
Abstract
Conotruncal heart defects (CTDs) are severe malformations of outflow tract with heterogeneous morphology. Several missense variants of CITED2 have been identified to cause CTDs in recent researches. In this study, we screened the coding regions of CITED2 in 605 Chinese children with CTDs and found two possible pathogenic mutant sites: p.Q117L and p.T257A, both located in the conserved regions of CITED2. Then, we investigated the biological and functional alterations of them. Western blotting showed low level of protein expression of mutant Q117 and T257A compared with wild-type CITED2. Dual-luciferase reporter assay demonstrated that mutant Q117 and T257A decreased the ability of CITED2 to modulate the expression of paired-like homeodomain transcription factor 2 gamma (PITX2C), which are closely related to cardiac growth and left–right patterning. Meanwhile, T257A also exhibited impaired ability to mediate vascular endothelial growth factor expression, another gene closely associated with the normal development of cardiovascular system. Three-dimensional molecular conformation showed reduced hydrogen bond between Asp254 and mutant Thr257, indicating the weakened stability and binding ability of CITED2. All these results suggest that CITED2 mutations in conserved regions lead to disease-causing biological and functional changes and may contribute to the occurrence of CTDs.
Introduction
C
CITED2, also known as CBP/P300-interacting transactivator with Glu/Asp-rich carboxy-terminal domain 2, is a CBP/P300-interacting protein, which is highly conserved among mammals and widely expressed in vertebrates (Chen et al., 2012). CITED2 is involved in developmental regulation of embryonic heart, neural tube, neural crest cells, adrenal glands, placenta, liver, lung, eye, and gonads by cooperating with other transcription factors (Lopes Floro et al., 2011). Lack of CITED2 during gestation is lethal, indicating the essential role of CITED2 in embryonic development. CITED2 has three regions, namely CR1, CR2, and CR3, which are conserved among CITED family members (Braganca et al., 2002). CR2 participates in all known important biological activities of CITED2 through its binding to P300-CH1 (Bhattacharya et al., 1999; Braganca et al., 2002), while the functions of CR1 and CR3 are poorly understood. CITED2 also has a unique Serine–Glycine-Rich Junction (SRJ, residues 161–199) (Andrews et al., 2000; Yin et al., 2002), which has been identified to be a mutational hotspot; variants located in this region have higher risk to cause congenital cardiovascular malformations.
As mentioned above, CITED2 lacks typical DNA binding sites and may act as a cofactor to regulate the expression of specific genes such as paired-like homeodomain transcription factor 2 gamma (PITX2C) and vascular endothelial growth factor (VEGF) (Bhattacharya et al., 1999; Bamforth et al., 2004); abnormal expression of both have been clearly identified to be associated with CTDs. In vivo, CITED2 regulates PITX2C through cooperation with TFAP2C. CITED2 knockout mice developed various kinds of laterality defects and conotruncal malformations such as right atrial isomerism, hyposplenia, double-outlet right ventricle, common arterial trunk, and IAA because of abnormal expression of PITX2C (Bamforth et al., 2004). CITED2 can also block HIF1A transcriptional activity by competitively inhibiting the interaction between HIF1A and CBP/P300. The expression of HIF1A-induced gene VEGF could be influenced if the competition between CITED2 and HIF1A for CBP/p300 is perturbed.
Since Sperling et al. (2005) first uncovered the connection between congenital heart diseases and CITED2 mutations in 2005, a growing number of mutations have been reported to cause CHDs. Most of them are located in the SRJ region (Sperling et al., 2005; Chen et al., 2012; Li et al., 2012; Xu et al., 2014). Our study focuses on CTDs, which is a main kind of congenital cardiovascular malformations; the mutations involved are both located in CITED2 conserved regions instead of SRJ. We seek to clarify the relationship between the CITED2 variants in CRs and CTDs by analyzing the biological and functional alterations of these mutants.
Materials and Methods
Ethics statement
The study conformed to the ethical guidelines of the 2013 Declaration of Helsinki and was approved by Local Ethics Committees of Xinhua Hospital (Shanghai, China).
Patient samples and DNA extraction
CTD patients' blood samples alone with detailed clinical diagnoses were obtained from Xinhua Hospital and Shanghai Children's Medical Center during May 2013 to March 2015. Phenotypes of these patients are listed in Table 1. Genomic DNA was extracted from the blood sample using TIANamp Genomic DNA Kit (TIANGEN, Beijing, China). Corresponding 300 control blood samples were also gained from the two hospitals and DNA was extracted using the same method.
Mutation screening and bioinformatic analysis
Exon regions of CITED2 were tested by target sequence technique for mutant screening. Then, all candidate mutations were confirmed by Sanger sequencing. The PolyPhen-2 and SIFT programs were used to predict the disease-causing potential of CITED2 mutations. Species protein sequences of CITED2 were obtained from NCBI (
Plasmid construction and site-directed mutagenesis
The CITED2 expression vector containing human CITED2 cDNA was purchased from OriGene (Rockville, MD). Online software PrimerX (
PITX2C promoter and VEGF promoter regions were amplified by PCR from human genomic DNA and then cloned into the luciferase reporter pGL3-basic vector at the sites of Kpn1 and Xho1.
The HIF1A cDNA expression vector and the TFAP2C cDNA expression vector are generous gifts from Prof. Hong Pan (National Research Institute for Family Planning, China).
All the above-mentioned plasmids were verified by DNA sequence, and the primers used for PCR are listed in Table 2.
Cell culture and transient transfection
C2C12 and 293T cells were maintained in DMEM high glucose medium (HyClone, Utah) with 10% fetal bovine serum (Invitrogen, California) and 100 U/mL penicillin/streptomycin in humidified atmosphere at 37°C with 5% CO2. For VEGF-Luc reporter assay, cells with transfected plasmids were maintained in hypoxia condition with 1% O2. Fugene HD transfection reagent (Promega, Madison, WI) was used for transfection according to the manufacturer's instruction.
Western blotting and cycloheximide chase assay
Total proteins of 293T cells were extracted using total protein lysis buffer RIPA (Beyotime, Shanghai, China), separated by 12.5% sodium dodecyl sulfate–polyacrylamide gel electrophoresis (SDS-PAGE), and transferred to nitrocellulose membranes using a Mini-PROTEAN Tetra System (Bio-Rad). Primary monoclonal antibody anti-CITED2 (1:2000; EPR3416(2); Abcam) and the secondary antibody anti-rabbit (1:10,000; 111035003; Jackson) were used to analyze the presence of CITED2 protein. β-actin was chosen as inner control. Detection of targeted protein bands was performed by ECL western blotting detection reagents according to the manufacturer's protocol. Gray value of every band was quantified using Image Lab software.
For cycloheximide chase assay, 293T cells were transfected with WT-CITED2 (wild type), Q117L, and T257A plasmids. Thirty-six hours after transfection, cells were treated with 100 μg/mL cycloheximide (MCE, HY-12320) for indicated time (0, 20, 40, 80 min). Western blotting was used to analyze the protein expression, and the results were exhibited as ratio of CITED2/β-actin gray value.
Quantitative real-time PCR
293T cells were maintained in 12-well plates. Total RNA was isolated using 1 mL TRIzol reagent (15596026; Invitrogen) according to the manufacturer's instructions 24 h after transfection. Total RNA (about 1 μg) was retrotranscribed with PrimeScript™ RT reagent (TaKaRa, Japan). qPCR was performed according to the SYBR® Premix Ex Taq™ II protocol (TaKaRa, Japan). GAPDH was used as control to normalize the expression levels. Primers used in qRT-PCR are listed in Table 2.
Dual-luciferase reporter assay
C2C12 cells were cotransfected with 50 ng WT-CITED2 or mutants, transcription factor TFAP2C or HIF1A, and 100 ng PITX2C or VEGF promoter reporter plasmid. Twenty nanograms of pRL-TK plasmid (E2241; Promega) were used as internal control. Firefly and Renilla luciferase activities were measured using the Dual-Glo Luciferase Assay System 6 Kit (Promega, Madison, WI) and the Centro XS3 LB 960 Microplate Luminometer (Berthold, Bad Wildbad, Germany). The results represent the means of three independent experiments performed in triplicate.
Three-dimensional reconstruction of CITED2 mutant protein
CITED2 C-terminal protein (216–259) three-dimensional (3D) model was obtained from SWISS-MODEL (
Statistical analysis
Statistical analysis was performed using t-test or one-way ANOVA. All data were analyzed by Prism GraphPad 6.02 software, and p < 0.05 was considered significant.
Results
Screening and bioinformatic analysis of CITED2 mutations
We sequenced the entire CITED2 open reading frame in 605 CTD cases and 300 unrelated control individuals. Several nonsynonymous variants were identified, among which two mutants—p.Q117L and p.T257A (Table 3)—had not yet been reported in CTD patients and were predicted to be pathogenic by SIFT and PolyPhen-2. The screening results were verified by Sanger sequence (Fig. 1b).
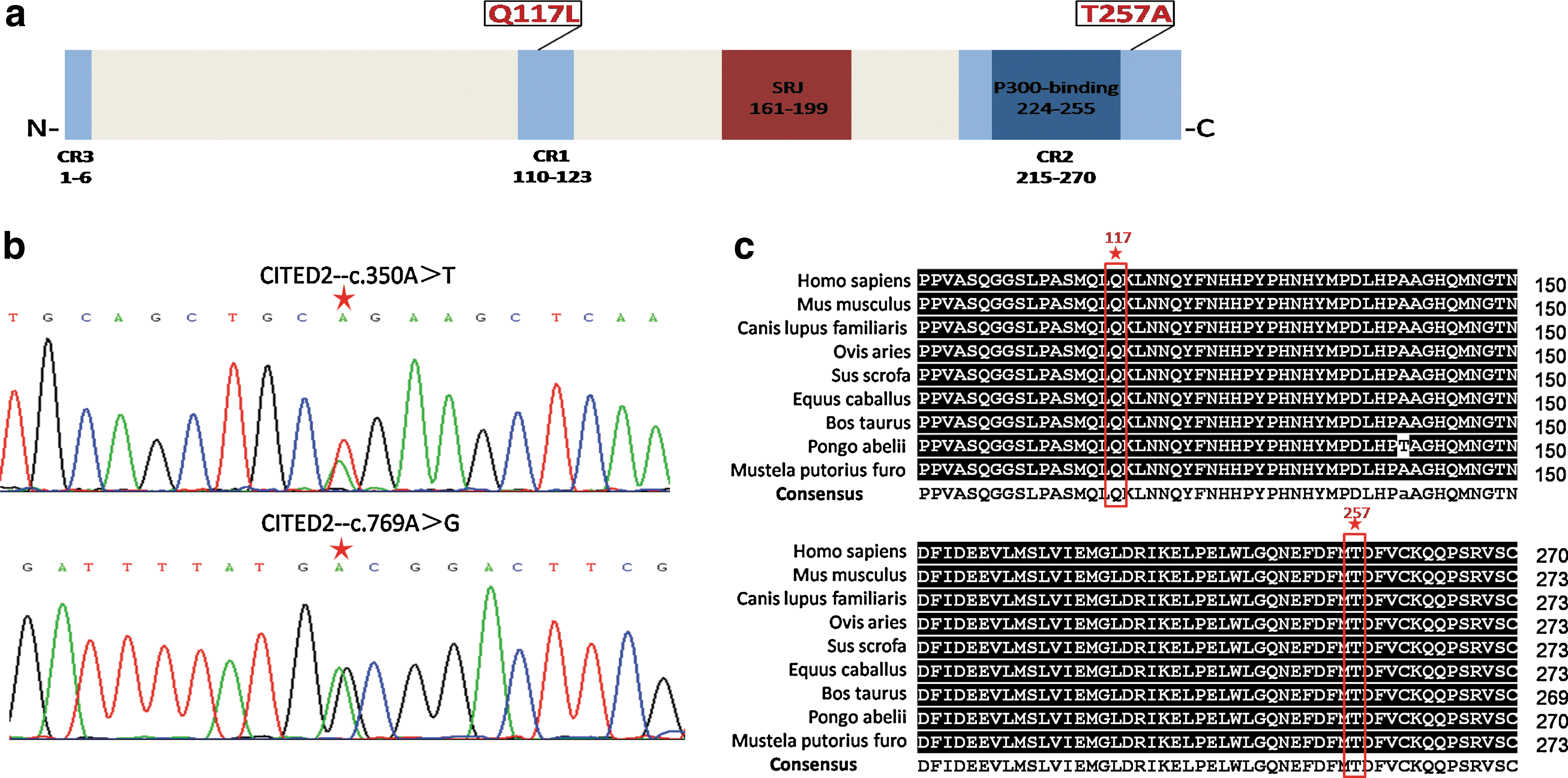
Biological analysis of CITED2 and mutants.
Mutant Q117L locates in CITED2 conserved region 1 (CR1); it was detected from a patient with TGA, patent ductus arteriosus, and atrial septal defect (ASD). T257A locates in another CITED conserved region—CR2 (Fig. 1a); the patient carrying T257A suffered from TOF, patent foramen ovale, and dextro-aortic arch. Homology analysis of CITED2 protein showed that two conserved region mutants are also highly conserved among species (Fig. 1c). These observations suggested that the two amino acids might be functionally important, and severe congenital cardiac defects could be associated with the p.Q117L and p.T275A variations.
CITED2 mutations decrease protein expression and have impact on transcriptional activities
To explore if the predicted disease-causing mutants could result in functional changes of CITED2, we first checked the protein expression level of WT-CITED2 and its mutants by western blot (Fig. 2a). Gray intensity ratio of CITED2/β-actin was analyzed using GraphPad software. The results showed that both Q117L and T257A had lower protein expression than WT-CITED2 (Fig. 2b).
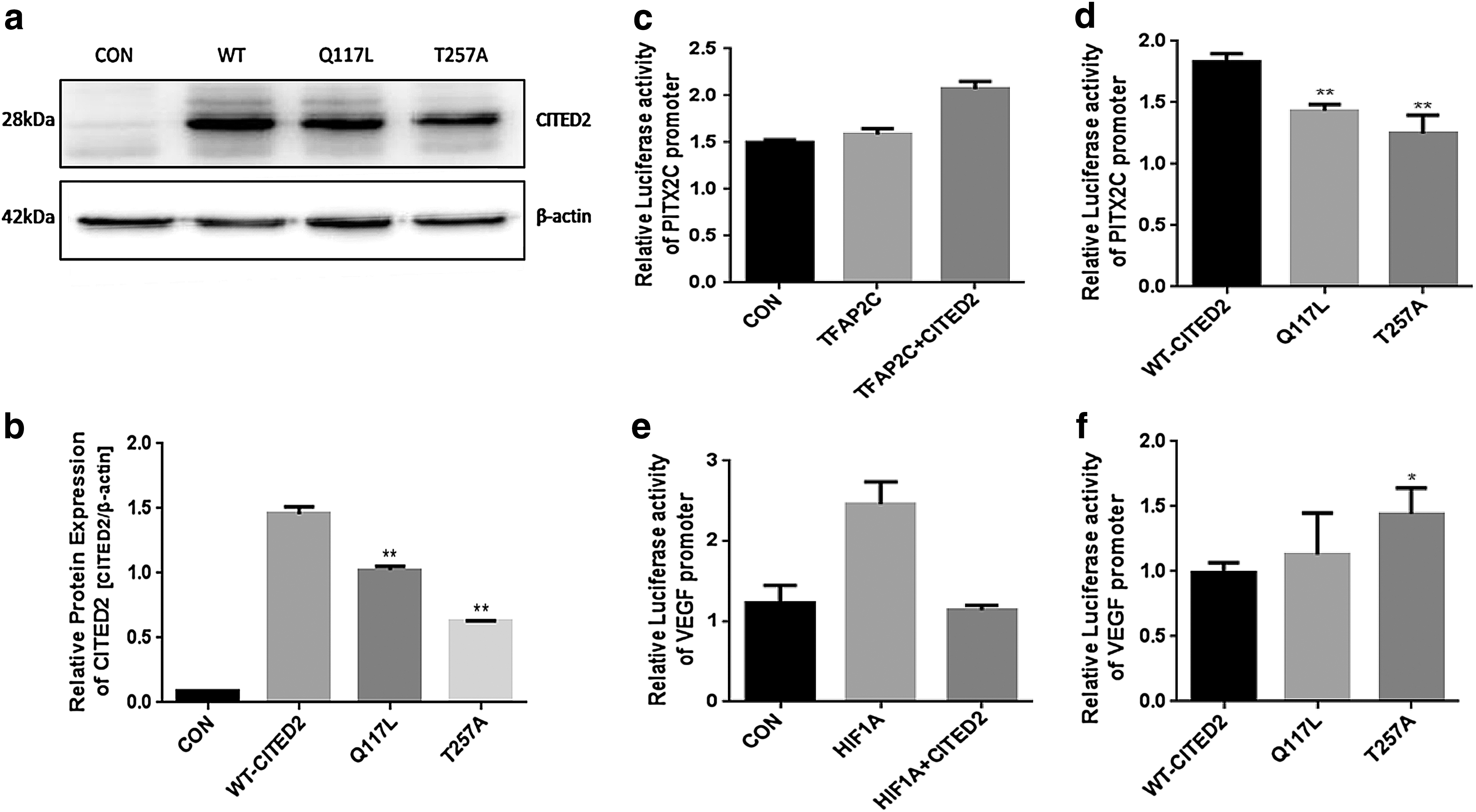
Functional analysis of CITED2 and mutants.
Dual-luciferase reporter assay was used to assess whether the mutations would have impact on transcriptional activities of CITED2. As shown in Figure 2c, TFAP2C alone exhibits a faint transcriptional activation in our assay and increased effect on targeted PITX2C promoter when cotransfected with WT-CITED2. Then, we cotransfected TFAP2C with CITED2 wild type and mutants. Both Q117L and T257A showed weaker synergistic effect, resulting in a lower expression of downstream PITX2C (Fig. 2d). Meanwhile, HIF1A activates VEGF promoter, and this mechanism was blocked when cotransfected with WT-CITED2 in hypoxia condition (Fig. 2e) (1% O2). Mutant T257A had higher luciferase activity than WT-CITED2, and Q117L showed little difference compared with WT-CITED2. Abnormal luciferase level of PITX2C and VEGF indicated that the mutants impair the transcriptional activities of CITED2.
3D protein structure analysis showing a loss of hydrogen bond between Thr257 and Asp254
To further predict the functional changes of these mutations, a C-terminal protein (216–259) 3D model was constructed using SPDBV software (Fig. 3a). In normal condition, Thr257 connects with 254Asp by two hydrogen bonds, while mutant Ala257 has only one hydrogen bond connecting to 254Asp (Fig. 3b, c). Due to the great significance of hydrogen bond in maintaining protein structures, this change may lead to the decreased stability of CITED2 protein and subsequently have impact on its biological functions to some degree.
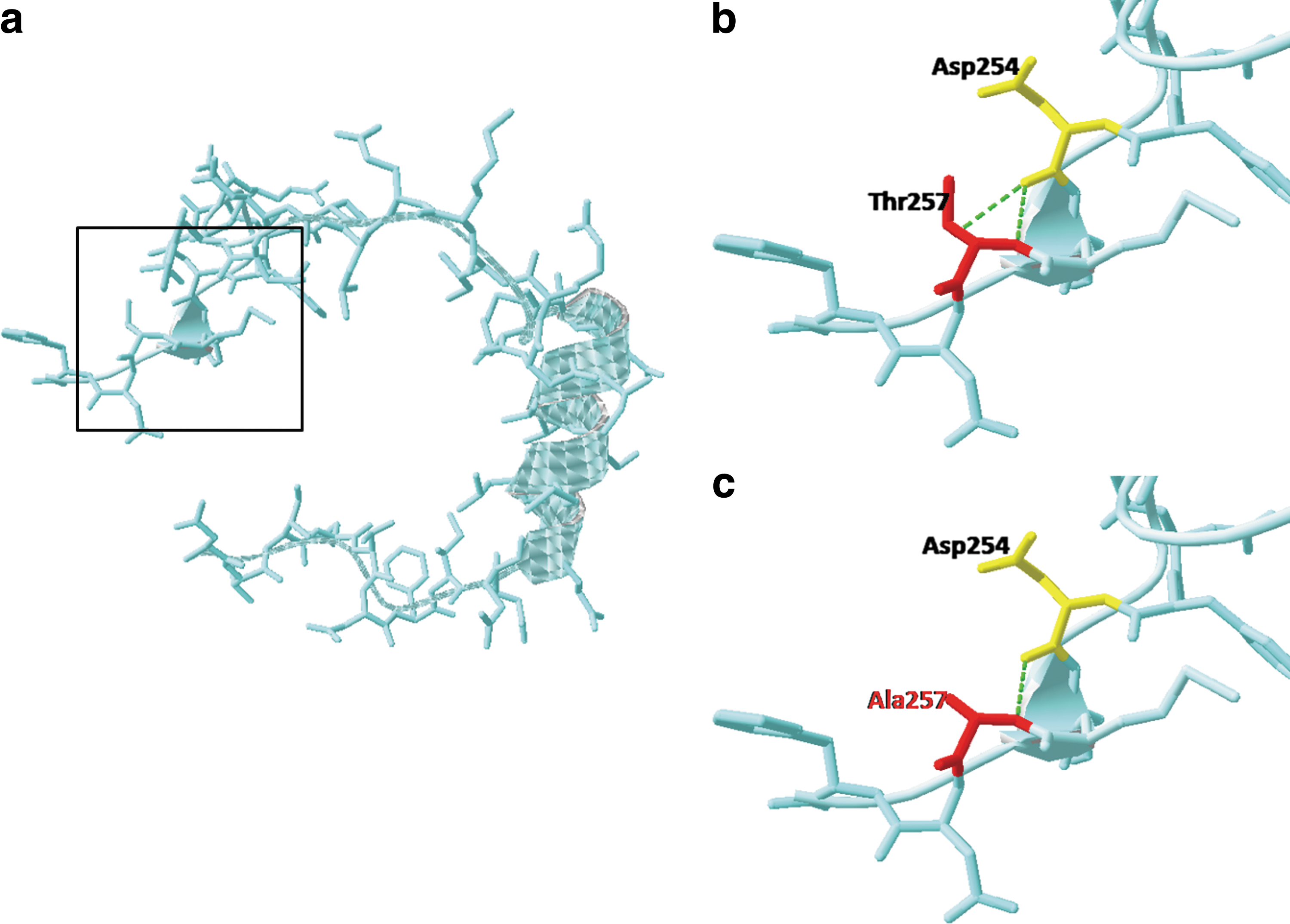
Three-dimensional molecular conformation of CITED2 aa216–259.
T257A encoded unstable proteins and led to low expression of CITED2
We first performed qRT-PCR to figure out if the mutations could alter the expression of mRNA. The results indicated that both Q117 and T257A had little impact on CITED2 transcriptional expression (Supplementary Fig. S1; Supplementary Data are available online at
Discussion
CITED2, a transcriptional activator which cooperates with many transcription factors through its CR2 binding to P300-CH1, has been identified significantly in the normal development of cardiovascular tissue and anatomical structure. CITED2 knockout mice showed various kinds of cardiovascular malformations, including ASD, VSD, left–right axis abnormity, and CTDs. Many variants in CITED2 have been identified to take the responsibility of CTDs in recent years.
Six hundred five Chinese CTD patients and 300 unrelated controls were involved in our study. We screened the coding region of CITED2 and found two mutations (Q117L and T257A) in CTD patients and predicted disease-causing. Q117L locates in CITED family conserved region 1 and T257A in conserved region 2. Multiple sequence alignments also found that these two mutations were highly conserved among different species. This result indicates that the two amino acids may be of great importance in the evolutionary process; mutations at these sites are likely to result in decreased biological function of protein.
We carried out western blotting and found that the protein expression of T257A was obviously lower than WT-CITED2 by calculating the gray intensity ratio of CITED2/β-actin. Meanwhile, Q117 also had reduced expression level. Glutanine (Q) and threonine (T) are classified as polar hydrophilic amino acids, and their replacements leucine (L) and alanine (A) are nonpolar hydrophobic amino acids. Fluctuations in the nature of these amino acids lead to decline in protein stability and might activate the endogenous protein degradation pathway, leading to reduced protein expression.
In vivo, CITED2 recruits TFAP2C by binding to P300 and increases the expression of TFAP2C-downstream PITX2C. PITX2C controls cell proliferation and is involved in morphogenesis during embryonic development (Dagle et al., 2003). PITX2C knockout mice showed valve defects, body wall dysraphism, gastroschisis, ectopia cordis, and other multiple organs polymorphous defects (Bamforth et al., 2004). CITED2 also competes with HIF1A for binding to P300 to reduce the expression of HIF1A and HIF1A-induced gene VEGF; both HIF1A and VEGF are closely related to cardiovascular growth. Previous study showed that overexpression of VEGF may lead to leaky, tortuous, dilated, and saccular vessels of OFT and interventricular septum (Xu et al., 2007). To further explore if the mutants could impact the ability of CITED2 cooperative function, we performed dual-luciferase reporter assay. Our study showed that both Q117L and T257A had lower abilities to coactivate TFAP2C, as a result, expression of PITX2C was reduced. Meanwhile, T257A weakened the inhibitory effect of CITED2 on HIF1A, leading to increased expression of downstream VEGF. Q117L showed little difference with WT-CITED2. So, we draw the conclusion that CITED2 mutations may influence CITED2 transcriptional activity, leading to abnormal expression of downstream genes, and we speculated that the impaired biological function had a relationship with a decline in CITED2 protein expression.
CITED2 is an unstable nuclear protein; almost all the CITED2 must combine with the P300 to play its biological function. David M. Livingston's study indicated that the carboxy-terminal region of CITED2, extending from aa215 to 270, with a core region of 32 amino acids (224–255), was necessary for p300–CH1 binding (Bhattacharya et al., 1999). The stability of CR2 domain protein, in particular the core region (224–255), relates to the normal biological function of CITED2. 3D structure analysis of CITED2 aa216–259 showed that Thr257 binds with Asp254 through two hydrogen bonds, while mutant Ala257 and Asp254 have only one. Hydrogen bond is a kind of secondary bond and is widespread in biological macromolecules. Hydrogen bond has been seen to play an important role in maintaining the secondary and tertiary structures and some physicochemical properties of protein. Changes in the spatial structure and physicochemical properties of CR2 proteins might not only give rise to T257A decreased protein stability but also damage its ability to bind with p300.
The declined protein stability found in our study may be due to physicochemical property alteration and molecular conformation variety of conserved region amino acids. Unstable proteins could start up protein degradation pathway, resulting in a decrease in the expression of CITED2. Reduced protein products along with spatial structure alterations weaken biological function of CITED2, leading to aberrant expression of CITED2-modulated genes such as PITX2C and VEGF. Recent studies indicated that CITED2 can also act as a cofactor of ISL1, another gene associated with cardiovascular development and CTDs (Osoegawa et al., 2014). CITED2 and ISL1 proteins interact physically and coordinate with each other to promote ESC differentiation toward cardiomyocytes (Pacheco-Leyva et al., 2016). It is worthwhile to explore whether CITED2 mutation could affect the interaction between CITED2 and ISL1 and lead to CTDs.
Summarily, by analyzing the biological and functional changes of two CITED2 mutations (Q117L and T257A), our study provided potent evidence that CITED2 gene is related to CTDs and CITED2 mutations in conserved regions contribute to the occurrence of CTDs.
Footnotes
Acknowledgments
The study was supported by National Natural Science Foundation of China (81670210); Shanghai Municipal Commission of Health and Family Planning three-year action plan (GWIV-23); the Science Committee of Shanghai (13J1401705).
Disclosure Statement
No competing financial interests exist.
References
Supplementary Material
Please find the following supplemental material available below.
For Open Access articles published under a Creative Commons License, all supplemental material carries the same license as the article it is associated with.
For non-Open Access articles published, all supplemental material carries a non-exclusive license, and permission requests for re-use of supplemental material or any part of supplemental material shall be sent directly to the copyright owner as specified in the copyright notice associated with the article.