
Abstract
Even though the functions of the klotho gene in aging of small animals such as mice have been well investigated, studies using large animal models such as pigs, which have many similarities to humans, have been limited due to the absence of cell lines or animal models. Therefore, the objective of this study was to generate porcine cell lines overexpressing human klotho (hKlotho) and tetracycline (Tet)-inducible hKlotho and to produce cloned embryos from these cell lines. We designed vectors for hKlotho overexpression (CA-Klotho) under control of CMV enhancer/chicken β-actin (CAG) promoter and Tet-inducible hKlotho overexpression (Tet-Klotho, under control of doxycycline-dependent promoter). The vectors were transfected into porcine fibroblasts then CA-Klotho and Tet-Klotho cell lines were established. The Tet-Klotho (+) cell line was cultured in the presence of doxycycline (2 μg/mL), whereas the Tet-Klotho (−) cell line was cultured without doxycycline. In polymerase chain reaction (PCR) and reverse transcription-PCR (RT-PCR) assays, integration and expression of the hKlotho gene were confirmed in CA-Klotho, Tet-Klotho (+), and Tet-Klotho (−) cell lines. The CA-Klotho cell line was subjected to real-time PCR and showed positively changed expression of genes related to aging and cell survival. Somatic cell nuclear transfer was performed to generate hKlotho overexpression cloned embryos by using CA-Klotho and Tet-Klotho (+) cell lines; blastocyst formation frequency was significantly higher in cloned embryos from CA-Klotho and Tet-Klotho (+) (21.5% and 20.2%, respectively) compared with the control (8.4%). In conclusion, we established hKlotho overexpression and Tet-inducible hKlotho overexpression cell lines and porcine embryos cloned from these cell lines.
Introduction
A
In contrast, overexpression of klotho in mice prolongs lifespan by 20% (Kurosu et al., 2005). Moreover, inducible klotho gene expression in klotho-deficient mice (KL−/− mice) rescues all the aging-like phenotypes generated before induction of the exogenous klotho gene (Masuda et al., 2005). In this regard, the klotho gene is considered to be one of the aging-suppressor genes that prolongs lifespan when overexpressed. Although the mechanism of the anti-aging action of klotho is not known exactly, one solid hypothesis is that the ability of klotho to inhibit insulin-like growth factor-1 (IGF1) signaling may contribute to the anti-aging properties of klotho, since numerous studies have indicated that moderate inhibition of IGF1 signaling is one of the anti-aging mechanisms (Tatar et al., 2003). In previous studies, inhibition of IGF1 signaling increased the lifespan of several animals, including Caenorhabditis elegans (Apfeld and Kenyon, 1998), Drosophila (Tatar et al., 2001), and mice (Holzenberger et al., 2003). In particular, transgenic mice overexpressing klotho showed suppressed oxidative stress together with moderately inhibited IGF1 signaling (Yamamoto et al., 2005).
The antioxidant effect of klotho may be attributed to Forkhead box O (FOXO) transcription factors, such as FOXO1, which are downstream targets of IGF1 signaling. In the absence of klotho, IGF1 signaling represses FOXO1 so that no downstream signaling occurs, whereas in the presence of klotho, the suppression of FOXO1 is lifted, leading to the activation of downstream target genes regulating antioxidant function, including catalase (CAT) and mitochondrial manganese superoxide dismutase (MnSOD) (Malik and Storey, 2011). In summary, inhibition of IGF1 signaling by klotho can activate FOXO1 repressed by IGF1 signaling under normal conditions, and increased activation of FOXO1 may upregulate antioxidant genes potentially contributing to anti-aging characteristics.
Many studies have been conducted to identify the functions of the klotho gene in aging of small animals such as mice. However, limited information is available on the functions of the klotho gene in large animals, such as pigs, which are widely used as models for studying human diseases due to their anatomical and physiological similarities to humans (Prather et al., 2013). The aim of this study, therefore, was to generate transgenic porcine fibroblasts and preimplantation embryos expressing a human klotho (hKlotho) gene to produce a more suitable animal model for elucidating the role of this gene in aging.
Materials and Methods
Primary culture of porcine fetal fibroblasts
Male fetal fibroblasts from one porcine fetus were isolated and cultured. The euthanized fetus was dissected into three parts: head, body, and tail. The fetal body parts were washed three times in phosphate-buffered saline (PBS) and then chopped into small pieces in a 60-mm dish with trypsin. Trypsinized tissues were then incubated for 30 min at 37°C. Well-dissociated tissues were centrifuged at 1500 g for 2 min. The supernatant was discarded, and the pellet was resuspended with PBS, and then centrifuged at 1500 g for 2 min. These procedures were repeated twice. Finally, the supernatant was discarded, and the pellet was resuspended in Dulbecco's modified Eagle's medium (DMEM; Gibco, Carlsbad, CA) supplemented with 15% fetal bovine serum (FBS; Gibco), 1% penicillin/streptomycin (P/S; Gibco), 1% nonessential amino acids (NEAA; Gibco), and 100 mM β-mercaptoethanol (β-ME), by inverting the tube several times. The cells resuspended in this medium were held at room temperature (∼25°C) for 5 min, and then the suspension was transferred into a cell culture dish for ∼10 days with the culture medium being changed every 2–3 days. These primary cells were cultured, expanded, and finally frozen at −196°C for further use. The cell cultures were maintained in DMEM with 15% FBS, 1% P/S, 1% NEAA, and 100 mM β-ME.
Generation of the hKlotho overexpression cell line
hKlotho (from Addgene,
Generation of tetracycline (Tet)-inducible hKlotho overexpression cell line
hKlotho was linked to an mCherry with F2A sequences by PCR amplification, and inserted into the piggybac transposon expression vector (pB-TET, Plasmid #20909; Addgene) using the In-Fusion Cloning Kit (Clontech). After the transfection of 1000 ng/μL of pB-Tet-hKlotho-F2A-mCherry, transposase (named pCy43, provided by The Sanger Institute), and pB-CAG-rtTA into 5 × 105 of porcine fibroblasts through nucleofection (Neon; Invitrogen) with a 1400 V, 20 ms pulse width, and pulse number 2 setting, 1000 μg/mL neomycin (G418; Gibco) was used for 7 days to isolate the transfected cells, and then growing cells resistant to neomycin were subcultured. This cell line was named Tet-Klotho (+) when treated with doxycycline and Tet-Klotho (−) when left untreated.
Polymerase chain reaction
Genomic DNA was extracted with the G-spin Genomic DNA Extraction Kit (iNtRON Biotechnology, Gyeonggi-do, Korea) according to the manufacturer's protocol. Amplification of target genes was achieved using Maxime PCR PreMix (i-StarTaq; iNtRON Biotechnology). The PCR reactions were denatured at 95°C for 20 s, annealed at 60°C for 10 s, and extended at 72°C for 30 s. A primer set for hKlotho (forward primer: 5′-TGGACCCACCTTGAGTTTTC-3′ and reverse primer: 5′-GGAGGGAAGCCATTTTTCTC-3′, 411 bp) was used.
RT-PCR
Total RNAs were isolated by the easy-spin Total RNA Extraction Kit (iNtRON Biotechnology) according to the manufacturer's instructions. To detect transcripts of hKlotho, the total RNAs were isolated and added to the Maxime RT Premix Kit [oligo (dT) primer; iNtRON Biotechnology] for the synthesis of complementary DNAs (cDNAs), and then the synthesized cDNAs were used as reverse transcription (RT)-PCR templates. Plasmid vector (pB-CA-hKlotho-F2A-mCherry) were used as a positive control for hKlotho. Primer sets for transcript of hKlotho (forward primer: 5′-TGGACCCACCTTGAGTTTTC-3′ and reverse primer: 5′-GGAGGGAAGCCATTTTTCTC-3′, 411 bp) and β-actin (forward primer: 5′-GTGGACATCAGGAAGGACCTCTA-3′ and reverse primer: 5′-ATGATCTTGATCTTCATGGTGCT-3′, 137 bp) were used.
Somatic cell nuclear transfer
Donor cells were subjected to somatic cell nuclear transfer (SCNT), which was performed as described in a previous report (Lee et al., 2016). In brief, in vitro matured pig oocytes were enucleated by aspirating the first polar body and the adjacent cytoplasm containing chromosomes with an aspiration pipette. Then, using a fine pipette, a trypsinized porcine fetal fibroblast, with a smooth cell surface, was transferred into the perivitelline space of each enucleated oocyte. These couplets were electrically fused with a single DC pulse of 200 V/mm for 30 μs using an electro cell fusion generator (LF101; Nepa Gene Co., Japan). Then, 30 min after fusion, fused couplets were activated with a single DC pulse of 1.5 kV/cm for 60 μs using a BTX Electro-Cell Manipulator 2001 (BTX, Inc., San Diego, CA). The resulting activated embryos were cultured in Porcine Zygote Medium-5 (PZM-5; Funakoshi Corporation, Tokyo, Japan) for 7 days. Cleavage and blastocyst formation frequencies were observed on days 2 and 7 after culture, respectively.
Quantitative PCR
All samples were stored at −80°C until analysis. Total RNA was extracted using the easy-spin Total RNA Extraction Kit (iNtRON Biotechnology), according to the manufacturer's protocol, and the total RNA concentration was quantified using a NanoDrop 2000 Spectrophotometer (Thermo Fisher Scientific, Wilmington, DE). cDNA was synthesized using Maxime RT Premix (iNtRON Biotechnology) according to the manufacturer's protocol. A PCR plate (MicroAmp optical 96-well reaction plate, Applied Biosystems, Singapore) was made by adding 1 μL cDNA, 0.4 μL (10 pmol/μL) forward primer, 0.4 μL (10 pmol/μL) reverse primer, 10 μL SYBR Premix Ex Taq (TaKaRa, Otsu, Japan), and 8.2 μL of Nuclease-free water (Ambion, Austin, TX). The reactions were carried out for 40 cycles and the cycling parameters were as follows: denaturation at 95°C for 15 s, annealing at 60°C for 1 min, and extension at 72°C for 1 min. All oligonucleotide primer sequences are presented in Table 1. GAPDH was selected for an internal control because similar CT values were obtained between control and CA-Klotho groups The expression of each target gene was quantified relative to that of the internal control gene (GAPDH) using the equation, R = 2−[ΔCt sample − ΔCt control].
Statistical analysis
All data were derived from experiments repeated at least three times (biological replication for percentage data (e.g., frequencies of cleavage and blastocyst formation) and technical replication for data on gene expression) and analyzed by one-way ANOVA followed by Tukey's multiple comparison test or paired t-test using SPSS 22.0 (SPSS, Inc., Chicago, IL) to determine differences among experimental groups. Percentage data were arcsine-transformed before analysis to maintain normality and homogeneity of variances. Statistical significance was determined when the p-value was <0.05.
Results
Establishment of CA-Klotho and Tet-Klotho cell lines
The hKlotho-F2A-mCherry transgene was constructed under the control of the CMV enhancer/chicken β-actin (CAG) promoter (Fig. 1A). The constructed transgene was introduced into porcine fetal fibroblast cells using a nucleofection gene delivery system. After transfection, the mCherry-positive cells were mechanically isolated and subcultured. Establishment of the CA-Klotho cell line was further confirmed by mCherry expression observed with fluorescence microscopy (Fig. 2B, B′).

A schematic diagram of constructs used in the study. hKlotho was linked to an mCherry with F2A sequences by PCR amplification, and inserted into the piggybac transposon expression vector (pB-CA or pB-Tet) by In-Fusion cloning.

hKlotho-F2A-mCherry expression in CA-Klotho, Tet-Klotho (+), and Tet-Klotho (−) cell lines.
To establish the Tet-inducible hKlotho cell line (Tet-Klotho), porcine fetal fibroblasts were transfected with a vector containing hKlotho, whose expression was controlled by doxycycline (Fig. 1B). The transfected fibroblasts were selected with medium containing neomycin, which was further confirmed by mCherry expression with (Fig. 2C, C′) or without doxycycline (Fig. 2D, D′).
PCR and RT-PCR
Integration and expression of the hKlotho gene were observed by genomic DNA PCR (Fig. 3A) and RT-PCR (Fig. 3B) in control, CA-Klotho, Tet-Klotho (+), and Tet-klotho (−) cells, respectively. PCR and RT-PCR data indicated that the hKlotho gene was integrated into porcine fetal fibroblasts. We compared transcript expression levels of hKlotho measured by RT-PCR (Fig. 3C) and the CA-Klotho group displayed the highest hKlotho expression (p < 0.05) compared with the other groups. In the Tet-Klotho (+) group treated with doxycycline, expression of hKlotho was significantly increased compared with Tet-Klotho (−) cells.
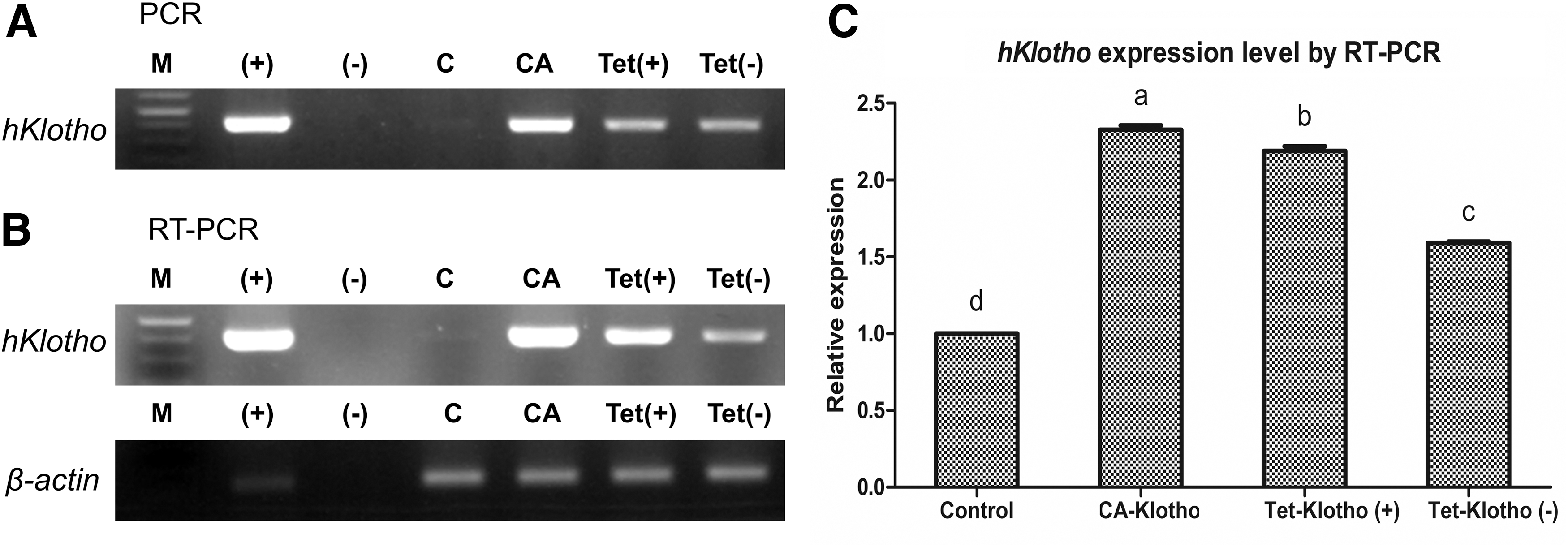
PCR and RT-PCR results for transfected cells.
Effect of hKlotho overexpression on gene expression
Gene expression in CA-Klotho cells and control cells is summarized in Figures 4 and 5. In this analysis, expression of genes related to aging (IGF1 signaling genes, FOXO1, and antioxidant genes) and cell survival (tumor suppressor genes, DNA methylation genes, and apoptotic genes) was evaluated.
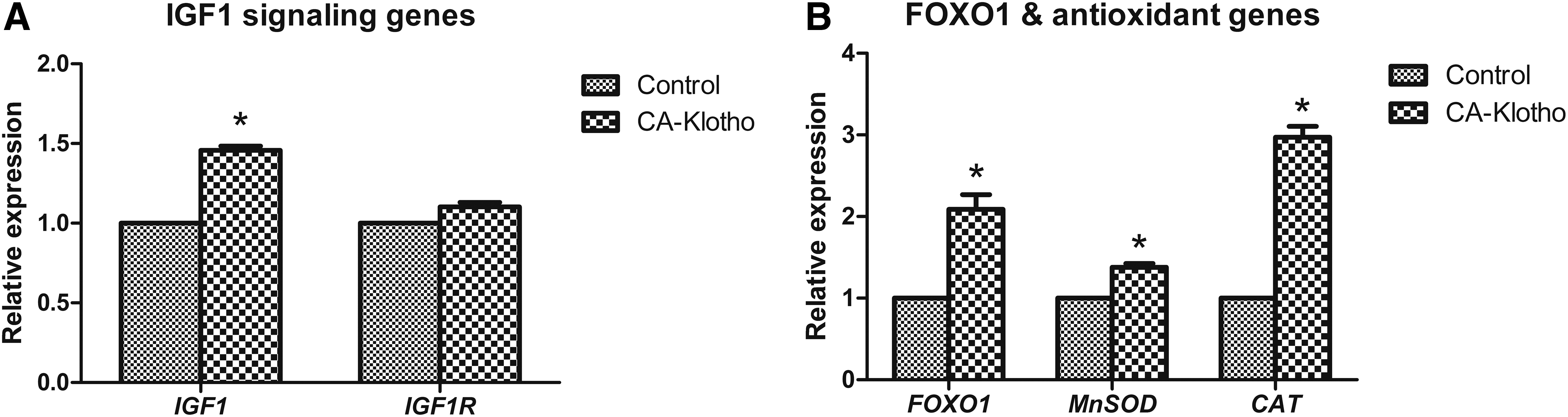
Aging-related gene expression in CA-Klotho cell line.
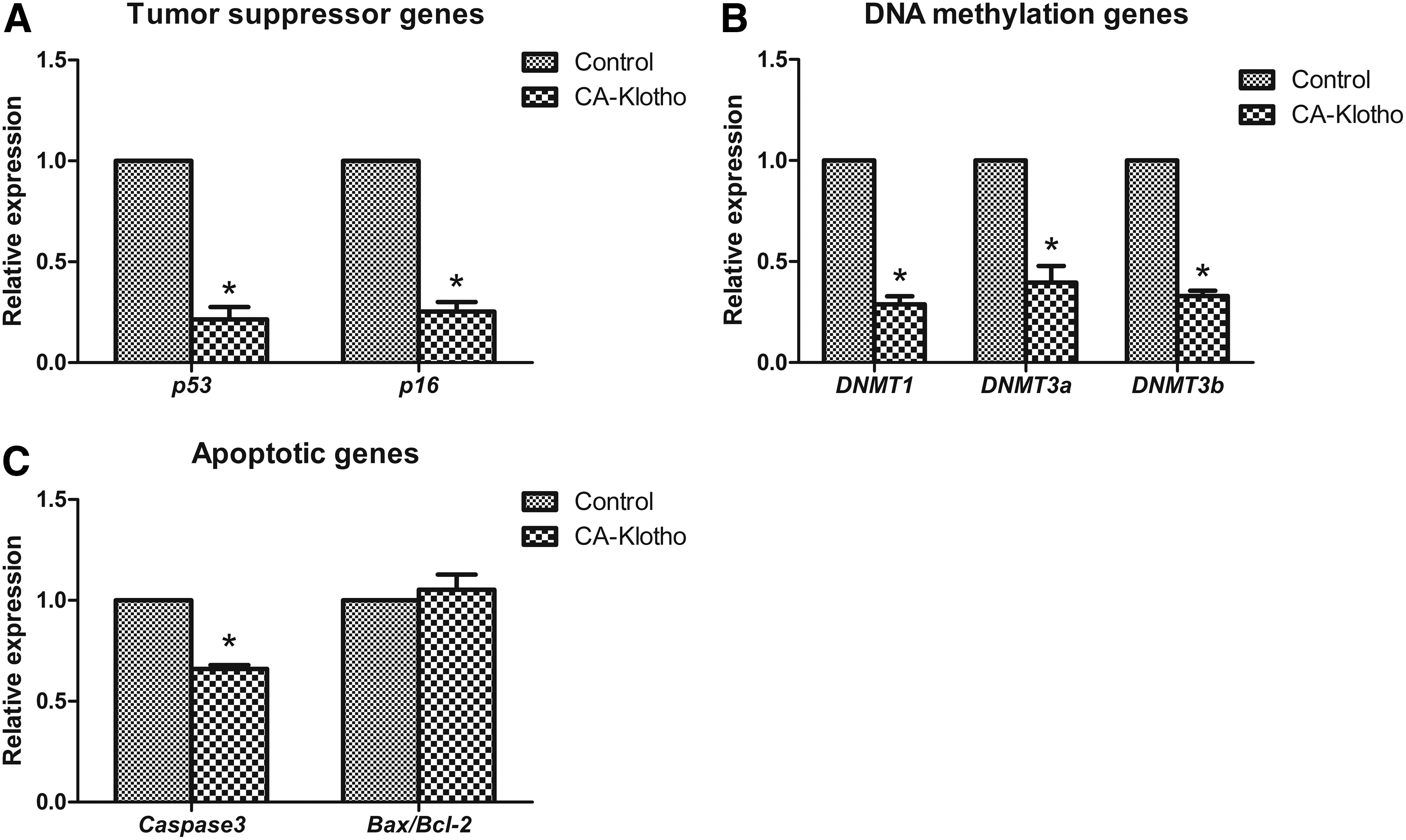
Cell survival-related gene expression in CA-Klotho cell line.
Regarding aging-related genes, transcripts of IGF1 were significantly increased in the CA-Klotho group. However, no significant difference in expression of the IGF1 receptor (IGF1R) was observed between the control and CA-Klotho group. The CA-Klotho group significantly increased transcripts of FOXO1 and its downstream target genes with antioxidant function (MnSOD and CAT).
In terms of cell survival-related genes, the tumor suppressor gene (p53 and p16) expression level was significantly reduced in CA-Klotho cells compared with control cells. Expression of DNA methylation genes (DNA methyltransferase (DNMT)1, DNMT3a, and DNMT3b) and proapoptotic gene (Caspase3) was significantly downregulated in CA-Klotho cells. However, the Bax/Bcl-2 ratio, an indicator of apoptosis, was not significantly changed between CA-Klotho and control cells.
Preimplantation development of cloned embryos derived from CA-Klotho and Tet-Klotho (+) cell lines
Development frequencies were evaluated in three groups: SCNT-derived embryos using control cells, CA-Klotho cells, and Tet-Klotho (+) cells as nuclear donors. Two days after activation, cleavage frequencies evaluated under a microscope were 78.5% ± 5.9%, 80.5% ± 4.6%, and 77.3% ± 2.8%, respectively. No significant differences were observed in cleavage frequencies among the three groups. However, significant differences were observed in blastocyst formation frequencies on day 7, which were 8.4% ± 1.5%, 21.5% ± 3.9%, and 20.2% ± 4.0%, respectively (Fig. 6).
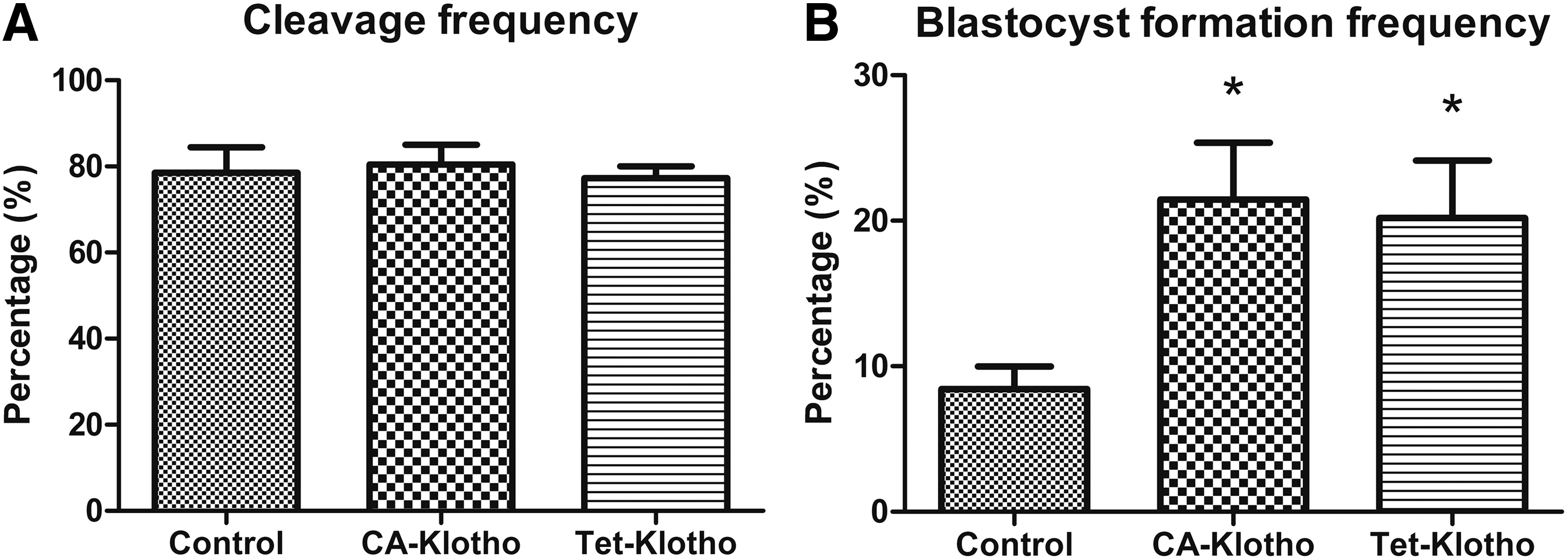
Development frequency of embryos cloned from control, CA-Klotho, and Tet-Klotho (+) cell lines. Cleavage
Discussion
In small animals such as mice, it has been reported that the klotho gene has an important role in suppressing aging by regulation of some signaling pathways. One of the important pathways regulated by klotho is IGF1 signaling. Klotho suppresses IGF signaling by inhibiting IGF1-induced autophosphorylation of the IGF1 receptor and subsequently it relieves the inhibitory effect of IGF1 signaling on FOXO1, an essential regulator of antioxidant gene expression. Activated FOXO1 triggers activation of the antioxidant genes, including CAT and MnSOD, which may exert the anti-aging functions of klotho. However, this information is limited to small animals. Therefore, in the present study, we generated a porcine fibroblast cell line expressing the hKlotho gene to produce a suitable animal model for studying human aging.
We established two cell lines: one is the CA-Klotho cell line, which overexpresses hKlotho under the control of the CAG promoter, and the other is the Tet-Klotho, which is doxycycline-inducible hKlotho overexpression cell line. Although doxycycline was used, the Tet-Klotho (+) cell line expressed significantly lower hKlotho than CA-Klotho. This was confirmed by the observation of mCherry expression with a fluorescent microscope (Fig. 2) and comparison of transcript levels of hKlotho by RT-PCR (Fig. 3). The reason why mCherry expression can confirm the expression of hKlotho is because hKlotho is directly connected to mCherry by the F2A sequence, and thus they express the same level of hKlotho.
One disadvantage of tetracycline-inducible systems is their well-described leakiness (Hoppe et al., 2014), which may be attributed to chromosomal integration, false promoters, or cryptic initiation signals under noninduced conditions (Garrick et al., 1998; Johansen et al., 2002). Considering the level of hklotho in Tet-Klotho (−) cell line was significantly higher than the control group (Fig. 3), our inducible system also seem to be somewhat leaky.
For investigating the properties of klotho overexpression cells at the molecular level, the expression level of several genes related to aging and cell survival was analyzed using control and CA-Klotho cells. CA-Klotho had a significantly higher expression level of IGF1, but not of IGF1R. This may be due to the ability of klotho to inhibit IGF1-induced autophosphorylation of IGF1 receptors. Inhibited signaling by klotho may induce expression of IGF1 to compensate for IGF1 resistance. This result is consistent with a previous study, which showed that treatment of mice with intraperitoneal klotho injections increased mRNA levels of IGFI in the liver (Shahmoon et al., 2014). CA-Klotho had also significantly higher FOXO1 and its downstream target genes with antioxidant function (MnSOD and CAT) than the control. This can be explained by evidence that inhibition of IGF1 signaling allows FOXOs to positively regulate the expression of anti-aging genes and negatively regulate the expression of proaging genes (Lee et al., 2003; Murphy et al., 2003). Based on these results, overexpression of klotho seems to exert anti-aging properties through inhibition of IGF1 signaling. In analysis of cell survival-related genes, CA-Klotho had significantly reduced tumor suppressor genes (p53 and p16), DNA methylation genes (DNMT1, DNMT3a, and DNMT3b), and Caspase3, which plays an important role in preventing cellular senescence. Tumor suppressor genes (p53 and p16) have crucial roles in regulating cellular senescence and cell-cycle arrest. When cells enter into senescence, expression of p53 and p16 is increased (Campisi, 2001). Therefore, decreased p53 and p16 reflect improved cell survival. Among the DNMTs, there are three well-characterized DNMTs: DNMT1, which is responsible for maintenance of methylation, and DNMT3a and DNMT3b, which are responsible for mediation of de novo methylation (Okano et al., 1998). DNA methylation in gene promoters decreases gene expression, by alterations of local DNA structure and prevention of transcription factor binding (Boyes and Bird, 1991). In other words, DNA hypomethylation by decreasing DNMTs is required for reactivation of specific genes associated with cell survival (Rojas and Galanti, 1991). Therefore, decreased DNMTs also represent improvement of cell survival. In addition, decreased apoptosis was reflected in downregulated Caspase3, one of the proapoptotic genes. In summary, klotho overexpression positively changed expression of cell survival-related genes.
The klotho overexpression cell lines were employed in porcine SCNT to compare embryo development frequencies. Embryos cloned from CA-Klotho and Tet-Klotho (+) cell lines had significantly higher blastocyst formation frequency than the control. Considering CA-Klotho cell line expressed significantly higher hKlotho than Tet-Klotho (+) and blastocyst formation frequency was numerically higher in the CA-Klotho versus Tet-Klotho (+), there is possibility that hKlotho expression influences blastocyst formation frequency dose dependently. However, blastocyst formation frequencies between the groups were not significantly different and further studies are required to elucidate whether hKlotho expression affects blastocyst formation frequency in a dose-dependent manner. In previous studies, it was concluded that SCNT using donor cells with DNA hypomethylation improves embryo development (Xiong et al., 2013), because proper DNA methylation patterns are essential for embryonic development (Robertson, 2002). Although we did not measure the level of DNA methylation, we speculate that decreased DNMTs caused by klotho overexpression in our donor cells might affect their DNA methylation levels and so result in higher embryo development. Compared with our previous studies (Lee et al., 2015, 2016), low blastocyst formation frequency of the control group in this study (<10%) may be due to seasonal effect. It is well known that developmental competence of bovine and porcine embryos was reduced in summer season (Al-Katanani et al., 2002; Suzuki et al., 2010), in which we performed SCNT.
We established hKlotho-overexpressed and inducible hKlotho-overexpressed fibroblast cell lines that may be useful cell sources for SCNT procedures to generate hKlotho overexpression pigs. In addition, we found that overexpression of hKlotho upregulates antioxidant genes (MnSOD and CAT), which are downstream target genes of FOXO1. This phenomenon seems to be related to activation of FOXO1 through inhibition of IGF1 signaling by klotho and can explain the anti-aging properties of klotho. Collectively, we may predict that hKlotho overexpression pigs will exhibit an extended lifespan with altered IGF1 signaling, FOXO1 and subsequent antioxidant gene expression and will be a good model for studying aging of humans. However, these predictions were based on cellular level, and further studies are required to generate hKlotho overexpression pigs.
Footnotes
Acknowledgments
We thank Dr. Goo Jang (Seoul National University, Korea) for providing piggybac transposon expression vectors. This study was supported by the Korean Institute of Planning and Evaluation for Technology in Food, Agriculture, Forestry and Fisheries (#114059-03-2-SB010, #311011-05-5-SB010), the Research Institute for Veterinary Science, TS Corporation, and the BK21 plus program.
Disclosure Statement
No competing financial interests exist.