
Abstract
Hepatitis C virus (HCV) can infect extrahepatic tissues, including lymphocytes, creating reservoir of the virus. Moreover, HCV proteins can interact with DNA damage response proteins of infected cells. In this article we investigated the influence of the virus infection and a new ombitasvir/paritaprevir/ritonavir ± dasabuvir ± ribavirin (OBV/PTV/r ± DSV ± RBV) anti-HCV therapy on the PBMCs (peripheral blood mononuclear cells, mainly lymphocytes) DNA base excision repair (BER) system. BER protein activity was analyzed in the nuclear and mitochondrial extracts (NE and ME) of PBMC isolated from patients before and after therapy, and from subjects without HCV, using modeled double-strand DNA, with 2′-deoxyuridine substitution as the DNA damage. The NE and ME obtained from patients before therapy demonstrated lower efficacy of 2′-deoxyuridine removal and DNA repair polymerization than those of the control group or patients after therapy. Moreover, the extracts from the patients after therapy had similar activity to those from the control group. However, the efficacy of apurinic/apyrimidinic site excision in NE did not differ between the studied groups. We postulate that infection of lymphocytes by the HCV can lead to a decrease in the activity of BER enzymes. However, the use of novel therapy results in the improvement of glycosylase activity as well as the regeneration of endonuclease and other crucial repair enzymes.
Introduction
E
Materials and Methods
Substrate oligonucleotides
The two 30-mer complementary oligonucleotides—the first containing 2′-deoxyuridine (dU) and the second unmodified—were purchased from CBMM PAN (Lodz, Poland). The modified oligonucleotide was labeled on the 5′ end by [γ32P]ATP using polynucleotide kinase (Karwowski et al., 2014). The sequences of the double-stranded oligodeoxynucleotides (ds-oligonucleotides) were as follows: d[AAA TTA ATA TGG TAT TGdU ATA TAA ATT ATT] and d[AAT AAT TTA TAT ACA ATA CCA TAT TAA TTT].
Preparation of an apurinic/apyrimidinic site
The ds-oligonucleotides that contained dU were treated with one unit of uracil-DNA glycosylase (UDG) (NEB) in 100 μL of buffer (10 mM Tris-HCl, pH 7.5, 50 mM NaCl, and 1 mM EDTA) for 30 min at 37°C to produce an apurinic/apyrimidinic (AP) site (Karwowski et al., 2014).
Preparation of NE and ME
PBMCs were isolated from blood samples by isopycnic centrifugation in Gradisol L (Aquamed, Poland). The cells were then fractionated to obtain nuclear and mitochondrial pellets using the Mitochondria Isolation Kit for Cultured Cells (Thermo Fisher Scientific). Both pellets were stored at −80°C. Immediately before DNA incision assay, the pellets were suspended in cold buffer containing 20 mM HEPES, 420 mM NaCl, 1.5 mM MgCl2, 0.2 mM EDTA, 0.5 mM DTT, 25% glycerol, and protease inhibitor. A Pierce™ Coomassie (Bradford) Protein Assay Kit (Thermo Fisher Scientific) was used to estimate the protein concentration in each extract.
Patients
Antiviral medication ombitasvir/paritaprevir/ritonavir (OBV/PTV/r) ± dasabuvir (DSV) ± ribavirin (RBV) were obtained from the manufacturer within the AMBER, named patient program, created to provide early access to patients with advanced liver disease and life-threatening course of HCV infection. These three patients were adults; one male and two female, with chronic HCV genotype 1 or 4 infection, with no previous antiviral treatment. Blood was taken from the patients before treatment and after 12 weeks. A control group was formed of subjects, which were found to be negative for HCV.
The dU excision assays
The ds-oligonucleotides containing dU (30 nmol) were incubated with 1 μg of NE or 2 μg of ME in 30 μL of reaction buffer (50 mM Tris-HCl, pH 7.5, 5 mM MgCl2, 20 mM NaCl, 1 mM DTT, 4 mM ATP) (Hou et al., 2007) at 37°C for 0, 15, 30, 60, and 120 min. To stop the reactions, 5 μL of denaturing stop solution (98% formamide, 2 mM EDTA, 0.025% bromophenol blue, and 0.025% xylene cyanol) was added. The samples were then subjected to electrophoresis on a 20% denaturing polyacrylamide gel (PAGE) containing 7 M urea in 1 × TBE (89 mM Tris-HCl, 89 mM boric acid, and 2 mM EDTA, pH 8.3) for 180 min at a constant power of 2000 W. The gel was exposed to a radiographic film for visualization of DNA, and quantified with Quantity One software (Bio-Rad). For the time-dependence study of the repair of investigated ds-oligonucleotide, the intensity of the bands representing either single-stranded DNA (ssDNA), or ssDNA with one, two, three, or four added bases was expressed as the percentage of the total intensities for all bands within each lane to overcome any differences in loading or in 32 P labeling.
The AP-site excision assays
The ds-oligonucleotides containing AP-site (30 nmol) were incubated with 0.5 μg of NE or a 1 μg of ME in 30 μL of the above-described reaction buffer at 37°C for 0, 1, 5, 15, 30, and 60 min. The subsequent procedures were the same as described in the dU excision assay.
The polymerization assay of NE supported by polymerase β
The ds-oligonucleotides containing an AP-site (30 nmol) were incubated with 2 μg NE and 1 U of polymerase β (polβ) (EURx, Poland) in 30 μL of the above-described reaction buffer with the addition of 20 mM of each dNTP (Hou et al., 2007) at 37°C for 0, 1, 5, 15, 30, and 60 min. The subsequent procedures were the same as described in the dU excision assay.
Results
Activity of UDG in NE and ME
The presence of dU in the investigated 30-mer double-stranded oligonucleotide was confirmed by one-pot digestion by UDG to create an AP-site, followed by a 60-min treatment with enzymes containing AP-site cleavage activity: human AP endonuclease 1 (APEX1), 8-oxoguanine DNA glycosylase (OGG1) or Nth (Fig. 2I), resulting in the appearance of a shorter oligonucleotide (17-mer). Incision of dU from the ds-oligonucleotides using the extracts was found to be significantly less effective in the case of patients before HCV therapy (ME = 11.5%; NE = 5%) than the patients after therapy (ME = 30.9%; NE = 33.5%) or control patients (ME = 48.8%; NE = 39.3%) (Figs. 2A, D and 3A, C).
Activity of APEX1 in NE and ME
The effectiveness of digestion of the enzymatically formed ds-oligonucleotide containing the AP-site by NE was revealed to be nearly the same for the patients before HCV therapy (70.9%), those after HCV therapy (86.2%), and the control group (97%) after 60 min (Fig. 2B, E). However, significant differences were found for the ME: before therapy—33.45%, after therapy—95.1%, and control group—100% (Fig. 3B, D).
Polymerase activity supported by the exogenous polymerase β
NE or ME alone could not facilitate the oligonucleotide elongation process since it was not measurable (data not shown). Thus, this analysis was performed with the addition of exogenous, isolated polβ (Fig. 2C, ssb +2, +3, +4) and it revealed that for the HCV-positive patients, polymerization activity reached a maximum (32%) after 15 min of incubation and increased only by 3% after another 45 min (Fig. 2H). In contrast, the post-HCV therapy and control patients demonstrated similar initial activity, but increased by 24% within the last 45 min (Fig. 2H). Moreover, a greater yield of the first nucleotide addition was observed for NE derived from control group PBMC after 30 min of incubation (51%), than patients before HCV therapy (34%, 5 min) or after therapy (40%, 15 min) (Fig. 2G). The presence of polβ and dNTP cause also a greater activity of AP-lyase after 60 min than in the previous experiment; however, the efficiency order retained the same (86.2%, 92.6%, and 97.5%) (Fig. 2F).
Discussion
The most frequent DNA lesions (8-oxoguanine, thymine glycol, dihydrothymine, dU) (De Bont and van Larebeke, 2014) are removed from the genome by the BER (Kim and Wilson, 2012) (Fig. 1). This repair system is initiated by specific glycosylases leading to short (SP) or long patch (LP) subpathways, which are differentiated by polymerase activity: in the SP, only one nucleotide is replaced by the correct one, whereas in the LP, two or more nucleotides are incorporated. Glycosylases play a crucial role in BER, as they initiate the pathway by recognizing and excising specifically modified bases. If these enzymes are lacking or fail to function properly, the DNA damage can be camouflaged to the p53 protein, thus preventing the apoptosis process from being triggered and prolonging the cell lifespan (Lakin and Jackson, 1999; Meek, 2004; Culmsee and Mattson, 2005). Since HCV can infect B lymphocytes and prolong their lifespan, the present study investigates the activity of selected BER proteins (UDG, APEX1, and polymerases) in NE and ME of PBMC.
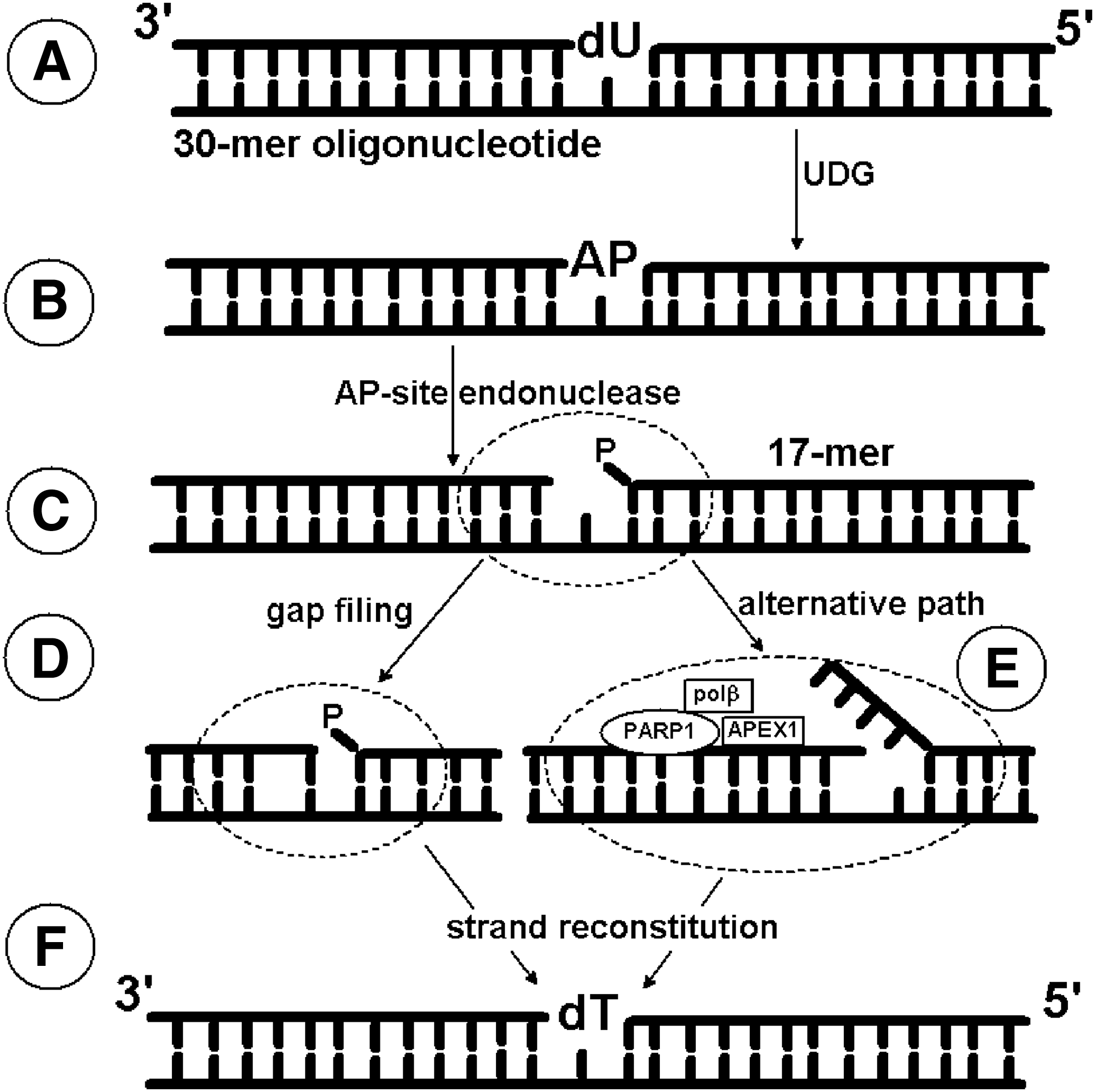
In our study, we used a modeled 30-mer double-strand oligonucleotide containing 2′-deoxyuridine (dU) as a DNA lesion. dU can appear in the genome due to cytidine deaminase activity, spontaneous 2′-deoxycytidine deamination, misincorporation instead of thymidine during replication, leading to an A:U mismatch formation. Consequently, the presence of dU as a DNA lesion is highly mutagenic and can be lethal to a cell. All organisms possess the following enzymes: uracil-DNA glycosylase removing dU from the genome and dUTP pyrophosphatase hydrolyzing 2′-deoxyuridine-5′-triphosphate (the substrate for polymerase) (Ingraham et al., 1986; Impellizzeri et al., 1991; Vassylyev and Morikawa, 1996; Vasilenko and Nevinsky, 2003). Therefore, we examined the effectiveness of the glycosylytic activity in NE and ME. In both cases, we found it to be lower in the patients before therapy than in the patients after therapy or the healthy volunteers (Figs. 2A, D and 3A, C). It is important to emphasize that UDG activity was only shown indirectly on the radiograms, since it produces only an AP site and not a single-strand break (ssb). The appearance of a shorter ssDNA fragment (17-mers), a manifestation of ssb in dsDNA structure, is a result of the AP-site cleavage activity by other enzymes, such as a specific endonuclease (e.g., APEX1), bifunctional glycosylase (e.g., hOGG1), or a spontaneous rearrangement under favorable conditions (Loeb and Preston, 1986; Kim and Wilson, 2012).
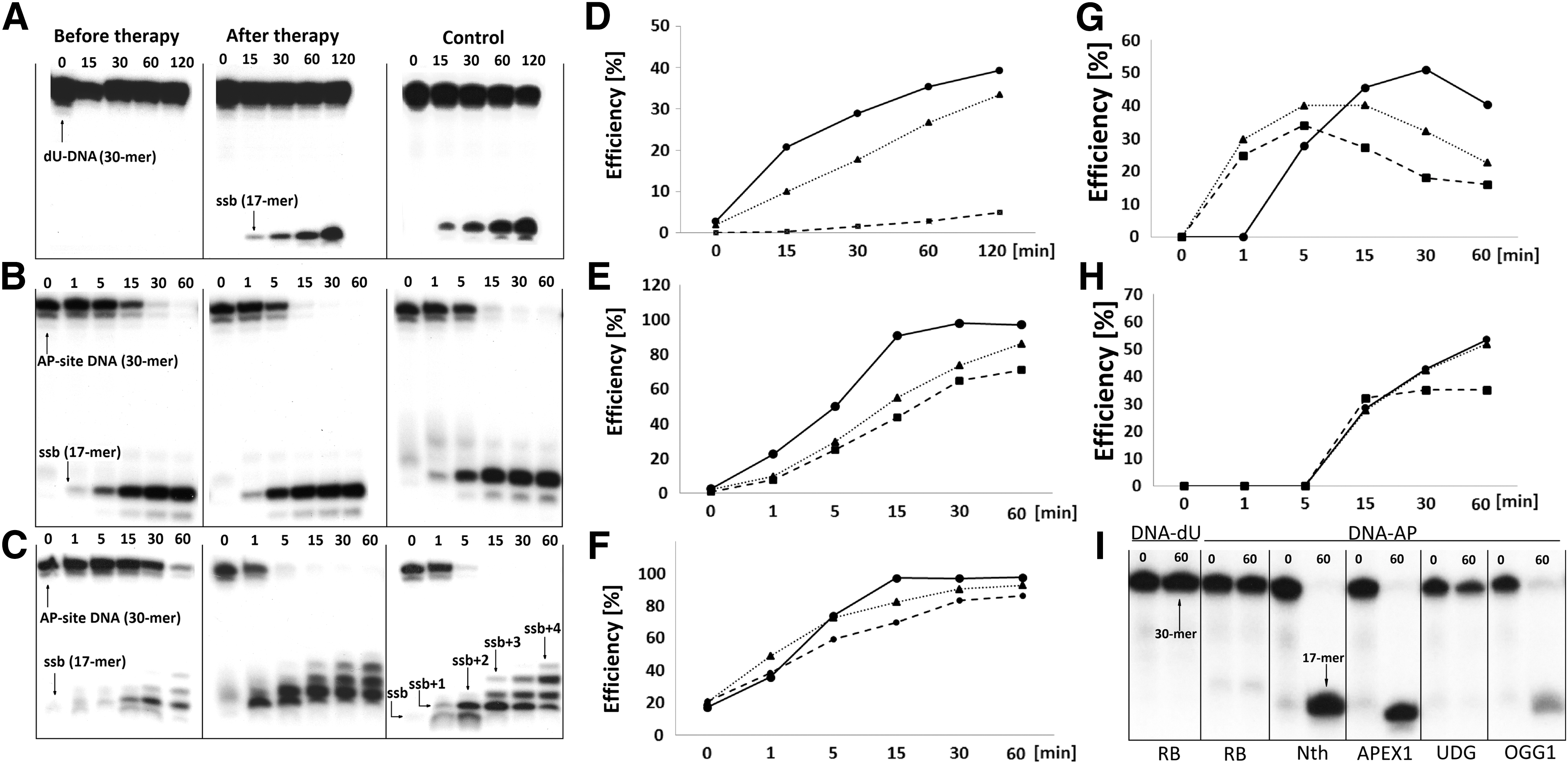
Representative denaturing polyacrylamide gels and corresponding time scale graphs showing the BER enzyme activity present in NE of PBMCs isolated from HCV-positive patients (before therapy --■--, after therapy ▪▪▲▪▪) and HCV-negative as control —●—.
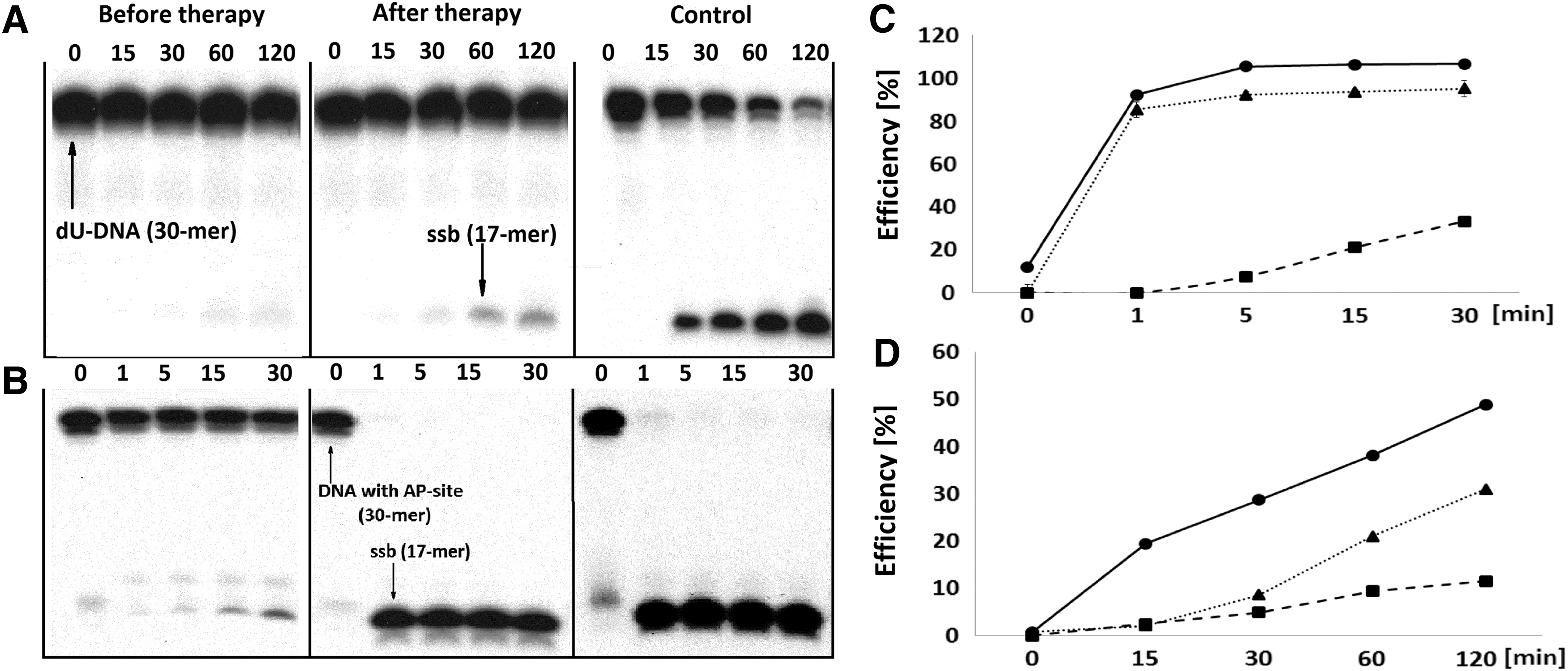
Representative denaturing polyacrylamide gels and corresponding time scale graphs showing the activity of the BER enzymes present in the ME of PBMCs isolated from patients: HCV positive (before therapy --■--, after therapy ▪▪▲▪▪) and HCV negative as control —●—.
To clarify this, the next stage examined the AP-site excision activity of the NE and ME. For NE, this activity was nearly the same for all studied groups (Fig. 2B, E), whereas in case of ME effectiveness in patients before therapy was significantly lower than in the patients after therapy or control group (Fig. 3B, D). These results indicate that proteins with AP-site lysis activity are more abundant in NE; suggesting that AP-site digestion is not significantly affected by HCV. If this were not the case, specific glycosylases would recognize the modified nucleobases and incise them, and the abundance of the enzyme could convert the formed AP-site to a ssb, providing an open frame for polymerase action. Both BER subpathways were found in the nucleus and the mitochondrion (Prakash and Doublié, 2015) and the schematic representation of their basic steps are summarized in Figure 1. Differences concerning the activity of polymerases were noted: polβ, polɛ, and polδ are present in the nucleus, whereas only polγ exists in the mitochondrion.
We also made an estimate of NE and ME polymerase activity, but it was very low and not measurable (data not shown). Accordingly, polβ was added to the extracts to force the reaction. It must be mentioned that polβ is a bifunctional enzyme with additional phosphatase activity (Sukhanova et al., 2005; Beard et al., 2006). In BER, polβ inserts only the first nucleotide in the reconstituted strand in both SP and LP, or releases an unhydrolyzed 5′-end sugar–phosphate moiety, leaving a 5′-end phosphate group that is acceptable for ligases. In all cases, the addition of more than one nucleotide was observed, but the reaction was most efficient in the control group and the patients after therapy (Fig. 2C). However, the first nucleotide was added in a greater effectiveness in the controls than in rest of the groups, but the curve profile adopted a similar shape in all investigated cases (Fig. 2G). Additionally, although in the presence of polβ and dNTP order of the ssb formation, efficiency retained the same as in the previous experiment lacking those components, the effectiveness was higher than before (Fig. 2F).
These results correlate well with the elongation of the ssb oligonucleotide (17-mers) by external polβ. Due to the fact that maximum efficiency was achieved after the first nucleotide addition (Fig. 2C, ssb +1) the yield decreases with subsequent additional bands, which appear on the radiogram of up to four units (Fig. 2C). We found that in the extracts isolated from patients before therapy, polymerase was active only for the first 15 min of incubation, whereas in patients after therapy and control group, this activity was observed throughout incubation (Fig. 2H). The difference in NE activity with the support of polβ indicates that the level of proteins interacting with polβ (e.g., PARP1, XRCC1, PCNA, RFC) was absent, or significantly lower in the NE derived from PBMC of HCV-positive patients than the posttherapy patients or the control group (Khodyreva et al., 2010).
The impairment of BER in lymphocytes after the infection with the HCV could be explained by integration of the genetic material of the virus to the part of the lymphocyte genome which is responsible for uracil-DNA glycosylase and AP-site endonuclease (e.g., APEX1), thus disrupting the activity of those enzymes (Figs. 2D, E and 3C, D). Moreover, we hypothesize that HCV decreases the level of proteins interacting with polymerases (e.g., PCNA, PARP1, XRCC1), causing interference with polβ, ɛ, δ, and γ or inhibiting their activity. As a consequence, the lifespan of HCV-infected lymphocytes is prolonged. As a matter of fact a lack of specific glycosylases or their low level can interfere with the proteins (e.g., p53) responsible for triggering the apoptosis process because of the failure to recognize DNA damage. As a result, HCV can exist in the lymphocyte reservoir for a long time. This could be, at least partly, the reason that classic therapy has been unsuccessful. The use of novel therapy results in regeneration of activity of UDG, endonuclease, and other crucial BER proteins such as PARP1 or PCNA. Therefore, it can be assumed that those medicines interact not only with hepatocytes, but also with PBMC. The alternatively proposed concomitant use of antiretroviral therapy can prohibit HCV infection of hepatocytes by lymphocytes.
We are fully aware of our work's limitation, particularly a small number of the patients participating in the study. There are several reasons for that. First and foremost, the OBV/PTV/r ± DSV ± RBV antiviral therapy is currently in clinical trials, thus the group of patients is selected and limited. Moreover, the cost of the therapy is very high, especially if we consider Polish conditions.
Conclusions
Nevertheless, based on the experimental findings presented above, we can conclude that the infection of lymphocytes by the HCV can significantly impair the activity of the DNA BER pathway in those cells. Moreover, use of the OBV/PTV/r ± DSV ± RBV antiviral therapy restore this DNA repair pathway, which could indicate that this therapy not only affects hepatocytes, but also eliminates the viruses' reservoir in lymphocytes. However, future studies are necessary.
Footnotes
Acknowledgments
This work was supported by grant UMO-2012/05/B/NZ5/01852 from the National Science Center and by the Medical University of Lodz, Poland, Research Program No. 503/3-045-02/503-31-002.
Disclosure Statement
No competing financial interests exist.