
Abstract
Zeaxanthin epoxidase (ZEP) plays important roles in plant response to various environmental stresses by involving in abscisic acid (ABA) biosynthesis and xanthophyll cycle. A full-length cDNA of PeZEP was isolated from moso bamboo (Phyllostachys edulis), which comprised of a 138-bp 5′-untranslated region (UTR), a 381-bp 3′-UTR, and a 2013-bp open reading frame (ORF) encoding a putative protein of 670 amino acids. PeZEP was mainly expressed in leaf blades and leaf sheaths, and less in roots and culms. The transcript level of PeZEP in bamboo leaf was elevated with the increasing light intensity. PeZEP was significantly upregulated in response to high light (HL: 1200 μmol·m−2·s−1) and reached to a higher level after 1 h treatment, and kept higher levels in the following hours. Besides, PeZEP was upregulated under high temperature (42°C), and downregulated under low temperature (4°C) and exogenous ABA treatment. The expression vector of PeZEP driven by CaMV 35S was constructed and transformed into Arabidopsis thaliana. The transgenic plants overexpressing PeZEP were generated and subjected to drought stress for morphological and physiological assays. Compared with Col-0, the transgenic plants demonstrated enhanced tolerance to drought stress, which appeared later wilting and higher survival rate. Moreover, higher value of F v/F m, higher activities of superoxide dismutase, peroxidase, and catalase, and lower concentration of malondialdehyde were also observed in transgenic plants. Transcript levels of AtP5CS and AtRD29b related to drought stress were enhanced in transgenic plants. These results indicated that PeZEP might play an important function in response to drought stress in bamboo.
Introduction
B
The xanthophyll cycle, in which the de-epoxidation for violaxanthin (V) to zeaxanthin (Z) through antheraxanthin (A) is favored under high light conditions, whereas epoxidation of Z to V predominates in moderate light conditions (Arnoux et al., 2009; Gonzalez-Jorge et al., 2016), is considered to be one of the critical processes of plant fitness and stress tolerance. The violaxanthin de-epoxidase (VDE) and zeaxanthin epoxidase (ZEP) are two key enzymes in the xanthophyll cycle. The transcript levels of PeVDE was upregulated in leaves exposed to HL, and the activity of PeVDE was validated by catalyzing V conversion to Z through A in vitro (Gao et al., 2013), suggesting PeVDE has a critical function in the protection of bamboo photosynthetic apparatus from light stress. However, the function of ZEP in bamboo is still unknown.
As a conserved lipocalin, ZEP catalyzes the conversion of Z into V (Flower et al., 2000), which is not only an essential part of ABA biosynthesis, but also an important part of carotenoid biosynthesis and the xanthophyll cycle (Nambara and Marion-Poll, 2005; DellaPenna and Pogson, 2006). The ABA biosynthesis and xanthophyll cycle are two important processes for plant acclimation to environmental stress conditions. In Arabidopsis thaliana, ZEP was tissue- and stress-specific accumulation in accordance with its functions in ABA biosynthesis and the xanthophyll cycle (Schwarz et al., 2015), and the transcript levels of ABA1/ZEP was upregulated in seed with the synthesis of ABA after exposure to high temperature (Toh et al., 2008). An apparent correlation between the expression of ZEP and the increased content of ABA was observed in wounding potato tubers (Suttle et al., 2013). The expression level of ZEP was increased in tomato roots, but not leaves during drought stress, whereas antisense downregulation of LeZEP1 causes accumulation of zeaxanthin in leaves (Thompson et al., 2000). The transgenic line of A. thaliana with partly disabled ZEP activity displayed changed carotenoid composition, xanthophyll cycle activity, and nonphotochemical quenching kinetics, indicating that ZEP activity resulted in an altered xanthophyll profile (Nowicka et al., 2009). Heterologous expression of MsZEP could enhance drought and salt tolerance of transgenic tobacco (Zhang et al., 2016). Therefore, ZEP plays important roles in response to various stresses through different pathways. To understand the characteristic and function of ZEP in bamboo, the full-length cDNA of PeZEP and its promoter were isolated from moso bamboo (Phyllostachys edulis) in the present study. Based on the bioinformatics analyses, the analyses of tissue-specific and expression patterns in bamboo under different stresses were conducted. Moreover, the biological functions of PeZEP in response to drought stress were investigated by heterologous expression of the gene in A. thaliana.
Materials and Methods
Plant materials and stress treatments
The potted seedlings of moso bamboo (P. edulis) were grown in a greenhouse at 25°C under controlled light condition (A photoperiod of 16-h light/8-h dark and a light intensity of 250 μmol·m−2·s−1). To examine the tissue-specific gene expression patterns, samples of leaf blades, leaf sheaths, stems, and roots were collected from 1-year-old seedlings.
When the moso bamboo seedlings were 6 months old, the plants were subjected to different stress treatments. The materials dark adapted for 24 h were moved from dark to different light intensities of 0, 300, 600, 900, 1200, and 1500 μmol·m−2·s−1 for 2 h. HL (1200 μmol·m−2·s−1) treatment was carried out and leaves were collected after 0, 1, 2, 4, 8, and 12 h. During the process where seedlings were subjected to drought stress with water control until the leaves became wilted (about 13 days), leaves were sampled at 0, 3, 5, 7, 9, 11, and 13 days. For temperature treatments, the seedlings were placed into growth chambers controlled at 4°C or 42°C, leaves were collected after 0, 0.5, 1, 2, 6, and 12 h, respectively. After uprooting and washing with sterile water, the seedling roots were immediately submerged in solution of 10 μM ABA, roots were collected after 0, 1, 2, 4, 8, and 12 h, respectively. All the collected samples were frozen immediately in liquid nitrogen and stored at −80°C.
RNA and genomic DNA extraction
Total RNAs were extracted from the abovementioned samples using TRIzol Reagent according to the protocol recommended by the manufacturer (Invitrogen), followed by the treatment of RNase-free DNase I (Promega) for 30 min at 37°C to remove residual DNA. The RNA concentration and quality were checked with NanoDrop 2000 (Thermo) and the qualified RNAs were used for the synthesis of first-strand cDNA with the Reverse Transcription Kit following the manufacturer's protocol (Promega). Moso bamboo leaves were used to extract the genomic DNAs using the cetyltrimethylammonium bromide method (Gao et al., 2006).
Gene and its promoter cloning
Based on the conserved sequences of ZEP homologous genes from monocots (such as Oryza sativa, Brachypodium distachyon, and Zea mays), two degenerate primers (ZEP-F and ZEP-R) were designed and used to amplify the conserved sequence in moso bamboo. The PCR products were gel purified, cloned into the pGEM®-T Easy vector (Promega), and subsequently sequenced by Sangon Biotech Co., Ltd. (Shanghai, China). According to the obtained sequence, specific primers (ZEP5-1 and ZEP5-2, ZEP3-1, and ZEP3-2) were designed for 5′ and 3′ rapid amplification of cDNA ends (RACE) to obtain the full-length cDNA of PeZEP. The RACE reactions were conducted using the specific primers with the SMART™ RACE cDNA Amplification Kit (Clontech). The full-length cDNA of PeZEP was obtained by assembling the conserved sequence with 5′ and 3′ end sequences. Based on the assembled sequence, the primers PeZEP-F and PeZEP-R were designed and used for the cloning of the ORF sequence. Both the cDNAs and genomic DNAs were used as templates in PCRs for amplification of PeZEP using the Pyrobest DNA polymerase (TaKaRa, Japan) with primers PeZEP-F and PeZEP-R. The promoter region of PeZEP, 1429 bases upstream to the start codon, was amplified using the primer pair PeZEPp-F and PeZEPp-R designed on the basis of the genomic sequence (PH01001603G0200 in BambooGDB) (Zhao et al., 2014). Nucleotide sequences of the PCR products and cloned cDNAs were determined by sequencing. The sequences of all relevant primers are listed in Supplementary Table S1 (Supplementary Data are available online at
Bioinformatics analysis
The nucleotide sequence, deduced amino acid sequence, molecular weight and isoelectric point (pI), and ORF of PeZEP were analyzed using bioinformatic tools at the websites (
Quantitative real-time PCR analysis
The expression pattern of PeZEP was assessed using quantitative real-time PCR (qRT-PCR) with specific primers PeZEPq-F and PeZEPq-R. The qRT-PCR was performed with the Roche Light Cycler 480 SYBR Green 1 Master Kit on a qTOWER2.2 system (Analytik Jena, Germany). The qRT-PCR procedure consisted of 95°C for 10 min and 40 cycles of 95°C for 10 s, and 60°C for 10 s. The reaction volume was 10.0 μL containing 5.0 μL of 2 × SYBR Green 1 Master, 0.8 μL cDNA, 0.2 μL of primer (5.0 mM, each), and 3.8 μL ddH2O. Calibration curves were obtained by amplification using a fivefold serial dilution of each cDNA sample, the correlation coefficients (R2) of curves was 0.994 and the PCR efficiency value was 0.98. Triplicate qRT-PCRs were performed for each sample, and the qRT-PCR experiments were performed as biological triplicates. The relative expression level of PeZEP was calculated with 2−ΔΔCt method (Livak and Schmittgen, 2001) using both NTB and TIP41 as the reference genes (Fan et al., 2013).
Vector construction and plant transformation
The ORF of PeZEP was amplified using primers PeZEP-F1 and PeZEP-R1 to introduce a BamHI site upstream of the start codon and a SacI site downstream of ORF. The amplification products were inserted into pBI121 vector (Clontech) digested with BamHI and SacI to generate the CaMV 35S::PeZEP construct. The construct was transformed into Agrobacterium tumefaciens EHA105 by electroporation (Main et al., 1995), and then transformed into Arabidopsis thaliana (Col-0) using the floral dip method (Clough and Bent, 1998). Putative transgenic plants were selected on MS (Murashige and Skoog, 1962) solid medium containing 30 mg/L kanamycin, which were further validated by semiquantitative RT-PCR using AtUbiquitin (NM180850) as a positive control (Yang et al., 2015). Three lines of T3-resistant seedlings showing no evidence of segregation were used in subsequent analyses.
To examine the activity of PeZEP promoter, the promoter sequence upstream of PeZEP was inserted into pBI101 vector (Clontech) digested with HindIII and XbaI sites to generate the vector for transient expression of GUS in tobacco leaves. Histochemical β-glucuronidase assays were carried out after two days following the protocol of the kit (Real-Times, China). Images were recorded with a digital camera (Canon EOS 700D, Japan).
Stress treatments of transgenic arabidopsis plants
To test stress tolerance of PeZEP transgenic Arabidopsis plants, 30-day-old seedlings were subjected to HL, chilling, and drought stress treatments. The seeds of Col-0 and transgenic plants were incubated in dark at 4°C for 2 days, and then grown in pots containing vermiculite and peat moss (1:1) under a long-day photoperiod (16-h light/8-h dark) at 22°C for about 3 weeks. For the HL or low temperature treatments, the plants were exposed to 1200 μmol·m−2·s−1 or 4°C conditions. The samples were collected after 1, 2, 3, and 4 h, respectively. For drought tolerance experiment, water was withheld for 25 days, and then the plants were rewatered for 7 days. The number of plants that survived after rehydration was recorded.
Measurement of chlorophyll fluorescence parameters
The chlorophyll fluorescence parameters, such as F v/F m and non-photochemical quenching (NPQ), are important indices to reflect the photoinhibition degree of plants under stresses. In vivo chlorophyll fluorescence parameters of T3 transgenic Arabidopsis plants (3-week-old) were measured using the Imaging-PAM fluorometer device (Walz, Germany) in combination with the ImagingWin software. Before measurements, the plants were dark adapted for 30 min. The parameters of F v/F m and NPQ were calculated as (F m − F o)/F m and (F m − F m′)/F m′, respectively (Maxwell and Johnson, 2000).
Measurements of antioxidant enzyme activities and malondialdehyde content and detection of reactive oxygen species
After drought treatment for 15 days, the activities of superoxide dismutase (SOD), catalase (CAT), and peroxidase (POD) in leaves were measured. For SOD, hydroxylamine method was used by measuring the inhibiting rate of the enzyme to O2 − using an assay kit (A001-1; Jiancheng, China); 1 U was defined as the amount of enzyme required to produce 50% inhibition of reduction of nitrite in 1.0 mL reaction solution by measuring the change of absorbance at 550 nm. For CAT, an assay kit (A007-1; Jiancheng) was used. Based on the hydrolysis reaction of H2O2 with CAT, 1 U was defined as the quantity of CAT required to decompose 1 μmol of H2O2 in 1.0 mg fresh tissue by measuring the decrease of absorbance at 405 nm. For POD, an assay kit (A084-3; Jiancheng) was used. According to the change of absorbance at 420 nm by catalyzing H2O2, 1 U was defined as the quantity of POD catalyzed and generated 1 μg substrate by 1 mg fresh tissues at 37°C. For measurement of malondialdehyde (MDA) content, thiobarbituric acid (TBA) method was used based on the reaction of TBA and MDA using an assay kit (A003-1; Jiancheng), which was measured by the absorbance of red TBA-MDA complex at 532 nm. Based on the average optical density (AOD) values, formulae for the calculation are listed in Supplementary Data S1. For the detection of reactive oxygen species, histochemical staining of superoxide and hydrogen peroxide were performed using modified Nitro Blue Tetrazolium (NBT) and 3, 3′-diaminobenzidine (DAB) staining methods, respectively (Zulfugarov et al., 2014).
Expression analysis of stress-related genes
Furthermore, the transcript levels of two stress-responsive genes, AtP5CS (1-pyrroline-5-carboxylate synthetase gene, AT3G55610) and AtRD29b (Responsive to desiccation29B gene, AT5G52300), were further analyzed in the Col-0 and transgenic plants. AtUBQ (Ubiquitin, NM_116771) was used as reference gene (Chen et al., 2015). The primers for qRT-PCR are shown in Supplementary Table S1.
Statistical analyses
Analyses were performed with SPSS Statistics for Windows (Version 17.0.; SPSS, Inc.). Figures were created using GraphPad Prism for Windows (Version 5.01.; GraphPad, Inc.). All data are presented as the mean ± standard error of three replications. One-way analysis of variance was used for the evaluation of the statistical significance of differences among means using the software in SPSS. Single and double asterisks indicate significant differences at the levels of p < 0.05 and p < 0.01, respectively.
Results
Isolation and characterization of PeZEP
A cDNA fragment of 450-bp was obtained from bamboo (P. edulis) leaves using the degenerate primers (ZEP-F and ZEP-R). This fragment was verified as a part of ZEP in bamboo using a BLAST search in GenBank database, and then the 5′ and 3′-RACE were performed to obtain the full-length cDNA sequence. It was 2532-bp, containing a 2013-bp ORF, a 138-bp 5′-untranslated region (UTR), and a 381-bp 3′-UTR. The ORF was determined by end-to-end PCR, which encodes a putative peptide of 670 amino acids with a predicted relative molecular mass of 73.4 kDa and a theoretical pI of 7.66, respectively. The gene was named as PeZEP and deposited in GenBank (KP274885). The genomic sequence corresponding to the ORF of PeZEP was obtained, which was 6094-bp in length containing 16 exons and 15 introns (Supplementary Fig. S1). Besides, there were two PeZEP homolog genes PeZEP1 (PH01001603G0200) and PeZEP2 (PH01000571G0610) in bamboo genome database by searching the whole genomic sequence (Peng et al., 2013). Sequence alignment analysis showed that PeZEP had high identities with PeZEP1 (91.2%) and PeZEP2 (86.9%) with the main difference in the N-terminal (Data no shown).
Alignment and phylogenetic analysis
The results of BLASTP indicated that the deduced amino acid sequence of PeZEP shares high homology with the ZEPs in other plants and algae. For example, PeZEP exhibited 65.8% identity with that of A. thaliana, and more than 80.0% with those of gramineous plants (O. sativa 89.8%, B. distachyon 86.8%, and Z. mays 85.7%). Multiple sequence alignment showed that ZEP is highly conserved among different plant species, they all contain four conserved domains, a long monooxygenase domain, two lipocalin conserved motifs and a Forkhead-associated domain (Fig. 1), which is consistent with the characteristics of lipocalin superfamily (Flower et al., 2000).
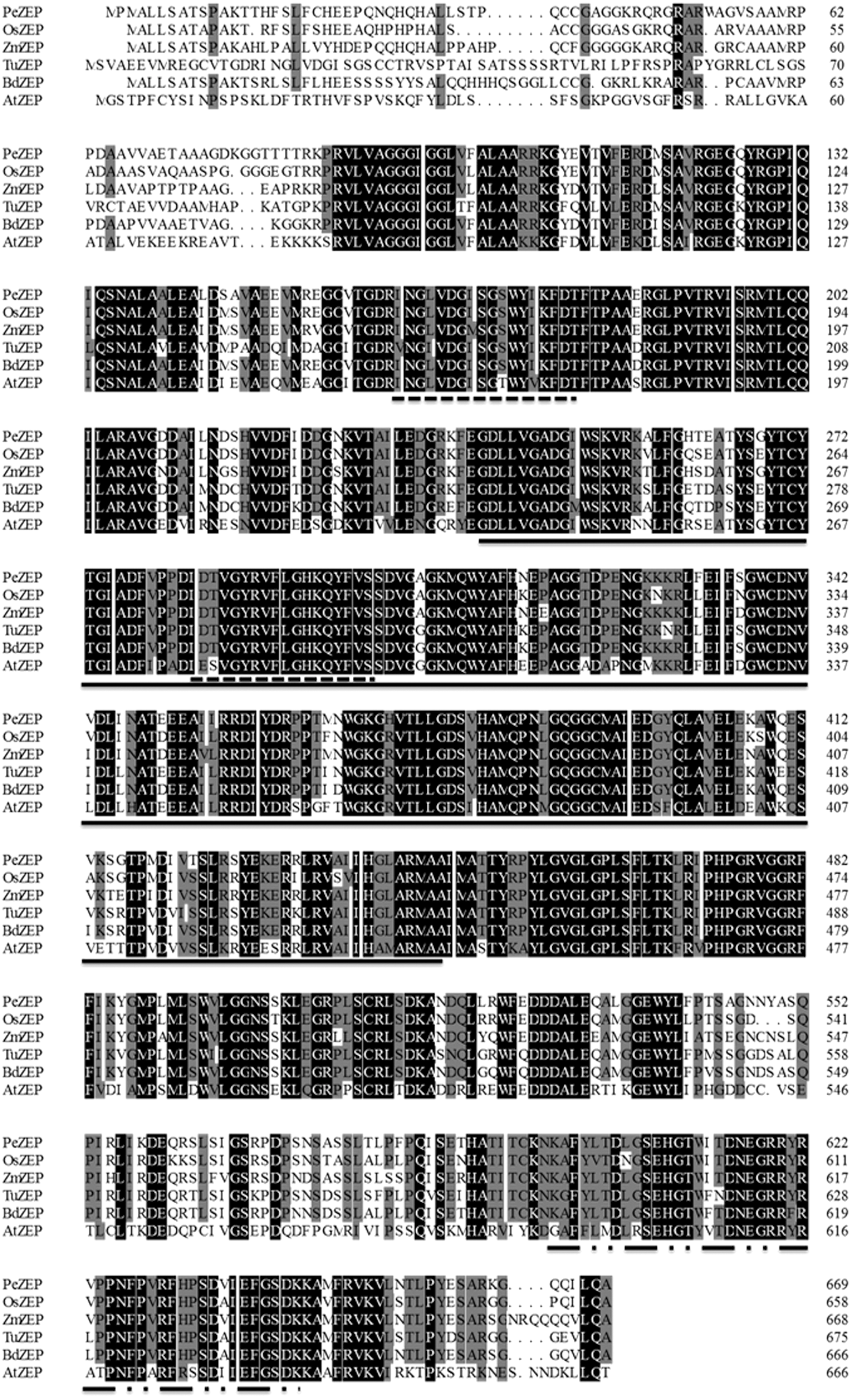
Alignment of deduced amino acid sequences of ZEP from six different plant species. The two lipocalin conserved motifs are highlighted with dash lines, the long monooxygenase domain and the Forkhead-associated domain are marked with solid line and dot–dash line, respectively. Identical amino acid residues are shaded in black. The similar residues in more than 80% of all the sequences are shaded in gray. At, Arabidopsis thaliana; Bd, Brachypodium distachyon; Os, Oryza sativa; Pe, Phyllostachys edulis; ZEP, zeaxanthin epoxidase; Zm, Zea mays.
To investigate the genetic relationship of ZEPs among different species, a phylogenetic tree was constructed based on the amino acids of PeZEP and ZEPs in other plants. As a result, ZEPs were grouped into three clusters that discriminated monocots, dicots, and algae. PeZEP was grouped in the same cluster with other monocots, including O. sativa, Z. mays, and B. distachyon, among which the closest one is that of O. sativa. ZEPs from algae not only have distant relationships with those in higher plants, but also have less similarity with each other (Fig. 2). This result is consistent with the existing plant phylogeny and classification.
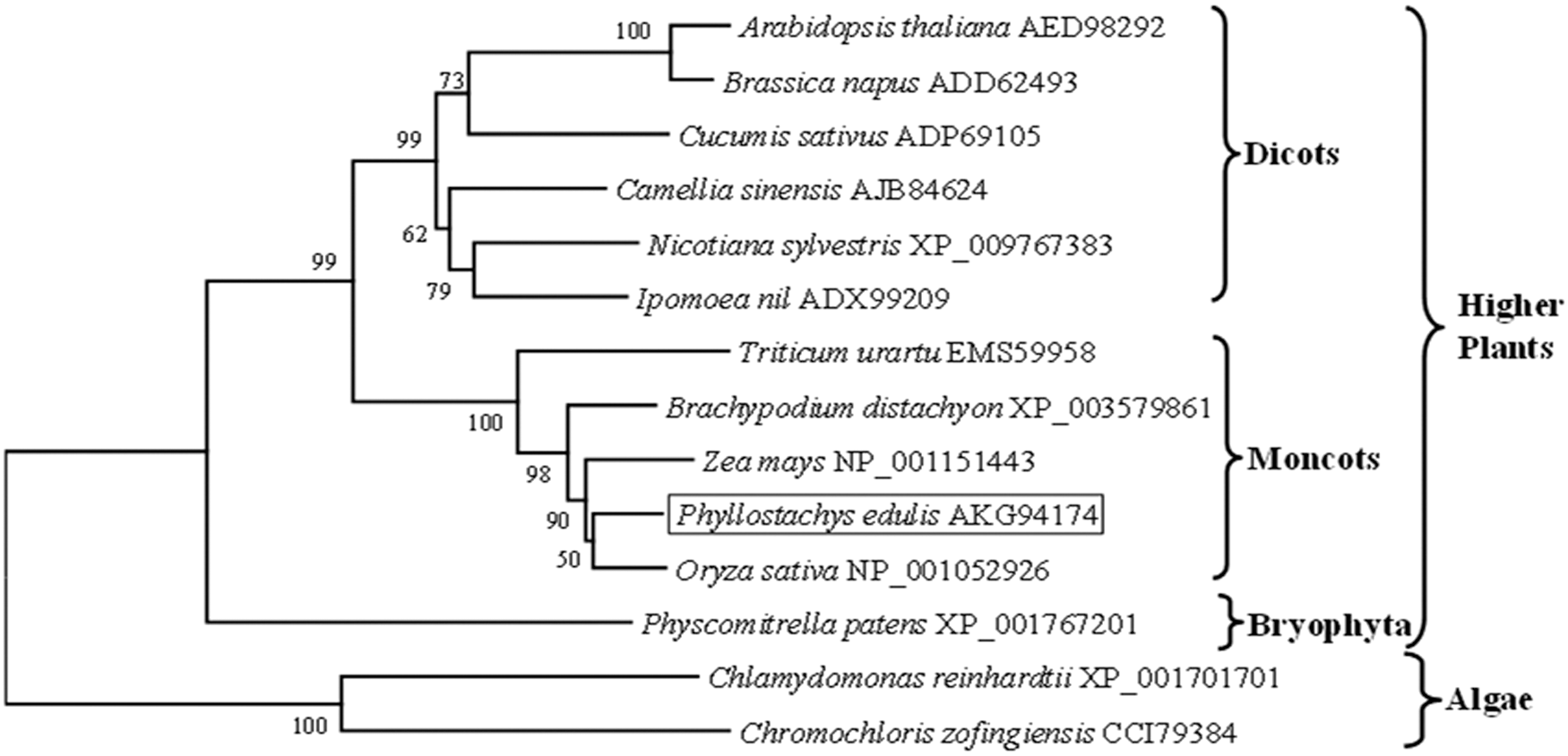
Phylogenetic analysis of PeZEP with other related ZEPs based on amino acid sequences using MEGA 6.0. The number beside the branches indicated the bootstrap values based on 1000 replicates.
Expression patterns of PeZEP in moso bamboo
Spatial expression patterns of PeZEP in moso bamboo were examined using qRT-PCR. As shown in Figure 3, PeZEP was differently expressed in all tissues, the abundant transcripts of PeZEP were detected in leaf blades and leaf sheaths, and less accumulation in roots and stems. The transcript level of PeZEP in leaf blades was approximately eight times higher than that in roots. Similar results could be seen in Supplementary Fig. S2 using the second reference gene (TIP41).
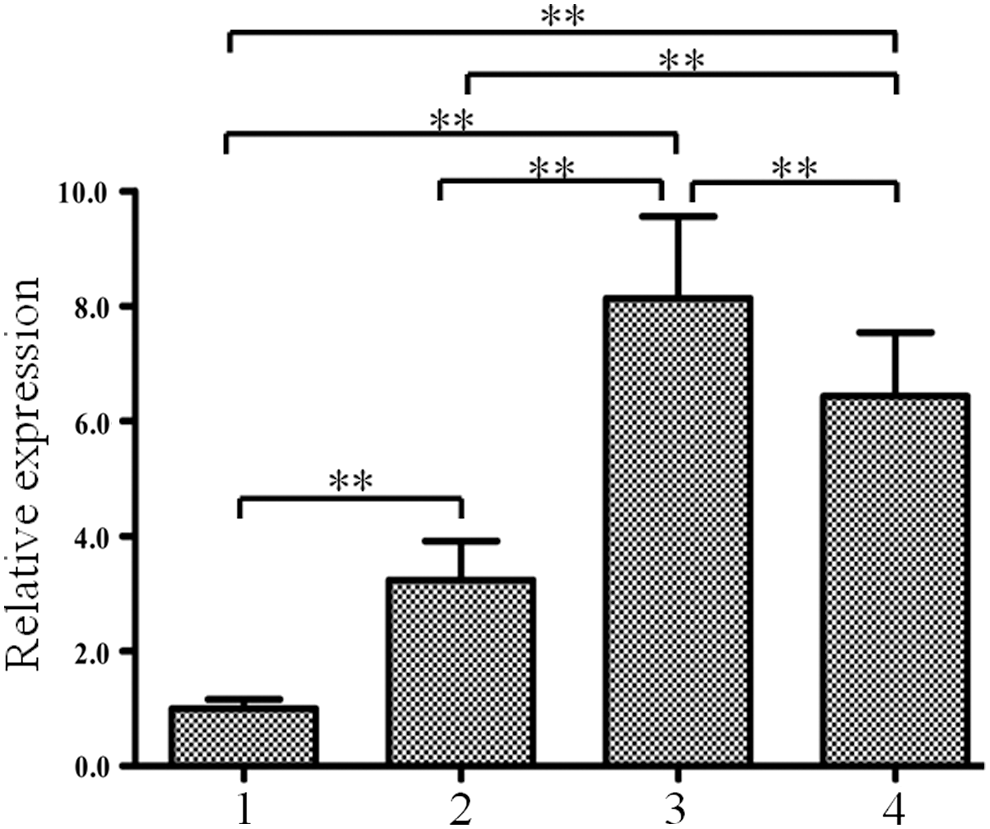
Expression analysis of PeZEP in different tissues of moso bamboo using qRT-PCR. 1, roots; 2, stems; 3, leaf blades; 4, leaf sheaths. The transcript levels of PeZEP were normalized to that of PeNTB. Asterisks indicate the significant difference at **p < 0.01. qRT-PCR, quantitative real-time PCR.
The expression patterns of PeZEP in response to light, drought, chilling, heat, and ABA stresses in leaf blades were analyzed using qRT-PCR, respectively. Because many carotenoid biosynthesis genes can be induced by light, we analyzed the accumulation of PeZEP transcripts in response to different intensities of light (0–1500 μmol·m−2·s−1). As shown in Fig. 4A, the transcript level of PeZEP was gradually upregulated with the light intensity increasing and peaked at about 1200 μmol·m−2·s−1 (about sixfold of the control). Under drought treatment, the transcript level of PeZEP fluctuated slightly in leaf blades with the prolonged time, and there were no significant differences compared with the control (Fig. 4B). Under the HL (1200 μmol·m−2·s−1) stress, PeZEP increased rapidly and reached a higher level after 1 h, and then it kept higher levels accompanied by fluctuations in the following hours (Fig. 4C). The transcript level of PeZEP in leaf blades was gradually downregulated and reached a minimum at 12 h (0.35-fold of the control) when the seedlings were subjected to ABA (10 μmol) treatment (Fig. 4D). Low temperature (4°C) induced the expression of PeZEP slightly in the early stage (0.5 h), but PeZEP was downregulated quickly with the prolonged treatment, and reached a minimum at 12 h (about 0.2-fold of the control) (Fig. 4E). In addition, different from the low temperature treatment, the expression of PeZEP was upregulated slightly during the whole process of high temperature (42°C) (Fig. 4F), in which the maximum appeared after 1-h treatment (about 1.53-fold of the control). The second reference gene (TIP41) was also used to validate the expression patterns of PeZEP (Supplementary Fig. S3), which demonstrated very consistent results with those of the former using NTB as native control.
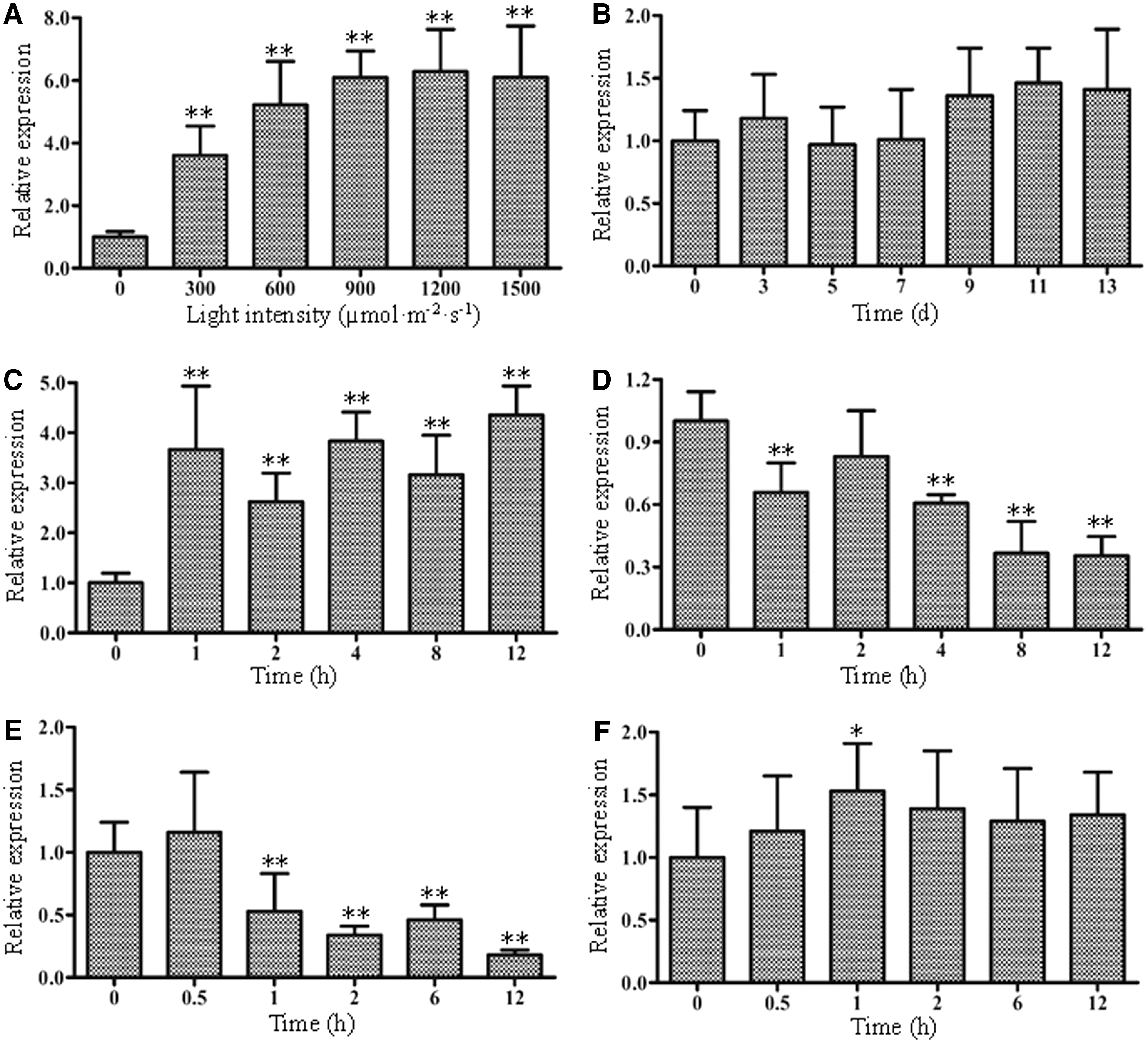
Expression analysis of PeZEP in moso bamboo leaves under different treatments using qRT-PCR. PeNTB was used as the reference gene.
Transient expression of PeZEP promoter in tobacco leaves
A 1429-bp fragment upstream of PeZEP translation start codon ATG was isolated from the genomic DNA of moso bamboo, in which the essential promoter elements of the putative transcriptional initiation site, TATA box, and CAAT box were found, indicating that it may be the promoter (PRO) of PeZEP (Supplementary Data S2). Besides, many cis-regulatory elements, such as AAAC-motif, ACE, G-Box, LTR, and MBS (Supplementary Table S2), were detected, which supported that the transcript levels of PeZEP might be affected by stresses. The construct of PRO::GUS was generated and transformed into tobacco leaves through the Agrobacterium-mediated transformation for transient expression analysis. The results showed that GUS activity was detected in tobacco leaves (Supplementary Fig. S4), indicating that PRO was able to initiate the transcription of GUS gene. The role of PRO in regulating PeZEP expression still needs to be studied.
Generation of PeZEP transgenic arabidopsis plants
To investigate whether heterologous expression of PeZEP is functional, overexpression vector of PeZEP was constructed and transformed into A. thaliana (Col-0). After kanamycin resistance was screened and verified by PCR, seven transgenic lines were obtained. No obvious phenotype differences were observed between those of Col-0 and transgenic plants under normal conditions. The transcript levels of PeZEP in three T3 generation transgenic lines (L1, L2, and L4), without resistance segregation, were determined using semiquantitative RT-PCR. The results showed that PeZEP was expressed in the three transgenic lines with slightly lower in L2 (Supplementary Fig. S5). Therefore, L1, L2, and L4 were selected for further analysis.
Chlorophyll fluorescence kinetics in transgenic arabidopsis plants under light stresses
To evaluate the role of the xanthophyll cycle in protecting the photosynthesis apparatus from excess light energy in transgenic Arabidopsis plants, the values of F v/F m and NPQ were determined. The values of F v/F m in transgenic plants decreased obviously compared with that in Col-0 plants under HL stress (1200 μmol·m−2·s−1) (Fig. 5). After HL stress for 4 h, the F v/F m value of Col-0 plants decreased about 46.17%, whereas those of transgenic lines decreased about 54.01% (L-4), 61.5% (L-1), and 65.62% (L-2), respectively. After HL stress, both Col-0 and transgenic plants were transferred to weak light conditions (100 μmol·m−2·s−1), and the values of F v/F m were all recovered. However, the recovery of F v/F m in the Col-0 plants was faster than those of transgenic plants. After 8 h, the F v/F m value in Col-0 plants could recover for 81.41% of the initial value, whereas those in the L-1, L-2, and L-4 only reached to 76.91%, 74.82%, and 72.02%, respectively. Meanwhile, the values of NPQ in Col-0 and transgenic plants demonstrated very similar with an increasing trend under HL stress and a downward trend during the recovery, but there were no significant differences between the Col-0 and transgenic plants (Fig. 5).
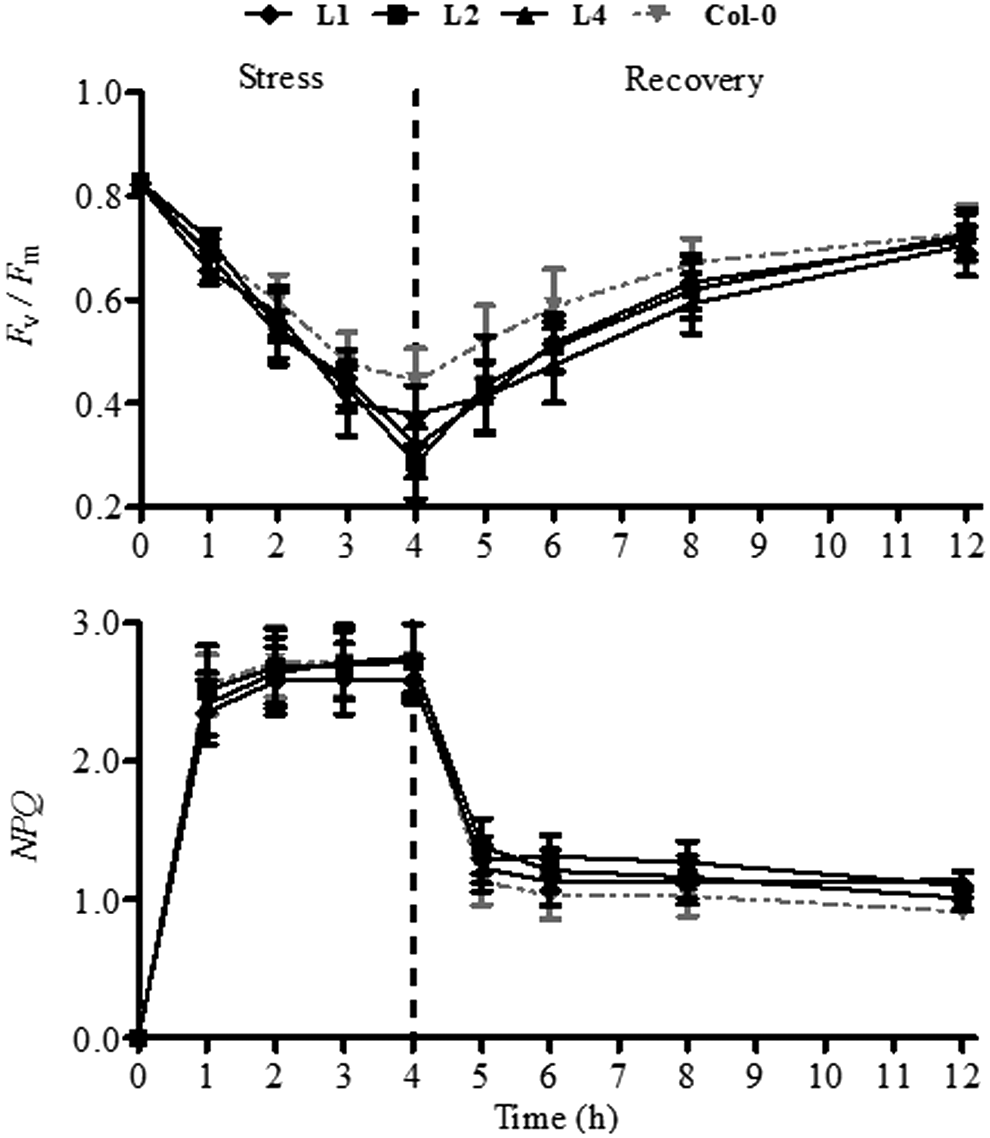
Changes of F v/F m and non-photochemical quenching (NPQ) in Col-0 and PeZEP transgenic plants during the HL treatment and subsequently recovery under weak light conditions.
Comparison of NPQ kinetics in Col-0 and transgenic plants induced by low light (LL: 100 μmol·m−2·s−1), moderate light (ML: 500 μmol·m−2·s−1), or HL (1000 μmol·m−2·s−1) was performed, and the NPQ data were displayed as false color images. The result indicated that the actinic light could induce a rise of NPQ in darkness-adapted leaves of both Col-0 and transgenic plants. The Col-0 leaves tended to red and those of L4 tended to yellow under LL, and the Col-0 leaves tended to green and those of L4 tended to blue under ML, whereas the leaf color of Col-0 and L4 was similar under HL (Fig. 6A). The similar results were also observed in the transgenic plants of L1 and L2 (data not shown). Besides, the NPQ induction and relaxation kinetics exhibited extremely different under the different light conditions. When the actinic light was set as LL and ML, the transgenic plants exhibited a faster NPQ kinetics in the light phase and a slower relaxation in the dark phase compared with Col-0 plants. However, if actinic light was set as HL, there was no obvious difference in the NPQ kinetics between Col-0 and transgenic plants (Fig. 6B).

NPQ of Col-0 and PeZEP transgenic plants induced by different light intensities. LL: 100 μmol·m−2·s−1; ML: 500 μmol·m−2·s−1; HL: 1000 μmol·m−2·s−1.
Drought tolerance of transgenic arabidopsis plants
To explore whether overexpression of PeZEP could confer drought stress tolerance in transgenic Arabidopsis plants, both Col-0 and transgenic plants were subjected to drought stress treatment. It was found that transgenic plants were more tolerant to drought than Col-0 plants. Col-0 plants were slightly withered after water control for 10 days, whereas nearly all the transgenic plants showed normal phenotype (Data no shown). Both Col-0 and transgenic plants completely withered after drought stress for 25 days (Data no shown). Rehydration was conducted after drought stress, the result indicated that transgenic plants had significant higher survival rate than that of Col-0 plants (p < 0.01) (Fig. 7A).

Effects of drought stress on the Col-0 and PeZEP transgenic plants. PeZEP transgenic lines: L1, L2, and L4.
To examine the physiological changes in PeZEP-overexpressing plants induced by drought, the F v/F m values, MDA content, SOD, POD, and CAT activities were measured in the leaves after drought treatment. The results showed that the values of F v/F m in transgenic plants decreased less than that in Col-0 plants under drought treatment. After drought treatment for 15 days, the F v/F m values of transgenic plants decreased about 13.5% in L1, 12.5% in L2, and 16.3% in L4, whereas that in Col-0 plants was about 17.0% (Fig. 7B). Under the nondrought stress condition, there was no significant differences on MDA content, SOD, POD, and CAT activities between the Col-0 and transgenic plants. However, both the content of MDA and the activities of SOD, POD, and CAT were increased in Col-0 and transgenic plants under drought treatment. The MDA content in transgenic plants was significantly lower (p < 0.01) than that of Col-0 plants (Fig. 7C). The SOD activity in the transgenic plants was significantly higher (p < 0.05) than that of Col-0 plants (Fig. 7D), whereas those of POD and CAT in the transgenic plants were significantly higher (p < 0.01) than those of Col-0 plants (Fig. 7E, F).
In investigation of the antioxidant capacity of PeZEP-overexpressing plants, the levels of superoxide and hydrogen peroxide were measured using histochemical staining with NBT and DAB, respectively. The color of the stained leaves was deeper in Col-0 plants than those of transgenic plants (L1 and L4) (Supplementary Fig. S6), indicating that the accumulated superoxide and hydrogen peroxide in Col-0 plants were more than those in the transgenic plants. This result suggested that the transgenic plants have stronger antioxidant capacity than Col-0 plants.
In addition, the transcript levels of AtP5CS and AtRD29b, two drought stress-related genes, were investigated. The results indicated that the transcript levels of AtRD29b in transgenic plants was higher (p < 0.05) than that of Col-0 plants (Fig. 7G), and no significant differences of AtP5CS was observed between transgenic lines and Col-0 plants under the nondrought stress condition (Fig. 7H). However, the increased transcript levels of AtP5CS and AtRD29b were all observed in transgenic lines under drought stress. The difference levels were significant for AtP5CS in L1 (p < 0.05) and strongly significant for those in L2 and L4 (p < 0.01) (Fig. 7G), those of AtRD29b were significant in L4 (p < 0.05) and strongly significant in L1 and L2 (p < 0.01) after drought treatment (Fig. 7H).
Discussion
With rapid development of molecular biology, large numbers of ZEP genes have been isolated and proven to play important roles in the ABA biosynthesis and xanthophyll cycle both in plants and algae in recent years (Marin et al., 1996; Wang et al., 2008; Eilers et al., 2016; Zhang et al., 2016). Although the functions of ZEP genes have been extensively studied in Arabidopsis and algae (Xiong et al., 2002; Couso et al., 2012; Eilers et al., 2016), the studies are still rare in forest plants. A novel ZEP gene, PeZEP, was identified and characterized from moso bamboo in the present study. Although two ZEP homolog genes (PH01001603G0200 and PH01000571G0610) exist in moso bamboo genome database, both of them failed to be isolated in all bamboo tissues after multiple attempts. We have to isolate the conserved region of the gene using degenerate primers, and for the 5′- and 3′-UTR with the RACE methods. Moreover, some studies had demonstrated that ZEP is a single-copy gene in angiosperms and part of green algae, and two or three copies only existed in some algae classes, such as Phaeodactylum tricornutum and Thalassiosira pseudonana (Frommolt et al., 2008; Goss and Jakob, 2010). Therefore, we hold the view that ZEP in moso bamboo is also a single-copy gene, and the two homolog genes in the database maybe the result of incorrect sequence assembly.
PeZEP encoded a hypothetical protein belonging to lipocalin superfamily, and its length was similar to those described in other plants, but slightly longer than those in algae. Sequence alignment showed that PeZEP shared high identity with ZEP proteins from other plants, including a long monooxygenase domain, two lipocalin conserved motifs and a Forkhead-associated domain. Among this, the deduced amino acid sequences of PeZEP showed the highest homology with ZEP in O. sativa, and clustered together with other monocot plants in the phylogenetic tree. These results indicated that the PeZEP protein is a typical ZEP protein, and it may have the similar function with the ZEPs in other plants. Based on hydrophobicity plots, a putative transmembrane helix has been identified at the N-terminal of PeZEP, which was in agreement with A. thaliana. This result indicated that PeZEP could bind to the thylakoid membrane, which needs to be further validated.
In recent study, the researchers found that the ZEP was a tissue-specific accumulation protein, and the highest levels were present in leaf chloroplasts and root plastids (Schwarz et al., 2015). PeZEP was constitutively expressed in all bamboo tissues, with the most abundant in green tissues, which was similar to previous works for AtZEP and MsZEP (Schwarz et al., 2015; Zhang et al., 2016). Light intensity could affect the expression of genes involved in xanthophyll cycle. PeVDE was upregulated by the increasing light intensity (Gao et al., 2013). Previous studies have shown that the ZEP transcription in leaves is regulated by the diurnal rhythm and not induced by light (North et al., 2005; Wang et al., 2008). Our study showed that the transcript of PeZEP was elevated during the change from dark to light, and it was gradually increased with light intensity increasing and with duration of HL. This result implied that PeZEP could be involved in response to HL in bamboo leaves.
It is well known that ABA is an important hormone of plant response to various environmental stresses, such as drought, cold, and heat (Baron et al., 2012; Sreenivasulu et al., 2012). As an important gene in ABA biosynthesis, the transcript levels of ZEP under drought stress were downregulated in leaves of Arabidopsis, tobacco, and tomato (Audran et al., 1998; Thompson et al., 2000; North et al., 2005). However, the expression of PeZEP showed no significant changes under drought stress, which was consistent with the expression of VuABA1 in cowpea (Iuchi et al., 2000). In alfalfa, MSZEP expression in leaves decreased significantly under both cold and heat stress conditions (Zhang et al., 2016). In the present study, unlike low temperature (4°C) reducing PeZEP expression, PeZEP had no obvious changes under high temperature (42°C), which indicated that high temperature had little influence on PeZEP in leaves. It was reported that the transcript levels of ABA1/ZEP in imbibed seeds of Arabidopsis were upregulated at high temperature (Toh et al., 2008), and exogenous ABA enhanced the expression of ZEP in Arabidopsis (Xiong et al., 2002). However, our data showed that exogenous ABA could downregulate the expression of PeZEP in leaves, which was consistent with the expression of MsZEP in alfalfa (Zhang et al., 2016).
Several studies about overexpression of the ZEP genes have been reported in plants. In tomato, LeZE related to the dissipation of excess excitation energy by participating in NPQ. Overexpression of LeZE impaired the function of the xanthophyll cycle and aggravated photoinhibition in tomato under HL and chilling stress. Antisense suppression of LeZE could alleviate chilling stress and salt stress-induced photoinhibition (Wang et al., 2008). Our data showed that the changing tends of F v/F m and NPQ values in both Col-0 and PeZEP-overexpressing transgenic plants under chilling stress (4°C) were similar to those under HL stress, which aggravated the photoinhibition, whereas there was no significant difference between Col-0 and transgenic plants (Supplementary Fig. S7). All the transgenic plants showed a better growth than the Col-0 plants, and had higher survival rate, higher F v/F m value, lower MDA content, and higher activity of antioxidant enzymes under drought stress than those of Col-0 plants, which was consistent with the results of AtZEP and MsZEP transgenic plants (Park et al., 2008; Zhang et al., 2016).
Previous studies had reported that RD29a, RD29b, and P5CS were identified as stress-responsive genes in the AtZEP-deficient mutant los6/aba1 (Xiong et al., 2002). In addition, both the drought and salt stress could upregulate the expression level of endogenous DREB and P5CS in MsZEP-overexpressing tobacco (Zhang et al., 2016). Our results showed that the expression levels of AtP5CS and AtRD29b were much higher in PeZEP-overexpressing Arabidopsis than in Col-0 plants under the drought stress, indicating that the enhanced drought tolerance in transgenic plants might be caused by affecting expression of endogenous stress-responsive genes. These results suggested that overexpression of PeZEP could confer drought tolerance in transgenic plants.
Moso bamboo is a large woody bamboo with multiple values, which resulted in so much attention having been paid on its research and development. In future study, the cloning and functional analyses of PeZEP1 and PeZEP2 in moso bamboo will be conducted to make a full knowledge on the evolution and functional diversification. Functional characterization of PeZEPs may not only illuminate the molecular mechanisms of PeZEPs involved in ABA biosynthesis, but also in the xanthophyll cycle and carotenoid biosynthesis, which provided global knowledge on the multiple functions of PeZEPs under different physiological conditions.
Footnotes
Acknowledgments
This work received financial support from the National Science Foundation of China (No. 31370588) and the Sub-Project of the National Science and Technology Support Plan of the Twelfth Five-Year in China (No. 2015BAD04B01).
Disclosure Statement
The authors declare no competing financial interests. All of the authors agreed to the submission of this article. Z.G. and Y.L. designed the experiments. Y.L. and H.S. performed experiments. Y.L. and Z.G. analyzed data. L.L. and H.Z. contributed reagents/materials/analysis tools. Z.G. and Y.L. wrote the article. All authors read and approved the final article.
References
Supplementary Material
Please find the following supplemental material available below.
For Open Access articles published under a Creative Commons License, all supplemental material carries the same license as the article it is associated with.
For non-Open Access articles published, all supplemental material carries a non-exclusive license, and permission requests for re-use of supplemental material or any part of supplemental material shall be sent directly to the copyright owner as specified in the copyright notice associated with the article.