
Abstract
Musclin is a bioactive factor that functions in regulating the muscle growth and metabolism. To investigate the transcriptional regulatory mechanism of the gene, the 1.4 kb musclin promoter in sheep was cloned (GenBank accession: JX966391) and the sequence was analyzed to predict the motifs associated with muscle growth. Next the enhanced green fluorescent protein (EGFP) was selected as the reporter gene and various wild-type and motif-mutant vectors were constructed. The transcriptional regulatory activities were compared by observing the fluorescence strength and detecting the EGFP mRNA expression in C2C12 myoblasts transfected with the vectors. The results showed that the different lengths of promoters could drive the transcription of EGFP and the mutation of some motifs up- or downregulated the activity of the promoter. Furthermore, the electrophoresis mobility shift assay showed that these motifs regulated the musclin gene transcription through binding to the corresponding transcriptional factors in sheep muscle tissue.
Introduction
M
The expression level of musclin gene was associated with hormone, nutrition, the differentiation status of myocyte, the myofiber type, and some other factors (Nishizawa et al., 2004; Staiger et al., 2006; Banzet et al., 2007; Gu et al., 2015; Sierra et al., 2016). But the transcriptional regulatory mechanism of musclin gene expression is not clear. Yasui et al. (2007) demonstrated that the forkhead transcription factor FoxO1 downregulating musclin might be because it repressed musclin promoter activity by interacting with Sp1 through Sp1 site along the musclin promoter. However, many other regulatory motifs along the musclin promoter still remain to be identified except Sp1 site.
To explore the transcriptional regulation mechanism of the musclin gene promoter, in this study, the 1.4 kb musclin gene promoter was amplified from the genome of sheep (GenBank accession: JX966391), and many putative regulatory elements of the promoter were analyzed through bioinformatics, and then some important motifs were identified by the mutation test and the electrophoresis mobility shift assay (EMSA).
Materials and Methods
Cloning and sequence analysis of the 5′ regulatory region of sheep musclin gene
The genomic DNA was extracted from the blood of Small Tail Han sheep with DNA extraction kit (Shanghai Sangon Biotech Co.). Based on the 5′ regulatory and untranslated region of bovine musclin gene (NC_007299), the high homologous region was found along the sheep genome sequence (ACIV010109253) by NCBI blast, and then PCR primers of sheep musclin gene promoter were designed as follows: sense primer, 5′-CTATATTGAGAGTTTTCCAT-3′, and antisense primer, 5′-CTAACTCTTTATCGGCTT TT-3′, and the 1.453 kb 5′ regulatory region of sheep musclin gene was amplified by PCR. The PCR was performed in a total volume of 50 μL containing 25 μL of 2× Es TaqMaster Mix (Takara Co.), 2 μL of forward and reverse primers, respectively (10 μM), and 1000 ng of template DNA. The reaction conditions were as follows: an initial denaturation at 95°C for 2 min, followed by 35 cycles of 95°C for 30 s, 58.7°C for 30 s, and 72°C for 1 min, and then a final extension at 72°C for 10 min.
After amplification, the PCR products were cloned to pEASY-T3 vector (Takara Co.) for constructing the recombinant vector T-MUSCLINpro (1.4 kb). Then the DNA fragment was testified by restriction enzyme digestion and DNA sequencing analysis. The regulatory motifs of sheep musclin gene promoter were further analyzed by the Genomatix MatInspector software (the parameter: Core sim. = 1, Matrix sim. >0.9).
Construction of MUSCLINproW-EGFP and MUSCLINproXM-EGFP vectors
Six musclin promoter fragments with different lengths (1.4, 1.2, 1.0, 0.8, 0.4, and 0.3 kb) were generated from T-MUSCLINpro (1.4 kb) by PCR with similar reaction system and condition as the above except for the primers (Supplementary Table S1; Supplementary Data are available online at
Site-directed mutations of some motifs, including E-box (E1–E8), GA-box, myocyte enhancer factor (MEF)2, FOXO, SMAD3, muscle-specific Mt-binding site (MTBF), MTATA, and MEF3, were introduced by PCR, and the MUSCLINPro XM -EGFP vectors containing different mutated motifs were constructed as before (Du et al., 2007). The involved schematic diagram and PCR primers are shown in Supplementary Figure S1 and Supplementary Table S2. In brief, the motif sequence was replaced with a suitable digestion site and the two fragments that flanked the targeted motif were amplified, respectively, and inserted successively into the corresponding digest sites of the reporter vector pEGFP-N1 (4.1 kb).
Cell culture, plasmid transfection, and fluorescence observation
C2C12 myoblasts were routinely cultured, and the negative control vector pEGFP-N1 (4.1 kb) and various test vectors MUSCLINpro W/XM -EGFP were transfected into C2C12 myoblasts as before (Du et al., 2007). After transfection, the cells were cultured for a further 24 h till the fluorescence observation and the fluorescence quantitative PCR were done. Fluorescence was observed by the fluorescent microscope (Leica Co.). Each experiment was repeated six times.
Fluorescence quantitative PCR and statistical analysis
The transfected cells were collected and then the total RNA was extracted using TRIzol reagent and was converted to cDNA using reverse transcription reagent according to the manufacturer's instructions (Takara Co.). The fluorescence quantitative PCR for EGFP was carried out using Prime Script™ RT Master Mix Kit according to the manufacturer's instructions (Takara Co.). Glyceraldehyde-3-phosphate dehydrogenase (GAPDH) was used as the endogenous control. The primers were designed as follows: reference gene (GAPDH, 171 bp): F 5′-ACCCAGAAGACTGTGGATGG-3′, R 5′-CACATTGGGGGTAGGAACAC-3′; target gene (EGFP, 187 bp): F 5′-ACGTAAACGGCCACAAGTTC-3′, R 5′-AAGTCGTGCTGCTTCATGTG-3′. The relative expression quantities of EGFP mRNA were calculated by 2−ΔΔCt analysis, and then SPSS and Excel software were used for data analysis. p < 0.01 and p < 0.05 were taken as the level of statistical significance.
Electrophoretic mobility shift assay
The nucleoprotein extracts from sheep muscle tissue were prepared using the Nuclear and Cytoplasmic Protein Extraction Kit (Beyotime Co.), and the wild-type and mutational probes for motifs were designed (Supplementary Table S3). The binding reactions of DNA probes and nucleoproteins were performed in EMSA-binding buffer with cooperation of Jiangsu KeyGEN BioTECH Co. For the negative control, the free-labeled probe was used. To testify the specificity of binding, the labeled mutational probe was used for the control reaction and the 1000-fold molar excess of unlabeled probe was used for the competition reaction. Then the DNA/protein complexes were loaded onto the 6.5% polyacrylamide gel and the gel was run at 150 V for 40 min. After that, the gel was transferred onto binding membrane, then the biotin-labeled probe was detected by chemiluminescence, and the G BOX chemiXR5 gels imaging system was used for imaging.
Results
Cloning of the 1.453 kb 5′ regulatory region of sheep musclin gene
The 1.453 kb musclin gene fragment in sheep, containing promoter and 5′UTR, was amplified, cloned, and sequenced (Fig. 1A). Compared with the sequence of sheep genome (ACIV010109253), the sequence cloned in this study only lacked one base (JX966391), which was attributed to polymorphism.

Amplification and activity detection of the 1.4kb sheep musclin promoter.
Analysis of the regulatory motifs along the musclin gene promoter
The bioinformatic analysis by the MatInspector Program revealed that there were many different putative gene-specific nuclear factor-binding sites, including some muscle-specific transcription factor-binding sites, along the promoter region of the sheep musclin gene (Supplementary Fig. S2 and Supplementary Table S4).
Transcriptional regulation activity of the sheep musclin promoter with different lengths in C2C12 myoblasts
The 5′ deletion analysis showed that different lengths of sheep musclin gene promoter drove the EGFP gene transcription in C2C12 cells (1.4 kb > 0.8 kb > 1.2 kb > 1.0 kb > 0.4 kb > 0.3 kb) (Figs. 1B, C and 2). What is interesting was that the activity of 0.8 kb promoter was higher than that of 1.0 or 1.2 kb promoter (p < 0.01, p < 0.05). This suggested that there were negative regulatory motifs along the sheep musclin promoter region from 0.8 to 1.0 kb. Furthermore, we predicted 24 putative motifs along the region from 0.8 to 1.0 kb by using MatInspector bioinformatics tool (Supplementary Table S5).
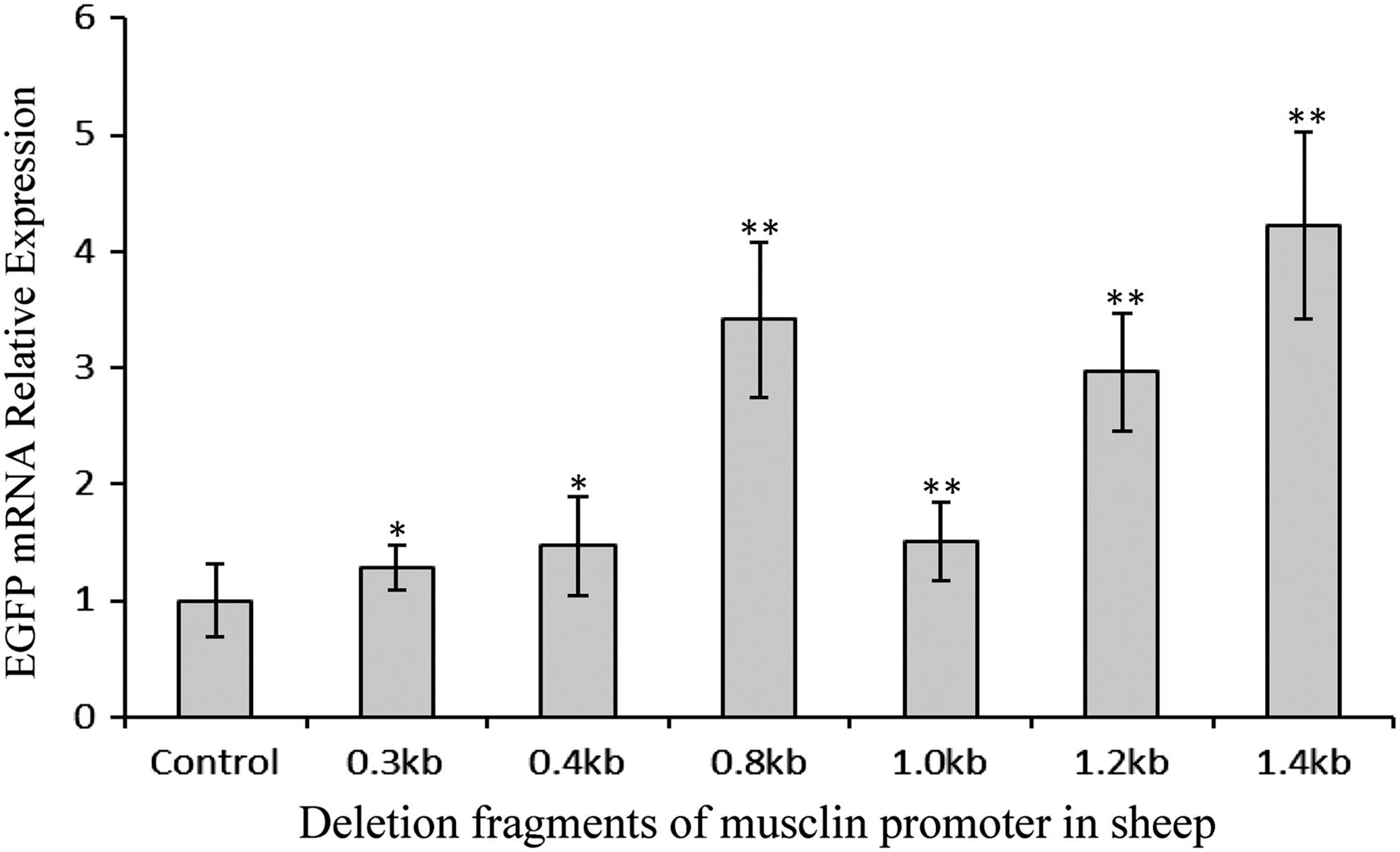
EGFP mRNA expression in C2C12 cells transfected with different lengths of sheep musclin promoter. Bars indicate means ± standard deviations for six replicates. * or ** indicates that there were significant differences compared with the control vector, p < 0.05 or p < 0.01.
Effects of motif mutations on the transcriptional regulation activity of the sheep musclin gene promoter in C2C12 cells
The results of fluorescent quantitative PCR showed that the expression levels of EGFP mRNA in C2C12 transfected with the motifs-mutant sheep musclin promoters were changed compared with the wild-type sheep musclin promoter (Fig. 3). The mutation of E1, E3, E4, E5, E6, E7 (enhancer box, E-box) or GA-box reduced the expression of EGFP mRNA (p < 0.01) by 57.93%, 35.87%, 41.64%, 28.94%, 66.18%, 56.00%, or 73.57%, respectively; whereas the mutation of E2, E8, MEF2-1, MEF2-3, FOXO (forkhead member of the class O), MTATA (muscle TATA box), SMAD3 (Smad-binding element), MTBF, or MEF3 enhanced the expression of EGFP mRNA (p < 0.01) by 11.13%, 84.91%, 228.28%, 87.64%, 253.73%, 252.54%, 366.00%, 300.57%, or 229.89%, respectively. These data suggested that these motifs played important positive or negative regulatory roles in the transcription of the musclin gene in sheep.
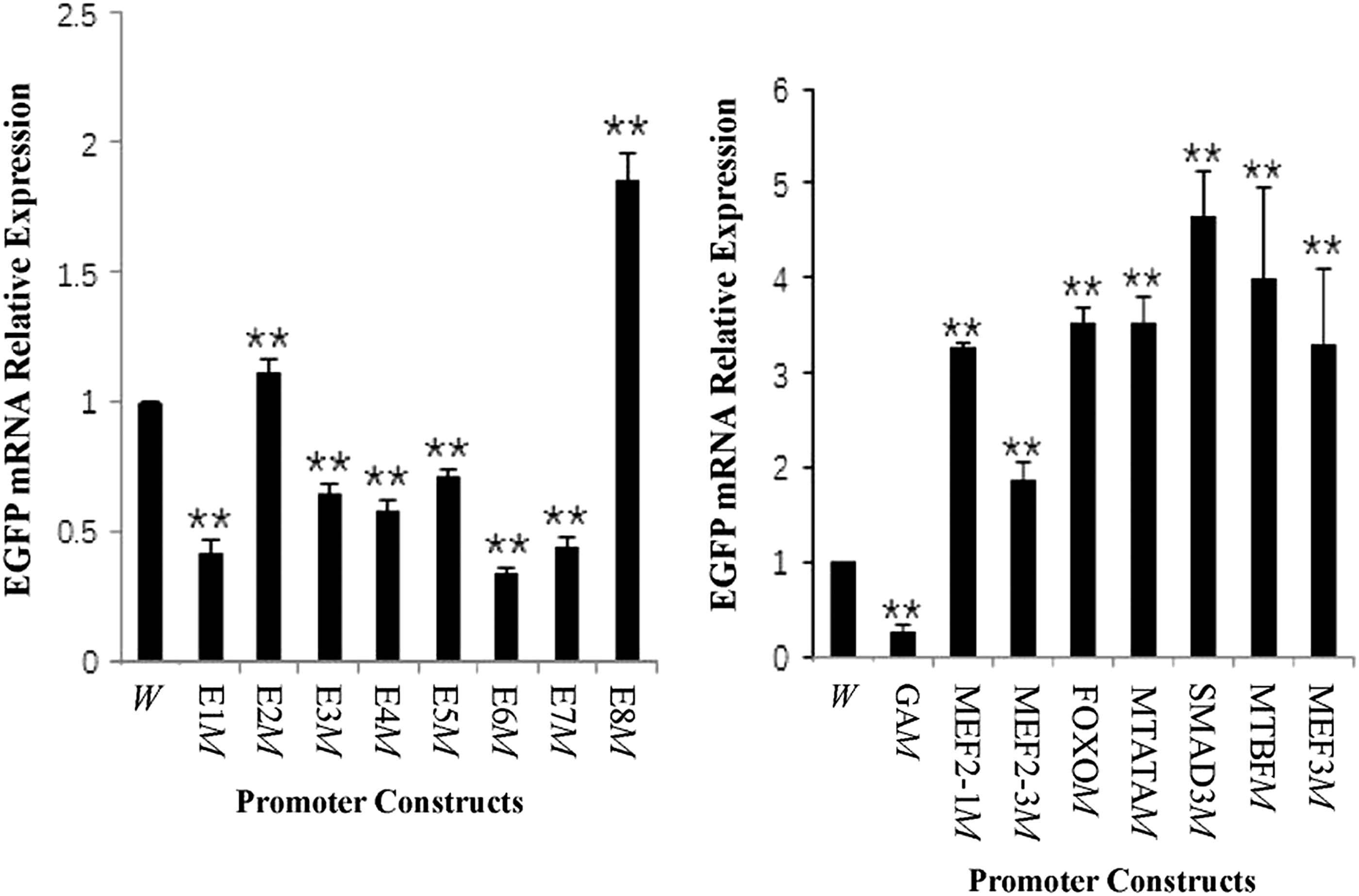
EGFP mRNA expression in C2C12 cells transfected with sheep musclin promoters containing different mutant (M) motifs. **Indicates that there were significant differences compared with the wild-type (W) promoter, p < 0.01. E, enhancer box (E-box); FOXO3, forkhead member of the class O; GA, GA-box; MEF2, myocyte enhancer factor-2; MEF3, myocyte enhancer factor 3; MTATA, muscle TATA box; MTBF, muscle-specific Mt binding site; SMAD3, Smad-binding element.
Binding of nucleoproteins to several regulatory motifs
The results of EMSA showed that some nucleoproteins, which could specifically combine with the motifs of GA-box, E1, E2, E5, E8, MEF2, FOXO3, MTBF, and SMAD3, existed in sheep gastrocnemius (Fig. 4). These results suggested that these motifs functioned by the mechanism of binding with the corresponding transcription factors.
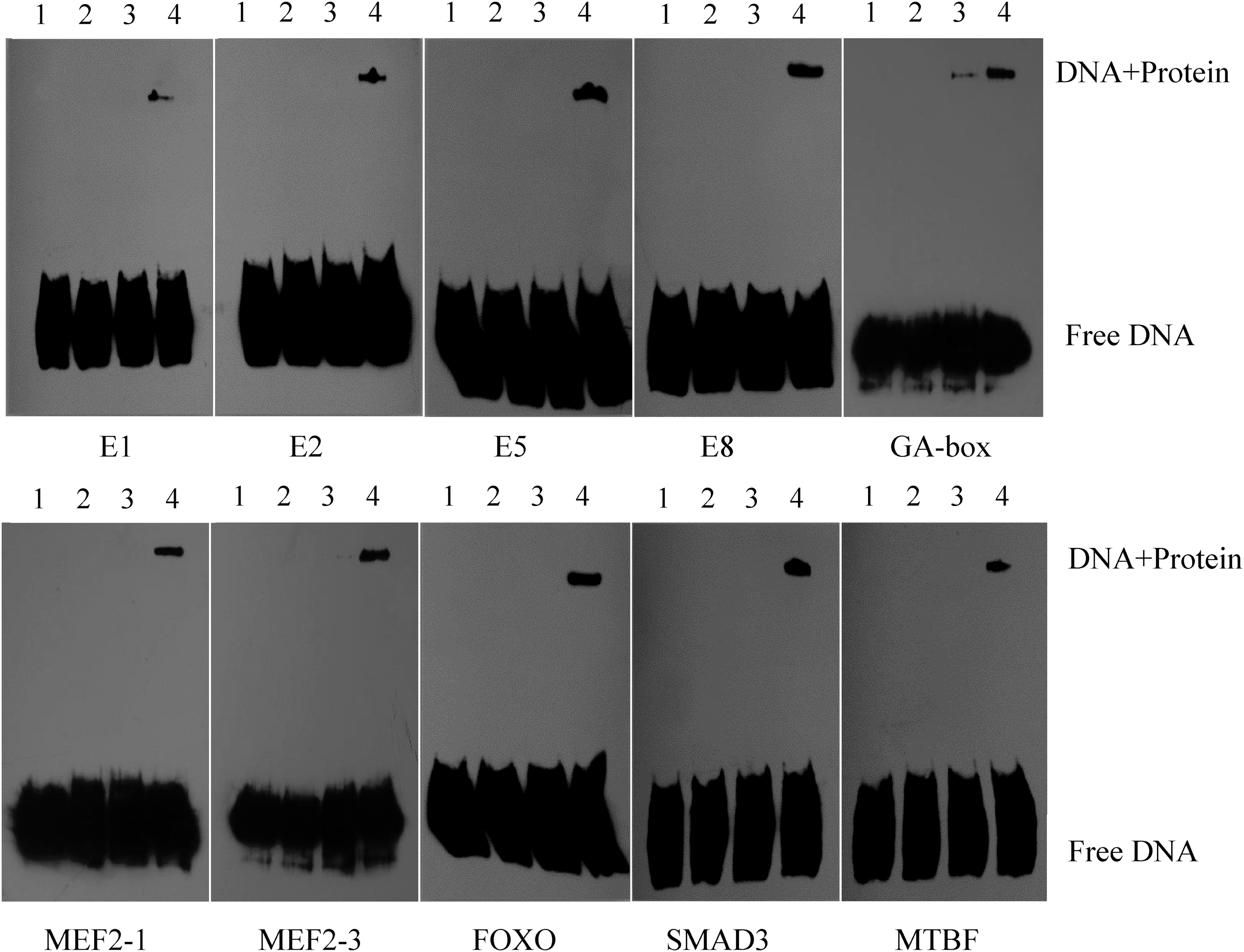
The EMSA results of the interreaction between some motifs along the sheep musclin promoter and the nuclear proteins in sheep muscle. 1. Free labeled probe; 2. Labeled mutational probe + nucleoprotein; 3. Labeled probe + unlabeled probe + nucleoprotein; 4. Labeled probe + nucleoprotein.
Discussion
Role of E-box in the musclin gene transcription
E-box exists in the promoter region of many muscle-related genes such as myostatin and myomaker and played important roles in regulating the transcriptions of these genes (Du et al., 2007; Millay et al., 2014). The core sequence of canonical E-box is CANNTG, and it can combine with the basic helix-loop-helix nuclear proteins such as E protein (including E12 and E47) and MRFs (including MyoD, myogenin, Myf5, and MRF4) (Jeong et al., 2014; Millay et al., 2014; Lee et al., 2015). In this study, eight E-boxes played different positive or negative roles in regulating the activity of the sheep musclin promoter, which might be due to the different central dinucleotide and flanking sequence of E-boxes. Some previous researches showed that E-boxes that had a G/C-rich central dinucleotide were associated with genes expressed in myotube during differentiation, whereas E-boxes that had an A/T-rich central dinucleotide were associated with genes expressed in myoblasts (Soleimani et al., 2012; Moncaut et al., 2013). Snail proteins (transcriptional repressors) tended to bind to E-boxes that had a G/C-rich central dinucleotide and thus Snail must be removed from E-box for the binding of MyoD when differentiation was coming up (Soleimani et al., 2012; Moncaut et al., 2013). In our study, the central dinucleotide of E2 and E8 was one G/C plus one A/T and it looked there was some transcriptional repressor protein like Snail binding to E2 or E8 in the myoblasts. But E3 and E4 that had two central G/C base pairs were positive motifs in this study, which suggested that the different flanking nucleotides influenced the functions of E-boxes. A previous research showed that the transcriptional regulatory function of E-box along the troponin I enhancer was changed when the troponin I E-box was substituted with other muscle-related gene E-boxes and the changes were associated with not only the central dinucleotides but also the flanking nucleotides of E-boxes (Yutzey and Konieczny, 1992). The different EMSA results for the E-boxes suggested that the E-boxes regulated the transcription of musclin gene by binding with the corresponding transcription factors or other mechanisms such as interacting with the adjacent motifs.
Role of GAGA-box in the musclin gene transcription
GAGA factor was combined with the sequence of rich GA base (GAGA-box) to regulate the transcription of downstream genes in drosophila through recruiting the relevant factors such as RNA polymerase II (Fuda et al., 2015). In mammals, the relevant researches are fewer. Some study showed that there also existed GAGA motifs in human growth hormone receptor gene (Kenth et al., 2011). In the study, we also found and identified some adjacent GAGA motifs in sheep musclin gene promoter, which played a positive regulatory role in the transcription of musclin gene by combining with nucleoproteins.
Role of MEF2 motif in the musclin gene transcription
There are binding sites for MEF2 factors in the promoter region of many genes that can regulate skeletal muscle growth and metabolism such as myostatin (Du et al., 2007) and GLUT4 (Goyaram et al., 2014), which suggests that it plays an important role in regulating the muscle genesis and development. In this study, we identified two MEF2 motifs through the mutation analysis, which played negative regulatory roles in the activity of the sheep musclin promoter and proved that MEF2 motifs functioned by binding with nucleoproteins. The nucleoproteins might be one or several subtypes (MEF2A, MEF2B, MEF2C, and MEF2D) of the MEF2 family.
Role of FOXO motif in the musclin gene transcription
As a subtribe of forkhead family, FoxOs such as FoxO1 (FKHR), FoxO3 (FKHRL1), and FoxO4 had single and/or combinational functions in inducing skeletal muscle atrophy (Fjällström et al., 2014; Jaitovich et al., 2015; Milan et al., 2015) and there was a correlation between FoxO1a/3a binding and inhibition of basal glucose-6-phosphatase catalytic subunit (G6Pase) gene transcription by insulin (Onuma et al., 2006). Yasui et al. (2007) showed that FoxO1 repressed the musclin promoter activity in mouse and human, but they did not find FKHR motif directly bound to FoxO1. In the study, FOXO (FKHRL1) was identified to be an important motif along the sheep musclin gene promoter and there existed nucleoprotein in sheep muscle that could combine with FOXO motif. It suggested that the corresponding transcriptional factor FoxO3 of FKHRL1 motif might have a negative impact on the transcriptional regulatory activity of the musclin gene. Musclin can decrease the role of insulin and inhibit the uptake of glucose (Nishizawa et al., 2004; Liu et al., 2008). Taken together, it is possible that for balancing the organism's function, FoxO promotes the glucose uptake function of insulin partly by inhibiting the musclin gene expression when FoxO participates in muscle atrophy.
Role of SMAD motif in the musclin gene transcription
In the nucleus, the combinations of Smad proteins with target genes have selectivity, and the transcriptional regulation of target genes by Smad proteins requires the interaction of Smad proteins with other transcription factors (Inman et al., 2002). In the study, we found that SMAD3 was a negative regulatory motif of the sheep musclin promoter and there existed the specific proteins in sheep muscle, which could combine with the SMAD3 motif. Smad3 could repress MEF2-dependent gene transcription in myogenic differentiation (Liu et al., 2004), and the recent studies showed that Smad3 augmented the FoxO3-induced expressions of MuRF-1 and Atrogin-1, which were required during muscle atrophy (Bollinger et al., 2014), and myostatin induced insulin resistance by degrading IRS1 protein through the E3 ligase in an Smad3-dependent manner (Bonala et al., 2014). Thus, it was possible that Smad3 inhibited the sheep musclin gene expression by binding to SMAD3 motif at the same time that Smad3 promoted the muscle atrophy and induced insulin resistance, which aimed at coordinating the functional balance of organism.
Role of MTBF motif in the musclin gene transcription
MTBF was found along the sheep myostatin promoter in our previous study (Du et al., 2007). The expression of reporter gene was enhanced when the MTBF motif was mutated in the study, which suggested that MTBF was a negative regulatory motif for the activity of musclin gene promoter. The EMSA results showed that there existed nucleoproteins in sheep muscle tissue that could combine with MTBF motif. The studies suggested that the binding of MTBF motif and the corresponding factors was important for regulating the transcription of the muscle-related genes.
Conclusion
In conclusion, in this study we identified some important motifs along the sheep musclin promoter by the motif mutation and EMSA. These motifs regulated the musclin gene transcription through binding to single or complex transcriptional factors. In addition, the interaction among various motifs might be present. It is necessary to identify the exact transcription factors targeted at the motifs and investigate the interaction of various motifs in a further study.
Footnotes
Acknowledgments
This work is supported by the National Natural Science Foundation of China (Nos. 31272527 and 31140074), the “131” Leading Talents Project of Shanxi Higher Education Institution, the Program for Young and Middle-aged Academic Leaders of Shanxi Agricultural University (No. XD201206), and the Fund for Shanxi Key Subjects Construction (FSKSC).
Disclosure Statement
No competing financial interests exist.
References
Supplementary Material
Please find the following supplemental material available below.
For Open Access articles published under a Creative Commons License, all supplemental material carries the same license as the article it is associated with.
For non-Open Access articles published, all supplemental material carries a non-exclusive license, and permission requests for re-use of supplemental material or any part of supplemental material shall be sent directly to the copyright owner as specified in the copyright notice associated with the article.