
Abstract
Long noncoding RNAs (lncRNAs) have been known to be involved in multiple diverse diseases, including osteoarthritis (OA). The present study aims at exploring the biological role of lncRNA plasmacytoma variant translocation 1 (PVT1) in OA and the underlying mechanism. Results showed that the expression of PVT1 was upregulated in OA chondrocytes compared with normal chondrocytes, silencing PVT1 inhibited the apoptosis of OA chondrocytes, and overexpression of PVT1 promoted the apoptosis of normal chondrocytes. To further investigate the underlying mechanism, miR-488-3p was predicted to be a targeted microRNA of PVT1. Different methods, including MS2 RNA immunoprecipitation (RIP), luciferase activity, and anti-AGO2 RIP, were performed to detect the interaction between PVT1 and miR-488-3p, which suggested that PVT1 negatively regulated miR-488-3p in OA chondrocytes. Moreover, PVT1 promoted the apoptosis of OA and normal chondrocytes through miR-488-3p. Collectively, this study revealed that lncRNA PVT1 regulated the apoptosis of chondrocytes by acting as a sponge for miR-488-3p in OA. PVT1 may be considered a new therapeutic target for the treatment of OA.
Intrduction
O
A growing body of evidence indicates noncoding RNAs (ncRNAs) as possible modulators of cell apoptosis (Su et al., 2016). MicroRNAs are small ncRNAs with a length of 20–25 nt that take part in the pathogenesis of OA. These ncRNAs are greater than 200 nucleotides in length with limited or no protein-coding capacity, and they are known as long noncoding RNAs (lncRNAs) (Frank et al., 2016). LncRNAs have been known to be involved in multiple diverse diseases, including OA (Song et al., 2014). LncRNA plasmacytoma variant translocation 1 (PVT1), located at 8q24.21, is associated with the cell proliferation, apoptosis, lymph node invasion, and metastasis (Cui et al., 2016). LncRNA PVT1 was originally identified as a common retroviral integration site in murine leukemia virus (MLV)-induced T lymphomas. Recently, PVT1 has been identified as a candidate oncogene. Increased copy number and overexpression of PVT1 have been found in many types of human cancers, including ovarian cancer, breast cancer, hepatocellular carcinoma, bladder cancer, and gastric cancer (Colombo et al., 2015). It has been reported that PVT1 was upregulated in osteosarcoma and promotes osteosarcoma development by acting as a molecular sponge to regulate miR-195 (Zhou et al., 2016). However, the role of PVT1 in OA remains unknown. A recent study found that PVT1 expressed nearly three-fold differently in OA than in normal tissues (Xing et al., 2014). Accordingly, we predicted that PVT1 may play a key role in the development of OA.
The present study, therefore, was aimed at exploring the biological role of lncRNA PVT1 in OA and the underlying mechanism. We determined the expression of PVT1 in OA chondrocytes, and we investigated the role of PVT1 in OA chondrocyte apoptosis. It has been reported that lncRNAs act as sponges for microRNAs during OA pathogenesis, and multiple microRNAs play a key role in OA (Tay et al., 2014). For example, miR-488 plays a positive role in chondrocyte differentiation/cartilage development by targeting ZIP-8 (Song et al., 2013). Therefore, we further analyzed the interaction between PVT1 and microRNA in the regulation of OA.
Materials and Methods
Human articular cartilage tissues
The 25 OA articular cartilages in this study were obtained from patients undergoing total knee arthroplasty. OA was confirmed by using radiographic images and diagnosed according to the American College of Rheumatology criteria. The 25 normal articular cartilages were isolated from patients with femoral neck fracture without OA or rheumatic arthritis who underwent total hip replacement surgery. Informed consent was obtained from all donors. The study was approved by the Human Ethics Committee of the People's Hospital of Hunan Province.
Chondrocyte cultures
The chondrocytes were isolated from articular cartilage samples as previously described (Yuan et al., 2016). Chondrocytes were incubated at 37°C under 5% CO2 in Dulbecco's modified Eagle's medium (DMEM) containing 10% fetal bovine serum.
Quantitative RT-PCR
Total RNAs were isolated by Trizol reagent (Invitrogen) and reverse transcribed. Quantitative analysis of the change in expression was performed by using SYBRs Premix Ex Taq™ (DRR42OA; TaKaRa) by the ABI 7300 system. GAPDH served as an internal standard. LncRNA PVT1 primers were as follows: 5′-TTGGCACATACAGCCATCAT-3′ (sense) and 5′-GCAGTAAAAGGGGAACACCA-3′ (antisense).
Cell transfection
The coding sequence of PVT1 was amplified and subcloned into the pcDNA3.1 (+) vector (Invitrogen). Chondrocytes were then transfected with a negative control vector or the pcDNA3.1 PVT1 plasmid by using Lipofectamine 2000 (Invitrogen). In addition, the siRNA against PVT1 were purchased from Riobo Company and transfected into chondrocytes by Lipofectamine 2000 according to the manufacturer's instructions. At 48 h post-transfection, cells were harvested for quantitative real-time PCR.
Cell viability and apoptosis assay
Cell viability was detected by using the cellTiter 96® AQueous one solution cell proliferation assay kit (Promega) according to the guidelines. Apoptosis was evaluated by using Hoechst Staining Kit (Beyotime) according to the manufacturer's instructions.
Western blot analysis
Proteins were resolved on 100 g/L SDS-polyacrylamide gels and electroblotted onto nitrocellulose membranes. Membranes were blocked with milk solution, followed by incubation with primary antibodies overnight at 4°C. The blots were then incubated with horseradish peroxidase-coupled goat anti-mouse or goat anti-rabbit secondary antibody. Primary antibodies are shown as follows: Bcl-2 (ab692, 1:5000; Abcam), Bax (ab32503, 1:5000; Abcam), MMP-1 (ab38929, 1:5000; Abcam), MMP-13 (ab39012, 1:5000; Abcam), and GAPDH (ab8245, 1:5000; Abcam).
RNA immunoprecipitation
OA chondrocytes were cotransfected with pcDNA-MS2, pcDNA-PVT1-WT-MS2, or pcDNA-PVT1-MUT-MS2 (GeneChem). After 48 h, cells were subjected to perform RNA immunoprecipitation (RIP) assays by using a GFP antibody (Abcam) and the EZ-Magna RIP™ RNA-Binding Protein Immunoprecipitation Kit (Millipore) according to the manufacturer's protocol.
For anti-AGO2 RIP, cells were transfected with miR-488-3p mimics or miR-NC. After 48 h, cells were subjected to perform RIP assays by using an AGO2 antibody (Abcam).
Luciferase reporter assay
PVT1-WT or PVT1-MUT reporter plasmid was co-transfected with miR-488-3p mimic or miR-NC into OA chondrocytes by using Lipofectamine 3000. Luciferase activity was measured by using a dual luciferase reporter assay system (Riobo).
Statistical analysis
Data were presented as mean ± standard deviation. Statistical analysis was performed by using SPSS software version 19.0. Comparisons of continuous data were analyzed by using the Student's t-test between the two groups. p < 0.05 was considered statistically significant.
Results
The expression of PVT1 is upregulated in OA chondrocytes
To explore whether PVT1 is altered in OA, the expression of PVT1 was measured by quantitative RT-PCR in 25 OA patients and 25 normal patients. As shown in Figure 1, the PVT1 expression was significantly upregulated in the OA group compared with the normal group.

The expression of PVT1 in OA chondrocytes. PVT1 expression of chondrocytes isolated from the articular cartilage of normal objects and OA patients, n = 25. Data are presented as mean ± SD. **p < 0.05.
Silencing of PVT1 inhibits the apoptosis of OA chondrocytes
Chondrocyte apoptosis is a key player that is responsible for cartilage degradation in OA patients. To determine the role of PVT1 in OA chondrocyte apoptosis, we explored the effect of silencing PVT1 on OA chondrocyte apoptosis. Three siRNAs were designed to silence PVT1 (Fig. 2A), and the si-PVT1#2 showed the best silencing effect, which was used for the following studies. Moreover, silencing of PVT1 could increase the cell viability and decrease the apoptosis cell number (Fig. 2B, C). Meanwhile, the upregulation of Bcl-2 and downregulation of Bax, cleaved caspase-3 also indicated that the silencing of PVT1 could inhibit the apoptosis of OA chondrocytes (Fig. 2D). Matrix metalloproteinases (MMPs) plays a critical role in keeping the balance between degradation and synthesis of extracellular matrix, especially MMP-1 and MMP-13. The upregulation of MMP-1 and MMP-13 is an important pathological marker of OA. Therefore, we further investigated the effect of silencing PVT1 on the expression of MMP-1 and MMP-13 in OA. As shown in Figure 2E, the silencing of PVT1 downregulated the expression of MMP-1 and MMP-13. These results indicated the pro-apoptotic effect of PVT1 in OA chondrocytes.
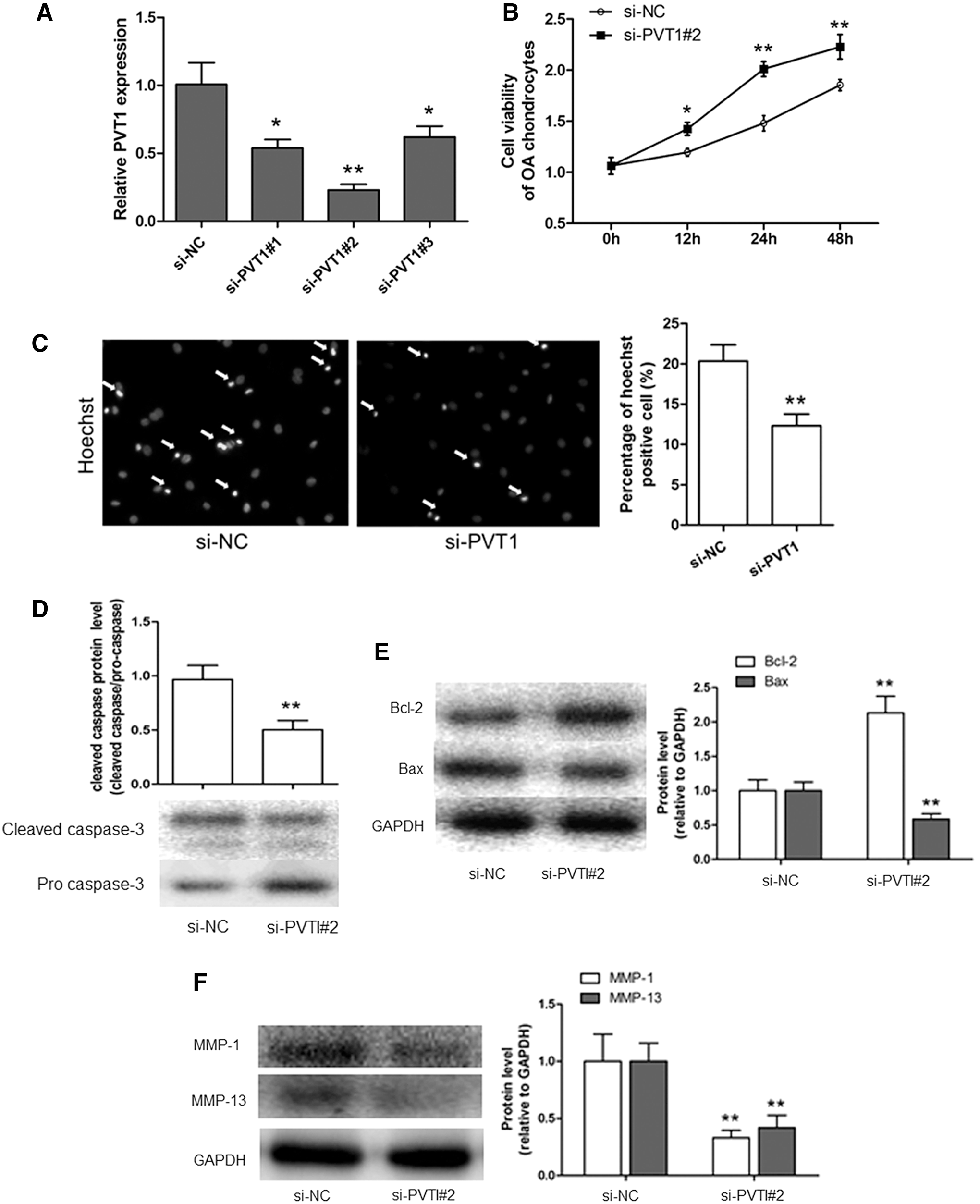
The effect of silencing PVT1 on OA chondrocyte apoptosis.
Overexpression of PVT1 promotes the apoptosis of normal chondrocytes
To further confirm the pro-apoptotic effect of PVT1 in OA, and considering the important role of chondrocyte apoptosis in OA, we investigated the effect of PVT1 overexpression on the apoptosis of normal chondrocytes. As shown in Figure 3A, pcDNA3.1 PVT1 was used to overexpress the PVT1, which could decrease the chondrocyte viability and increase the apoptosis cell number (Fig. 3B, C). Meanwhile, the downregulation of Bcl-2 and upregulation of Bax, cleaved caspase-3 also indicated that the overexpression of PVT1 could promote the apoptosis of normal chondrocytes (Fig. 3D, E). Moreover, PVT1 overexpression upregulated the expression of MMP-1 and MMP-13 (Fig. 3F). These results suggested that PVT1 played an important role in the apoptosis of normal chondrocytes during OA.
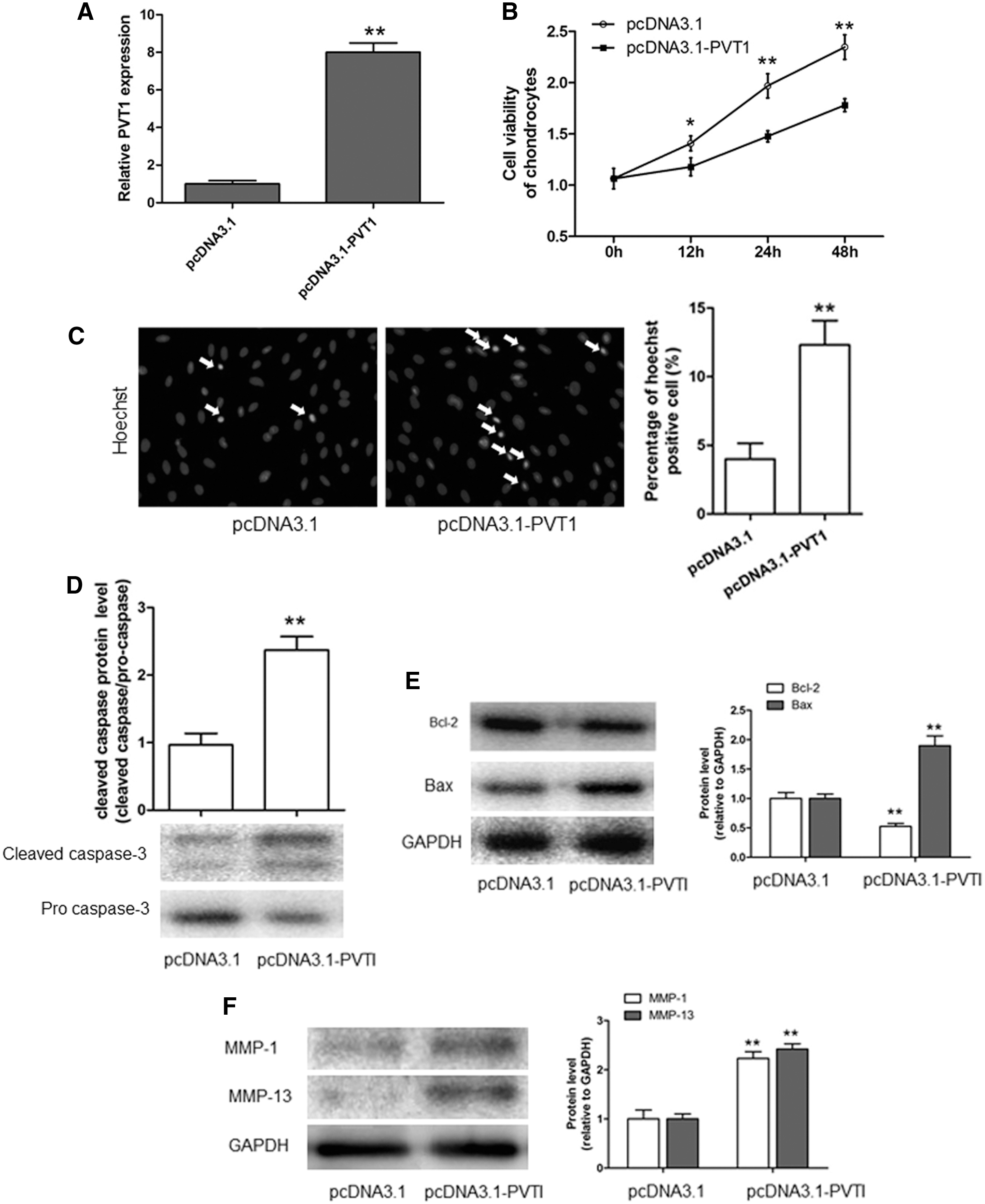
The effect of PVT1 overexpression on normal chondrocyte apoptosis.
PVT1 interacts with miR-488-3p in OA chondrocytes
A series of results mentioned earlier have indicated the pro-apoptotic effect of PVT1 in OA chondrocytes; however, the underlying mechanism remains unknown. Recent studies suggest a complex signaling network between lncRNAs and miRNAs in a variety of biological and pathological responses. We reason that a similar link between PVT1 and miRNA is likely to exist in OA. To investigate the interaction between PVT1 and miRNAs, a total of 23 miRNAs were predicted as a target of PVT1 by Starbase 2.0. OA chondrocytes were transfected with si-NC and si-PVT1 to detect the differential expression of 23 miRNAs, and it was found that the expression of miR-488-3p showed the most obvious change (Fig. 4A). Moreover, we measured the expression of miR-488-3p in OA, and we found that the miR-488-3p expression of chondrocytes was significantly decreased in the OA group compared with the normal group. These results suggested that PVT1 was involved in regulating miR-488-3p during OA (Fig. 4B). To further investigate the mechanism of interaction between PVT1 and miR-488-3p, the following studies were conducted. The miR-488-3p binding site of PVT1 was also predicted, as shown in Supplementary Figure S1 (Supplementary Data are available online at
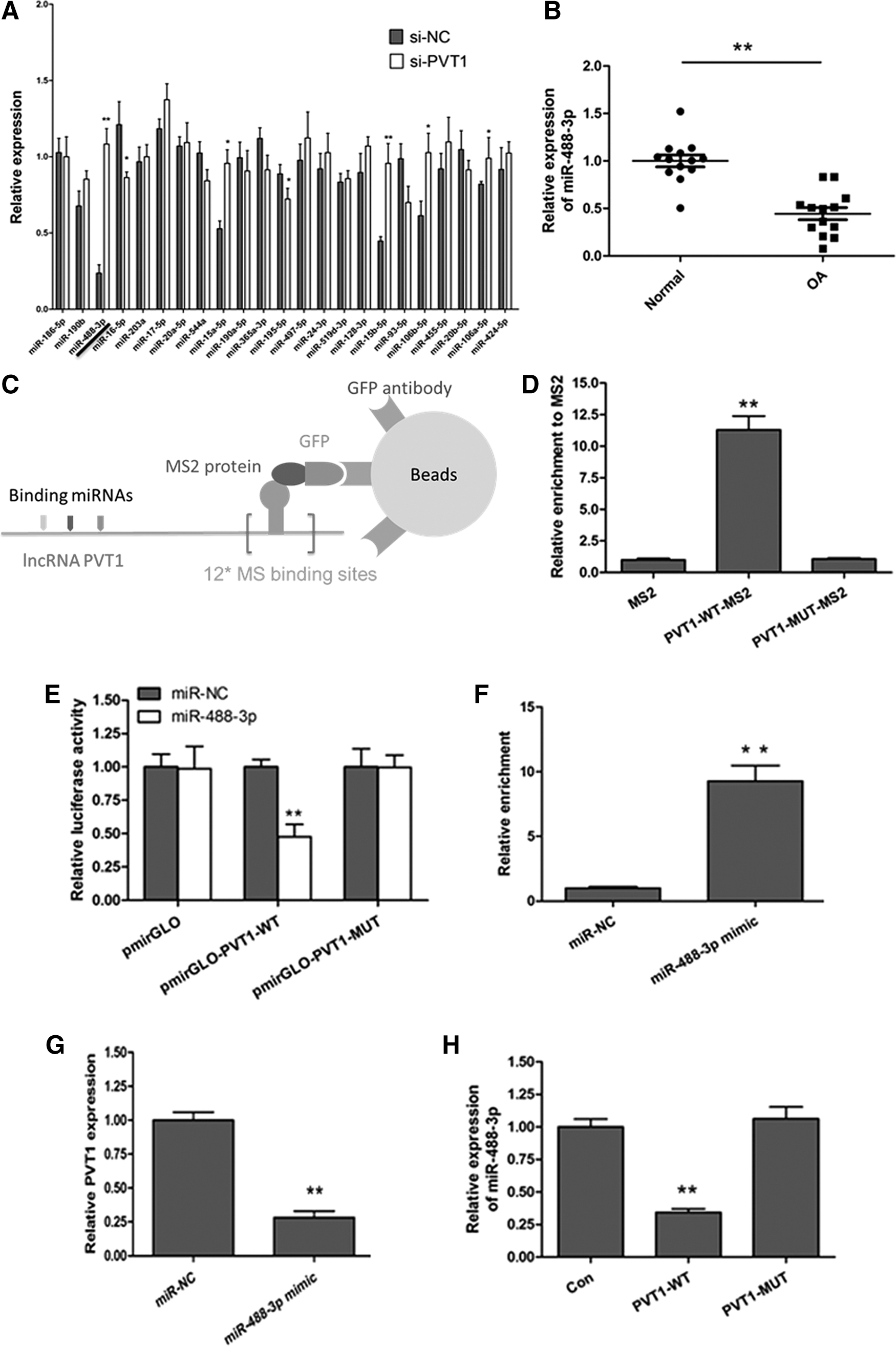
PVT1 interacts with miR-488-3p in OA chondrocytes.
It has been known that miRNAs bind the mRNAs in an AGO2-dependent manner. To determine whether PVT1 interacts with miR-488-3p through AGO2, we, therefore, conducted anti-AGO2 RIP in chondrocytes that were transiently overexpressed with miR-488-3p. The result showed that PVT1 pull-down by AGO2 was significantly enriched in miR-488-3p overexpression of chondrocytes (Fig. 4F). Finally, we found that overexpression of miR-488-3p could decrease PVT1 expression (Fig. 4G), and overexpression of PVT1 suppressed miR-488-3p expression (Fig. 4H). All of these results suggested that PVT1 acted as a sponge of miR-488-3p in OA chondrocytes.
PVT1 promotes cell apoptosis through miR-488-3p in OA chondrocytes and normal chondrocytes
In view of PVT1 playing an important role in the apoptosis of chondrocytes during OA and PVT1 acting as a sponge of miR-488-3p in OA chondrocytes, we speculate that miR-488-3p is involved in PVT1 and promotes chondrocyte apoptosis during OA.
To investigate whether miR-488-3p is involved in the regulation of PVT1 in OA chondrocyte apoptosis, we transfected the miR-488-3p inhibitor into PVT1-silencing chondrocytes and detected the effect on apoptosis. As shown in Figure 5A, the miR-488-3p inhibitor could decrease the upregulating effect of si-PVT1 on miR-488-3p. The miR-488-3p inhibitor also abolished the increase of the cell viability and decrease of the apoptosis cell number induced by si-PVT1 (Fig. 5B, C). Meanwhile, si-PVT1 induced the upregulation of Bcl-2 and downregulation of Bax, cleaved caspase, MMP-1, and MMP-13, which were reversed by the miR-488-3p inhibitor (Fig. 5D–F). Song et al. have reported that miR-488-3p inhibitor treatment only would promote chondrocyte differentiation/cartilage development (Song et al., 2013). These results revealed that PVT1 promoted OA chondrocyte apoptosis through miR-488-3p.
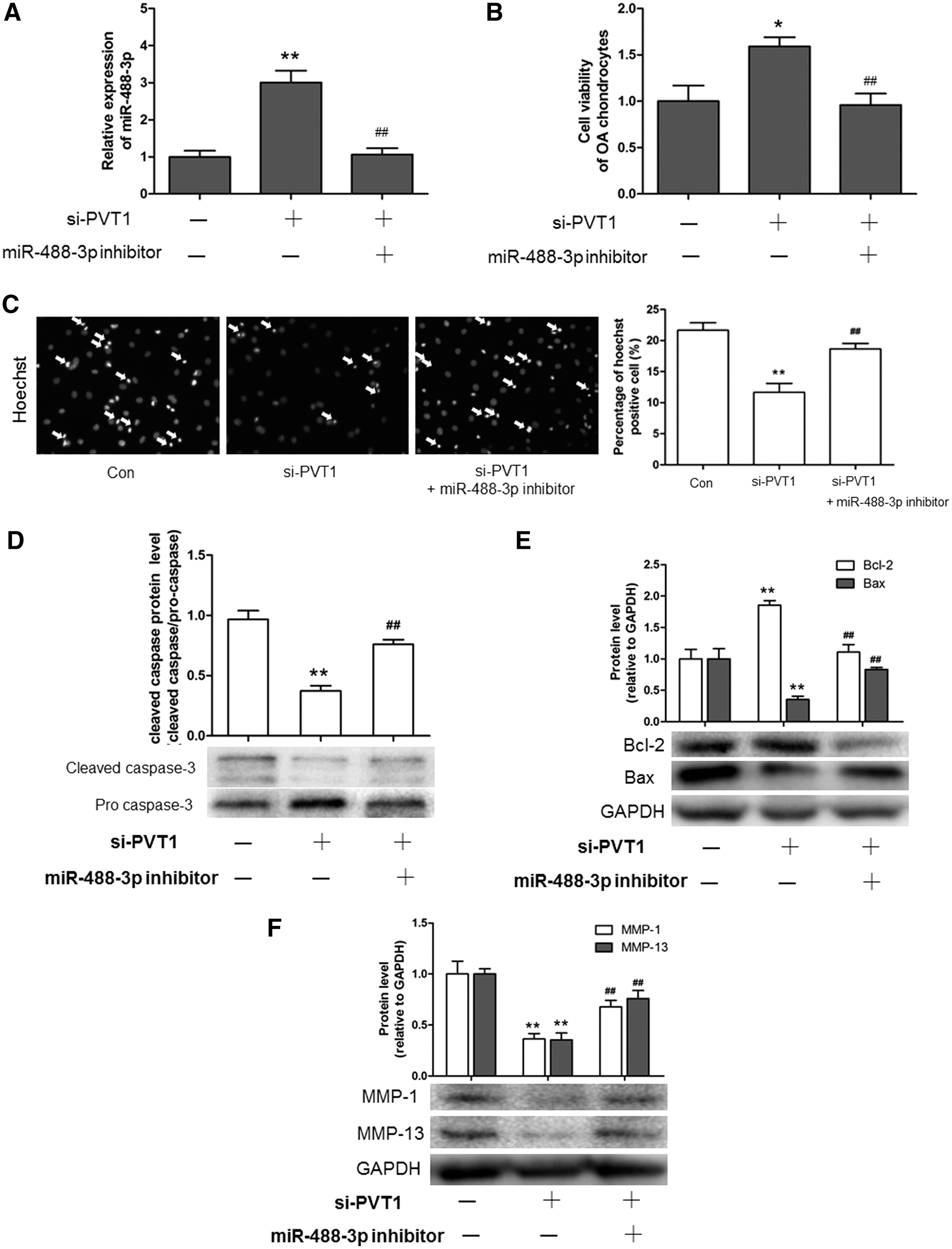
PVT1 promotes OA chondrocyte apoptosis through miR-488-3p.
Moreover, to investigate whether miR-488-3p was involved in the regulation of PVT1 in normal chondrocyte apoptosis, we transfected the miR-488-3p mimic into PVT1 overexpression of normal chondrocytes and detected the effect on apoptosis. As shown in Figure 6A, the miR-488-3p mimic could increase the downregulating effect of pcDNA3.1 PVT1 on miR-488-3p. The miR-488-3p mimic also abolished the decrease of the cell viability and the increase of the apoptosi cells number induced by pcDNA3.1 PVT1 (Fig. 6B, C). Meanwhile, pcDNA3.1 PVT1 induced the downregulation of Bcl-2 and the upregulation of Bax, cleaved caspase, MMP-1, and MMP-13, which were reversed by the miR-488-3p mimic (Fig. 6D–F). These results suggested that miR-488-3p was involved in PVT1 and promoted chondrocyte apoptosis during OA.
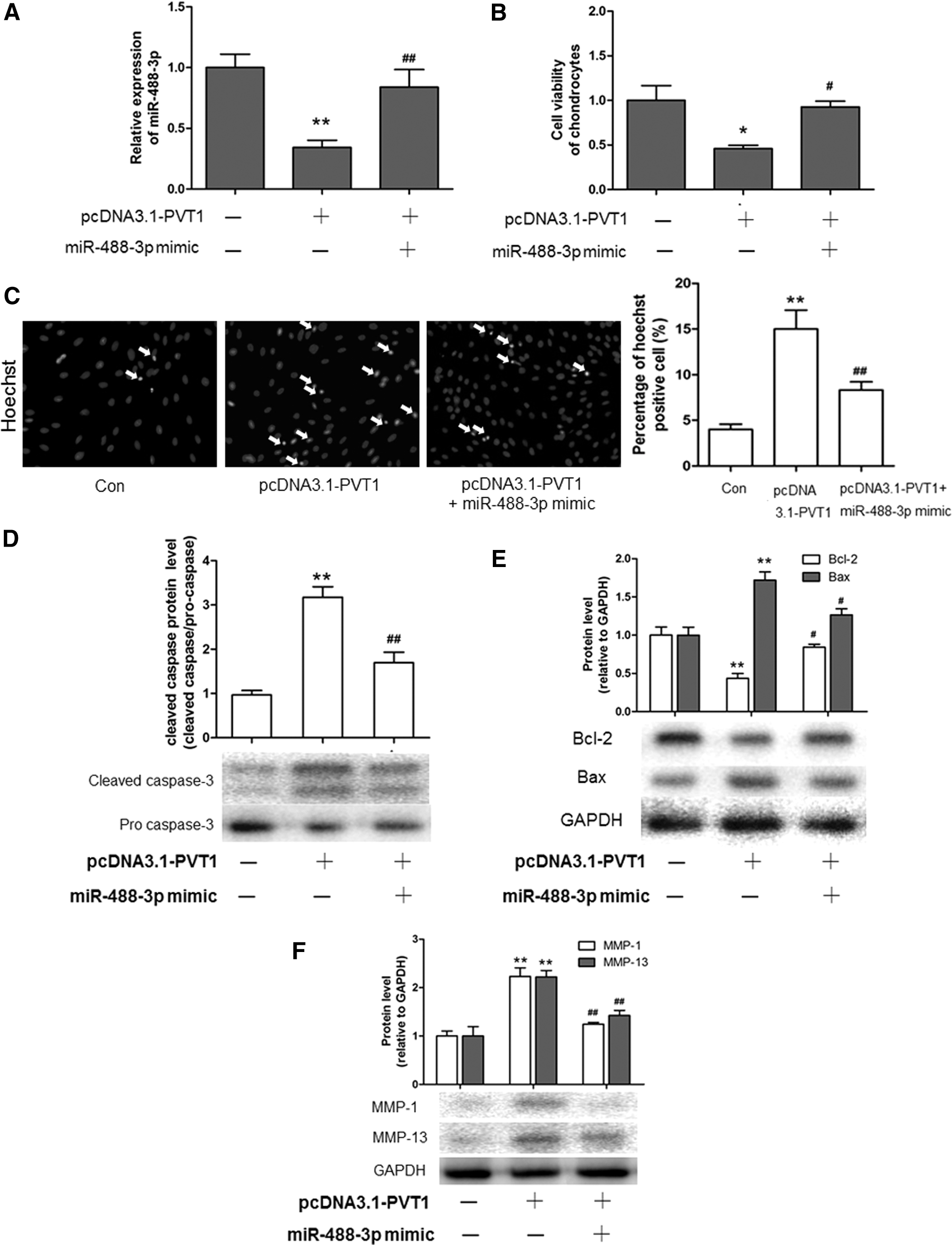
Overexpression of PVT1 promotes normal chondrocyte apoptosis through miR-488-3p.
Discussion
The scope of RNA's biological functions has been recently expanded from a single role of a messenger for protein synthesis to a wide spectrum of regulatory and structural roles in protecting chromosome integrity, maintaining genomic architecture, X-chromosome inactivation, imprinting, transcription, translation, and epigenetic regulation (Khorkova et al., 2015). The newly discovered RNA functions are mostly carried out by noncoding RNA (ncRNA), whereas miRNAs have been extensively studied. LncRNAs still constitute a new, potentially fascinating, territory that is yet to be explored (Perry and Ulitsky, 2016).
LncRNA PVT1 is encoded by the human PVT1 gene (also known as the Pvt1, oncogene). PVT1 was first discovered in 1984 as an activator of MYC in murine plasmacytoma variant translocations, which plays an oncogenic role in various types of human cancer (Colombo et al., 2015). For example, lncRNA PVT1 accelerated tumorigenesis of nonsmall cell lung cancer (Yang et al., 2014) and participated in tumor progression of hepatocellular carcinoma patients (Ding et al., 2015). Moreover, PVT-1 resulted in a poor prognosis by inhibiting apoptosis of colorectal cancers (Takahashi et al., 2014); PVT1 promoted proliferation and was associated with a poor prognosis in gastric cancer (Kong et al., 2015). Furthermore, PVT1 was associated with a poor prognosis in patients with pancreatic cancer (Xie et al., 2016). Recently, Zhou et al. (2016) found that PVT1 expression was higher in osteosarcoma tissues, and they demonstrated that PVT1 was involved in regulating proliferation, migration, invasion, and apoptosis of osteosarcoma cells. In the present study, the expression of PVT1 was upregulated in OA chondrocytes. Moreover, we found that silencing of PVT1 expression by siRNA inhibited the apoptosis of OA chondrocytes, and overexpression of PVT1 promoted the apoptosis of normal chondrocytes. These results indicate that PVT1 plays a key role in regulating the apoptosis of chondrocytes during OA.
An increasing number of studies support that lncRNAs may possess competing endogenous RNA (ceRNA) activity to act as sponges for microRNAs during OA pathogenesis (Tay et al., 2014). For example, PCGEM1 stimulates proliferation of osteoarthritic synoviocytes by acting as a sponge for miR-770 (Zhang et al., 2016), and UFC1 promotes proliferation of chondrocytes in OA by acting as a sponge for miR-34a (Kang et al., 2016). These findings trigger us to explore the functional association between lncRNA PVT1 and microRNA. In the present study, 23 miRNAs were predicted as targets of PVT1 by starbase 2.0, and it was found that the expression of miR-488-3p showed the most obvious change after silencing of PVT1. It has been reported that miR-488 was significantly decreased in OA chondrocytes, which plays a positive role in chondrocyte differentiation/cartilage development (Song et al., 2013). We then confirmed that PVT1 negatively regulated miR-488-3p in OA chondrocytes through different methods, including MS2 RIP, luciferase activity, and anti-AGO2 RIP. Following these results, we further found that miR-488-3p was involved in the pro-apoptotic effect of PVT1 in OA and normal chondrocytes.
The expression of miR-15a-5p and miR-15b-5p also showed significant upregulation on PVT1 silencing apart from miR-488-3p. It has been reported that hsa-miR-15a exerts protective effects against OA by targeting aggrecanase-2 (ADAMTS5) in human chondrocytes (Lu et al., 2016). Recently, Chen and Tian (2017) found that miR-15a-5p regulated viability and matrix degradation of human OA chondrocytes via targeting VEGFA. Therefore, PVT1 regulates OA chondrocyte apoptosis; further research needs to be conducted to verify whether this occurs through miR-15a-5p.
Conclusion
In summary, this study revealed that lncRNA PVT1 regulated the apoptosis of chondrocytes in OA by acting as a sponge for miR-488-3p. PVT1 may be considered a new therapeutic target for the treatment of OA. The opposite effects of PVT1 on apoptosis in cancer and OA may be due to different downstream signal pathways, which would serve as the emphasis in our further research.
Footnotes
Acknowledgment
This work was funded by The Scientific and Technological Funds of Hunan Province (2015JC3064).
Disclosure Statement
No competing financial interests exist.
References
Supplementary Material
Please find the following supplemental material available below.
For Open Access articles published under a Creative Commons License, all supplemental material carries the same license as the article it is associated with.
For non-Open Access articles published, all supplemental material carries a non-exclusive license, and permission requests for re-use of supplemental material or any part of supplemental material shall be sent directly to the copyright owner as specified in the copyright notice associated with the article.