation in the study.
ONFH, osteonecrosis of the femoral head; SD, standard deviation.
Genomic DNA extraction and SNP selection
Genomic DNA was extracted from 2-mL blood using the genomic DNA extraction kit (DP318; TianGen, Beijing, China) following the manufacturer's protocols. The HapMap database and related literature were used to select TagSNPs of the IGFBP3 gene by analyzing their population distribution in different countries, nationalities, and regions, particularly in data obtained from an Asian population. The two tag SNPs in the IGFBP3 gene, rs3110697 and rs2453839, were selected based on linkage disequilibrium (LD) analysis by HapMap (
Genotyping
The detection primer, probe sequence, and product size after the ligase reaction of rs2453839 and rs3110697 are shown in Table 2. PCR was performed in a buffer containing 1-μL DNA, 1.5-μL MgCl2, 0.3-μL DNTP, 0.15-μL primer mix, and 0.3-μL Taq DNA ligase in a final reaction volume of 15-μL (all reagents from the ABI Company, Willimantic, CT). The reaction mixture was heated to 94°C for 3 min for denaturation and then subjected to 35 cycles at 94°C for 15 s, annealing at 54°C for 15 s and extension at 72°C for 30 s, and a final extension at 72°C for 5 min. The specific amplified fragments were used in an ligase detection reaction (LDR) assay to identify the mutations related to rs2453839[C/T] and rs3110697[A/G]. The LDR assay was performed in a reaction volume of 10 μL that contained 3 μL PCR product, 1 μL 10 × ligase reaction buffer, 0.125 μL (40 U/μL) Taq DNA ligase, and 0.01 μL probe (10 pmol)/each probe, added to deionized H2O to 10 μL. The ligation reaction was performed using a GeneAmp PCR System 9700 (ABI Company) with the following temperature program: 2 min at 95°C and 30 cycles of 30 s at 94°C and 3 min at 56°C. The products were analyzed using an ABI PRISM 3730xl DNA sequencer (ABI Company). CHROMAS software was used to analyze the sequencing peak chart.
Primer to amplify the gene fragment of the target dbSNP site.
Probe to discriminate the target SNP genotype.
Product size indicates the size of the ligase reaction product from the different SNP genotypes.
SNP, single-nucleotide polymorphism.
IGFBP3 and IGF1 protein expression and lipid detection in serum
The expression of IGFBP3 and IGF1 protein in serum was detected using enzyme-linked immunosorbent assay (ELISA) kits (DRE10118; Solarbio Company, Shanghai, China) according to the manufacturer's protocols. Briefly, the assay range of IGFBP3 and IGF1 was 5–100 μg/L and 4–200 μg/L, respectively. Set standard wells on the ELISA plates coated (density: 90, 60, 30, 15, 7.5 μg/L for IGFBP3 and IGF1, respectively). Set blank wells separately (blank wells do not add sample and horseradish peroxidase [HRP]-conjugate reagent, other step is the same). Add sample dilution 40 μL to sample wells and then add testing sample 10 μL (sample final dilution is fivefold). Incubate for 30 min at 37°C, then wash with washing buffer, and repeat five times; add HRP-conjugate reagent 50 μL, incubate for 30 min at 37°C, and wash five times; add Chromogen Solution A and B 50 μL separately, keep in dark place for 15 min at 37°C, and add Stop Solution 50 μL to each well. After adding Stop Solution within 15 min, take the blank well as 0, and read absorbance at 450 nm using microplate reader of multiwavelength (Varioskan, Flash; Thermo Scientific, Waltham, MA). The serum TC, TG, high-density lipoprotein cholesterol (HDL-c), and low-density lipoprotein cholesterol (LDL-c) level were measured by automatic biochemistry analyzer (AU680; Beckman Coulter, Inc., Brea, CA).
Statistical analyses
SHEsis software (
Results
Genotypes, allele frequencies, and haplotypes of rs3110697 and rs2453839 of the IGFBP3 gene
We calculated the LD coefficients (|D′| and r
2) between rs3110697 and rs2453839 using SHEsis software (
11, homozygotes for the major allele; 12, heterozygotes; and 22, homozygotes for the minor allele. Logistic regression analyses were used for calculations. Bold values: p < 0.05.
p-Values of deviation from Hardy–Weinberg equilibrium between the ONFH group and control group.
CI, confidence intervals; MAF, minor allele frequency; OR, odds ratios.
Association of genotypes and haplotypes of the IGFBP3 gene with the clinical phenotypes of ONFH
We completed the correlation analysis between the genotypes of 3110697, rs2453839 and gender, disease duration, etiological classification, and clinical stages. The results revealed that the genotypes of rs2453839 were significantly associated with the clinical stages of ONFH (among stage II, III, and IV, p = 0.017). Among them, the stage IV ratio of the CT or CC genotype carriers was statistically increased or decreased compared with that of stage III (p = 0.027) or stage II (p = 0.012), respectively. In addition, in the TT genotype carriers, the stage IV ratio also was statistically lower compared with stage II (p = 0.027), as shown in Table 4. The haplotypes of the IGFBP3 gene failed to show statistical association with hip lesions and clinical stages of ONFH (Table 5), despite the higher stage IV ratio of the AT haplotype carriers (12.7% of stage IV vs. 6.1% of stage III).
II versus III versus IV, p = 0.017, II versus IV, p = 0.012, II versus III, p = 0.405, III versus IV, p = 0.027.
Alc, alcohol induced; Ster, steroid induced; Idio, idiopathic.
χ
Serum lipid levels and their correlation to the IGFBP3 gene polymorphism
The serum level of TG and LDL-c of ONFH group was significantly increased (p = 0.01, p = 0.005, respectively), but the HDL-c level was statistically decreased (p = 0.0001) compared with the control group. Correlation analysis between the IGFBP3 genotypes and serum lipid levels in the ONFH group failed to reveal statistical difference, while the results in the control group showed that the HDL-c levels of the AG genotype carrier of rs3110697 were significantly higher compared with the AA genotype carrier (p < 0.05). The serum TC level of the CT genotype carrier of rs2453839 was statistically enhanced compared with the CC genotype carrier (p < 0.05), but HDL-c levels of the CC genotype carrier of rs2453839 were significantly decreased compared with the CT genotype carrier (p < 0.01), as shown in Figure 1.
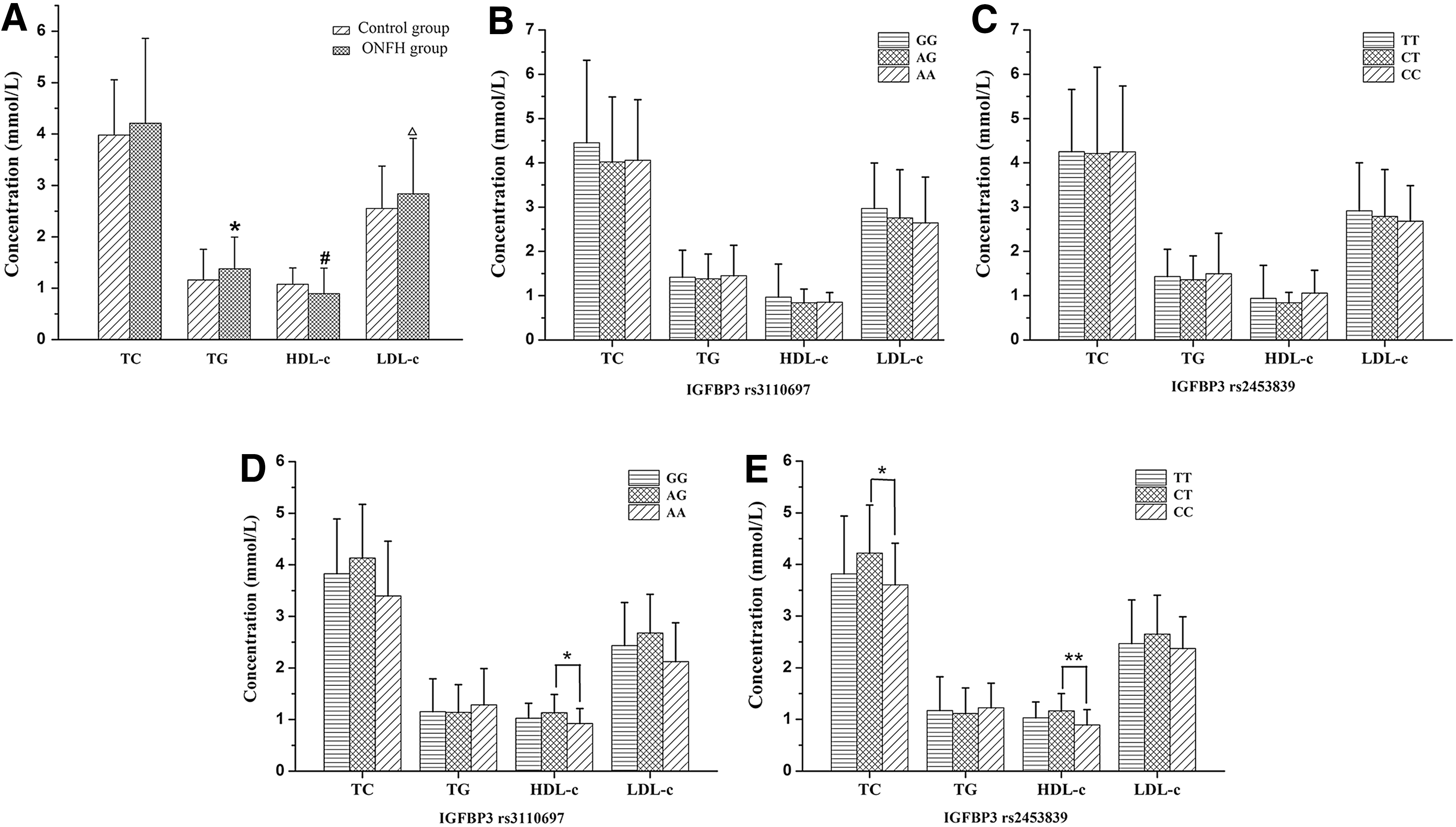
The serum lipid levels and the association of IGFBP3 genotypes with serum lipid levels.
The serum protein levels of IGFBP3 and IGF1 and their association with the gene polymorphism of IGFBP3 and the clinical phenotypes of ONFH
The serum IGFBP3 level, as well as the IGF1 level of the ONFH group, was significantly increased compared with the control group (p = 0.044, p = 0.007), respectively. Correlation analysis of the ONFH group revealed that the serum IGFBP3 level of TT or CT genotype carriers of rs2453839 was statistically higher compared with CC genotype carriers (p = 0.019, p = 0.007, respectively), while the serum IGF1 level failed to reveal the association with the IGFBP3 genotypes. The analysis results of the control group showed that the serum IGFBP3 level of TT genotype carriers of rs2453839 was significantly increased compared with CT genotype carriers (p = 0.035). Interestingly, the serum IGF1 level of TT genotype carriers of IGFBP3 rs2453839 was also statistically increased compared with the CT-type carriers (p = 0.034), as shown in Figure 2. The analysis results between the levels of IGFBP3 and IGF1 and the clinical phenotypes of ONFH further revealed that the serum IGF1 level of the patients with bilateral hip lesions was significantly lower compared with the patients with unilateral hip lesions (p = 0.047), as shown in Figure 3.
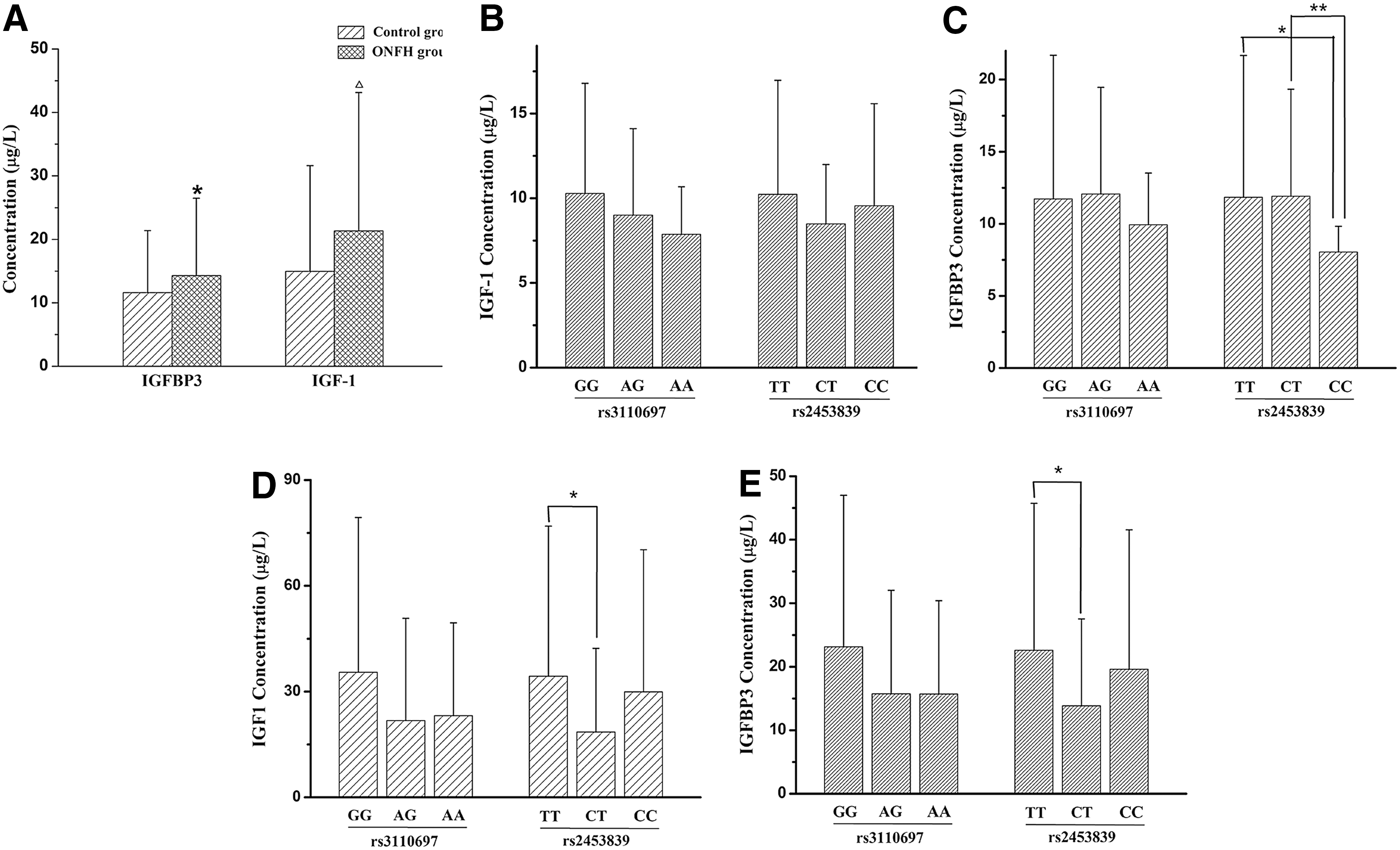
The protein expression of IGFBP3 and IGF1 in serum and the association of the expression with IGFBP3 genotypes.
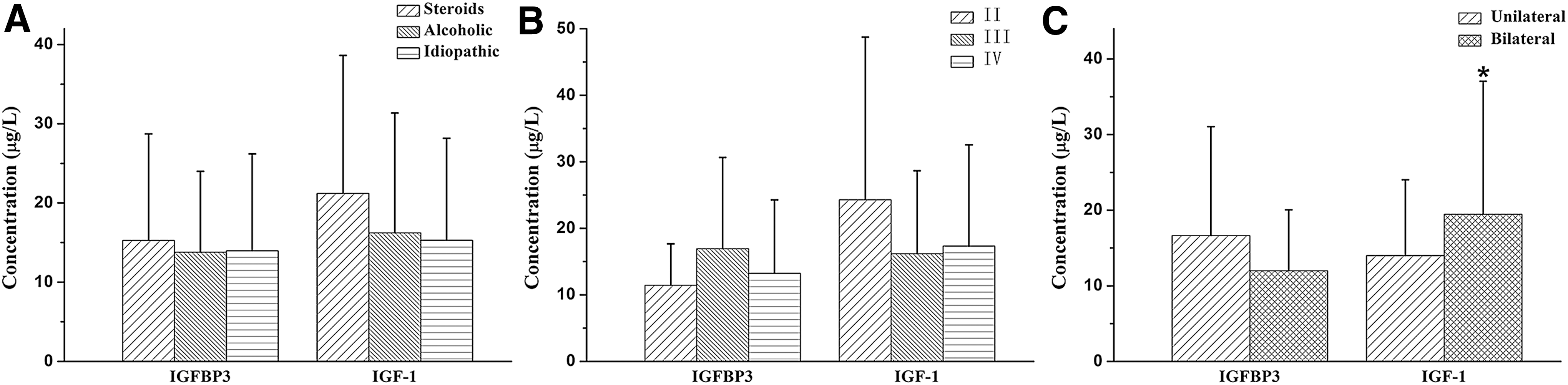
Association of serum protein expression of IGFBP3 genes with the clinical phenotypes of ONFH.
Discussion
IGFBP3 is a 39–42 kD binding protein of growth hormone dependence and the main carrier of serum IGF1 and stabilizes the local IGF1 concentration as its storage pool in blood circulation (Ceda, 1995; Jones and Clemmoner, 1995). IGFBP3 inhibits IGF1 activity by competitively binding to the ligand (Yamada et al., 2010). The critical roles of IGFBP3 in the regulation of bone metabolism were initially discovered in the research of vitamin D receptor (VDR). IGFBP3 and VDR are located in the nucleus and inhibit the transcriptional regulation of VDR induced by 1,25-(OH)2D3 and the activity of alkaline phosphatase (ALP) during osteoblast differentiation. After silencing of the VDR gene, IGFBP3 had no effect on ALP activity induced by 1,25-(OH)2D3, indicating a new regulatory mechanism of osteoblast differentiation by IGFBP3 (Chan et al., 2009). A recent study shows that the adenovirus vector containing IGFBP3 complementary DNA may inhibit the activity of nuclear factor-k-gene binding (NF-kB), production of chemokines, and secretion of matrix metalloproteinases in cultured fibroblast-like synovial cells and an arthritis mouse model induced by collagen (CIA), respectively, and decreased the severity of arthritis and pathological changes of CIA model mice, suggesting that IGFBP3 may reduce inflammatory bone lesions (Lee et al., 2014).
In view of the important effects of IGFBP3 on bone and joint diseases, we analyzed the association of the genotypes, allele frequencies, and haplotypes of rs2453839 and rs3110697 with the development of ONFH. The results showed that both the recessive model of rs3110697 and dominant model of rs2453839 were significantly associated with an increased risk of ONFH. Moreover, these results obtained from a Chinese Han population were also consistent with the results obtained from a Korean population (Hong et al., 2010) and further demonstrated that the polymorphism of the IGFBP3 gene closely related to the risk of ONFH. Furthermore, to explore the polymorphism roles of rs3110697 and rs2453839 in the development of ONFH, we performed a correlation analysis between the polymorphisms of the IGFBP3 gene and the clinical phenotypes of ONFH.
The results revealed that the genotypes of rs2453839 were associated with the clinical stages of ONFH (p = 0.017); that is, the CT genotype of rs2453839 might be a risk genotype of hip lesion progression of ONFH, while the CC and TT genotypes of rs2453839 might be protective genotypes against the hip lesion progression. Thus, these results indicated the risk and protective genotypes related to the hip lesion progression of ONFH. Our previous results revealed the correlation between the rs2453839 polymorphism and the clinical phenotypes of ONFH in a smaller 49 ONFH patients and 42 health controls (Song et al., 2012). To further validate the previous results, we reanalyzed the association of the rs2453839 genotypes with the clinical phenotypes of ONFH in an expanded system of 182 ONFH patients and 179 health controls, which did not contain the previous individuals of 49 ONFH patients and 42 health controls. The present results further confirmed that the rs2453839 genotypes of IGFBP3 gene associated with the clinical stages of ONFH. However, 182 ONFH patients and 179 health control system also attribute to a smaller sample system, especially for the subgroup analysis between the genotypes and clinical phenotypes. Therefore, the association of the rs2453839 genotypes with the clinical stages of ONFH still needs to be deeply investigated in the larger sample system.
To clarify the effect of IGFBP3 gene polymorphism on its expression, we detected the serum protein expression of IGFBP3 as well as IGF1 closely related to IGFBP3 function in 182 patients and 179 controls. The results showed that both serum IGFBP3 and IGF1 level of the ONFH group were significantly increased compared with the control group (p = 0.044, p = 0.007). Previous studies have reported that elevated serum IGFBP3 levels were associated with ONFH risk (Hong et al., 2010), and our results further confirmed that the increased IGFBP3 as well as IGF1 levels were closely related to ONFH risk. A correlation analysis in the ONFH group further demonstrated that the serum IGFBP3 level of rs2453839 TT or CT genotype carriers was significantly higher than the CC genotype carriers, indicating the genotypic effects on IGFBP3 gene expression. In addition, we also analyzed the association of IGFBP3 gene polymorphisms and serum IGFBP3 and IGF1 levels with the clinical phenotypes of ONFH, and the results demonstrated that serum IGF1 levels in patients with bilateral hip lesions were significantly decreased compared with patients with unilateral hip lesions, suggesting the possible role of IGF1 in the hip lesion progression of ONFH.
IGF1 regulates osteoblast function in autocrine and paracrine behavior and plays important roles in proliferation and differentiation of osteoblast and formation of the bone matrix (Feng et al., 2014). The biological relevance between calcium and IGF1 has been reported to associate bone metabolism. IGF1 is mainly produced by bone cells, and the lack of IGF1 in bone cells has been shown to impair bone development and bone turnover (Sheng et al., 2013). IGF1 directly binds to calcium channels to open the calcium reservoir (Sheng et al., 2014). Multiple evidence sources have indicated the complex interactions between IGF1 and calcium homeostasis and their regulation (Wong et al., 2006). Our results showed that both the IGFBP3 and IGF1 levels in serum of ONFH patients were significantly increased as well as closely associated with the clinical phenotypes of ONFH, indicating their possible roles in the development of ONFH.
The rs3110697 and rs2453839 are located in the intron3 and intron4 region of IGFBP3 gene, respectively. The effect of the two SNPs on serum IGFBP3 protein expression has not been reported, while other SNPs of IGFBP3 gene affecting the serum expression were more involved in cancers. A genome-wide association study (GWAS) of 10,280 individuals confirmed a known association of SNPs in the IGFBP3 gene region on chromosome 7p12.3 with serum IGFBP3 concentration using a significance threshold of p < 5 × 10−8 (Kaplan et al., 2011). The correlation investigation of the −202 A/C SNP of IGFBP3 gene with IGFBP3 serum levels and risk of prostate cancer in a Korean population showed that lower serum IGFBP3 levels were associated with an increased number of C alleles (Park et al., 2010). The other result also demonstrated that the IGFBP3 gene −202 A allele is associated with higher IGFBP3 serum levels and better growth response to growth hormone treatment in patients with severe growth hormone deficiency (Costalonga et al., 2009). The mechanism of the IGFBP3 gene SNPs affecting serum IGFBP3 protein expression remains obscure. Generally, the SNPs in promoter, 3′, 5′ UTR (untranslated region), or open reading frame region, may upregulate and downregulate gene expression or protein function (Martin et al., 2012), while the SNPs in intron region may mainly involve in mRNA transcription and translation regulation and tissue-specific expression (Costa et al., 2016). The detailed mechanisms remain to be explored.
In addition to the regulation of bone metabolism, the important effects of IGFBP3 on adipogenic differentiation have been demonstrated in recent years. Lipid metabolism disorder has been recognized as one of the core pathogenesis of ONFH. Thus, we also analyzed the serum lipid levels and their association with the genotypes of rs3110697 and rs2453839 in the ONFH and control groups, respectively. The results showed that TG and LDL-c levels of ONFH group were significantly increased (p = 0.01, p = 0.005), but the HDL-c level was statistically decreased (p = 0.0001) compared with the control group, demonstrating significant lipid metabolism disorder of ONFH patients. The correlation analysis results of the control group further showed that the serum HDL-c levels of the AA genotype carriers of rs3110697 were significantly decreased compared with AG genotype carriers (p < 0.05) and the serum TC level of the CT genotype carriers of rs2453839 was significantly increased (p < 0.05) compared with the CC genotype carriers, while the serum HDL-c level of the CC genotype carriers was statistically decreased compared with the CT genotype carriers (p < 0.01).
These results further revealed that the gene polymorphism of IGFBP3 significantly associated with serum lipid levels. Considering the potential effects of ONFH relation factors on the analysis, we completed the correlation analysis between the genotypes and the serum lipid levels in the ONFH and control groups, respectively. The analysis results of the control group showed that the IGFBP3 genotypes were significantly associated the serum lipid levels, but the results of ONFH group failed to reveal the association; a possible factor may be related to the bigger standard deviation of lipid levels in ONFH group. The detailed mechanisms remain to be investigated.
In addition, the association of IGFBP3 genotypes with clinical phenotypes involved in the two–two comparison multiple test of three genotype subgroups. Generally, multiple comparisons may increase false positive. Especially the SNP number of a GWAS may be more than 1 million, and such a huge number of multiple comparisons may lead to many false positives. Multiple-test adjustment of the significance threshold may well decrease the false positive of GWAS (William, 2009). Considering this study only involving in two SNPs, instead of the adjustment of the significance threshold, we calculated the corresponding p-values of each SNP controlling for age and sex as covariates (Cui et al., 2014; Peng et al., 2014) or adopted the single factor variance analysis with the least significant difference (LSD) for the serum protein and lipid levels analysis among genotypes, which is considered as effectively controlling the fairway-wise error rate. However, the correlation between IGFBP3 gene polymorphism and clinical phenotypes needs to be further investigated on the basis of the adjustment of the significance threshold.
In conclusion, our results confirmed that both the gene polymorphisms and gene expression of IGFBP3 gene are not only significantly associated with ONFH risk but also closely related to the clinical phenotypes of ONFH. Specifically, we suggested the possible risk and protective genotypes involving in the hip lesion progression of ONFH, respectively. These findings first simultaneously showed the possible critical roles of the polymorphisms of IGFBP3 gene, the abnormal expression of IGFBP3 and IGF1 protein, and lipid metabolism disorder in a case–control comparison in the development of ONFH.
Footnotes
Acknowledgments
The authors appreciate the support of all of the surgeons in the Department of Orthopedics who were involved in patient care and control and all of the graduated students in The Research Center who were involved in the genomic DNA extraction. This work was supported by the Project of Health Management Department of Jilin Province, China (Grant No. 20132003), the Project of Bethune Youth Foundation of Jilin University, China (Grant No. 2015409), the Department of Science and Technology of Jilin Province, China (Grant No. 20130727038YY, 20140311006YY, 20150312022ZG), and the Development and Reform Commission of Jilin Province, China (Grant No. 2014G073, the project of application demonstration center of precision medicine for molecular diagnosis in Jilin Province [2016–2018]).
Disclosure Statement
No competing financial interests exist.