
Abstract
DNA and Cell Biology (DNA&CB) is officially retracting the paper by Zhao H, Wang Y, Liu J, Shao Y, Li J, Chai H, Xing M, entitled, “Molecular Characterization and Biological activity of Interferon-α in Indian Peafowl (Pavo cristatus),” [Epub ahead of print]; 2017, DOI: 10.1089/dna.2017.3798. The Editor-in-Chief of DNA&CB, Dr. Carol Shoshkes Reiss, was alerted to a discrepancy between the findings in the article by Zhao et al., and those of others, about the absence of expression of ISG15 in chickens. Dr. Reiss requested from the authors a clarification in their observations and inquired about the failure to include relevant citations in the reference section of the paper. Based on the response from the authors, it appeared that they did not have the confidence in the data as they were not able to repeat the experiments, and were also unsure of the molecular probes that were used in the study. Therefore, the Editor has determined that the paper should be officially retracted from DNA and Cell Biology.
Introduction
I
Type I IFNs have multifunctional activities, different induction of ISGs resulting in overlapping but distinct properties. For example, both human IFN-α and IFN-β constitute a primary antiviral mechanism in many cell types, the former is more potent in antiviral activity, and the latter is more active in antiproliferation (Ivashkiv and Donlin, 2014). Till now, human recombinant IFN-αs have been elaborated as part of the innate immune response, suppress influenza A virus replication, and reduce disease in ferrets, mice, and nonhuman primates (Tumpey et al., 2007; Kugel et al., 2009; Matzinger et al., 2011). Also, porcine IFN-α controls swinish viral diseases by stimulating the immune response, after inhibiting replication of swine fever virus (Xia et al., 2005), foot-and-mouth disease virus (Dias et al., 2011), and porcine parvovirus (Liu et al., 2011). However, there has not been much progress in the expression and characteristics analysis of Pavo cristatus IFN-α (PcIFN-α). With the development of the peacock breeding industry and the spread of various avian diseases, further studies on PcIFN-α are required.
The Indian peafowl (P. cristatus), a rare ornamental animal species, is a group of small, relatively somber forest pheasants of tropical Asia (Madge, 2002). Recently, it has invoked great inclination because of their tail feathers of train, courtship display, and keratin homogeneity in the tail feathers (Pabisch et al., 2010). However, this species has been plagued by many diseases. Newcastle disease virus (NDV), an acute infectious disease, causes major losses and devastation to poultry industry covering 200 species of birds (Alexander, 1996). Lee et al. (2004) and Dou and Yang. (2007) have isolated NDV from peafowls in Korea and China. Munir et al. (2012) have investigated a disease outbreak involving high mortality of peafowls in Pakistan with successful isolation of NDV. In addition, herpesvirus, pox viruses, avian mycobacteriosis, and Eimeria pavonina have entangled peafowl permanently (Al et al., 1979; Hauck and Hafez, 2012; Seimon et al., 2012; Stepien-Pysniak et al., 2016). What is worse, in a clinical case of Chlamydophila psittaci infection on peacock, severe pneumonia, airsacculitis, and diarrhea were observed with 60% of morbidity and 10% of mortality, even six farmers osculatory encountered severe acute respiratory syndrome characterized by pneumonia, high fever, and respiratory distress (Yang et al., 2011). Therefore, there is an urgent need to develop nontoxic and effective antiviral drugs to control viral infections in Indian peafowl.
Materials and Methods
Cell and viruses
The erythrocyte of Indian peafowl was collected from the Livestock Research Institute of Academy of Agricultural Sciences in Harbin, Heilongjiang province. Vesicular stomatitis virus (VSV) were kindly provided by Dr. Qu Liandong, NDV and AIV were kindly provided by Dr. Wang Xiurong, both from Harbin Veterinary Research Institute, Chinese Academy of Agricultural Sciences, specific pathogen-free (SPF) eggs were also purchased here. Chicken Gallus gallus IFN-α (ChIFN-α) was preserved in our laboratory, whose mature protein has been cloned and expressed in E. coli. Two New Zealand White Rabbits (female, 4 weeks old) were purchased from the Laboratory Animal Center of Harbin Veterinary Research Institute. Animal experiment protocols were reviewed and approved by the Animal Care, Use, and Ethics Committee at Northeast Forestry University (approval No. UT-31; June, 20 2014).
Cloning of the gene encoding PcIFN-α and analysis of sequences
Pairs of specific primers for PcIFN-α (Table 1) were designed according to the conserved region of ChIFN-α sequence from GenBank (GU119896); genomic DNA extracted from the erythrocyte of Indian peafowl was applied as template. PCR products were cloned into the pMD19-T Vectors (Takara, Japan) and sequenced (BOSHI, Harbin, China). Characterizations were analyzed by bioinformatics software. Sequences analysis was performed on the DNAStar program (DNASTAR, Inc.). The obtained sequences were identified by BLAST (
Forward primer (mPcIFN-A) containing an EcoRI site (underlined), and reverse primer (mPcIFN-B) containing a HindIII site (underlined).
IFN, interferon; ISGs, IFN-stimulated genes; PcIFN-α, Pavo cristatus IFN-α.
Prokaryotic expression of PcIFN-α
Prokaryotic expression in vitro constructs for PcIFN-α was generated in the vector pET32a (Novagen, Darmstadt, Germany), which included an N-terminal detection and purification epitope (His·Tag). PCR products were cut with EcoR I and Hind III restriction sites, then ligated into a pET32a Vector and transformed into E.coli/BL21. The recombinant PcIFN-α protein was expressed and purified as previously described (Tian et al., 2014). After denaturation and renaturation, we obtained the soluble homogeneous protein with biological activities and stored at −80°C for further use. The final protein concentration was determined using a BCA Protein Assay Kit (Pierce). Both PcIFN-α and ChIFN-α were diluted with PBS to the same concentration.
Production of rabbit antiserum against recombinant PcIFN-α
Rabbits were immunized subcutaneously with 0.1 mg of purified PcIFN-α protein emulsified with 50% Freund's complete adjuvant (Sigma). Afterward, the rabbits were boosted with 0.1 mg purified PcIFN-α protein in 50% Freund's incomplete adjuvant (Sigma) every 2 weeks for three times. Two weeks after the last injection, blood was collected from the rabbits' hearts, antiserum against PcIFN-α (pAb) was separated at 1000 g and 4°C for 10 min and frozen at −20°C.
Western blot analysis of PcIFN-α
After sodium dodecyl sulfate polyacrylamide gel electrophoresis (SDS-PAGE), proteins were transferred to the polyvinylidene fluoride membrane from gel without staining, 6-poly histidine monoclonal antibody (Zen BioScience, China) was used as the primary antibodies (1:500), and specific reaction products were detected with horseradish peroxidase (HRP)-conjugated goat antirabbit IgG. The conditions for Western blot have been described previously (Zhao et al., 2017).
Antiviral activity and antibody blocking assay in vitro
VSV, NDV, and AIV titers were determined using an endpoint dilution assay, and the titers were expressed as the tissue culture infectious dose 50 (TCID50) per milliliter using the Reed-Muench method (Pizzi, 1950). The antivirus activities of PcIFN-α and ChIFN-α were determined and compared by cytopathic effect inhibition assays on chicken embryo fibroblasts (CEFs). CEFs were freshly prepared from 9 to 11-day-old-embryonated SPF eggs. The cells were grown in Dulbecco's minimal essential medium (HyClone, UT) supplemented with 10% fetal bovine serum (HyClone), 0.22% NaHCO3, and 100 U/mL penicillin and streptomycin at 37°C in a humidified 5% CO2 incubator. In brief, monolayers CEFs were seeded in a 96-well plate (100 μL/well) for 24 h. Then, the cells' supernatant was replaced with 100 μL of fresh medium containing serial dilutions of PcIFN-α or ChIFN-α (0.5 mg/mL). After 24 h incubation, an additional 100 μL of medium containing VSV, NDV, and AIV (100 TCID50/well) was added. Simultaneously, positive control (without IFN-α, only virus) and negative control (only IFN-α, without virus) were set. After observing the results with inverted microscope for 12–72 h, the dilution of
To check whether this antiviral activity was specific, the neutralization antiviral activity of PcIFN-α was measured according to the method as previously described (Shao et al., 2015) with some modifications. CEFs were treated with PcIFN-α (100 μL) neutralized by rabbit anti-PcIFN-α antibody (diluted 1:20) for 4 h at 37°C. Then, all cells were challenged with 100TCID50 of VSV. The preimmune rabbit serum was used as negative control.
Detection of primary physicochemical characteristics of PcIFN-α
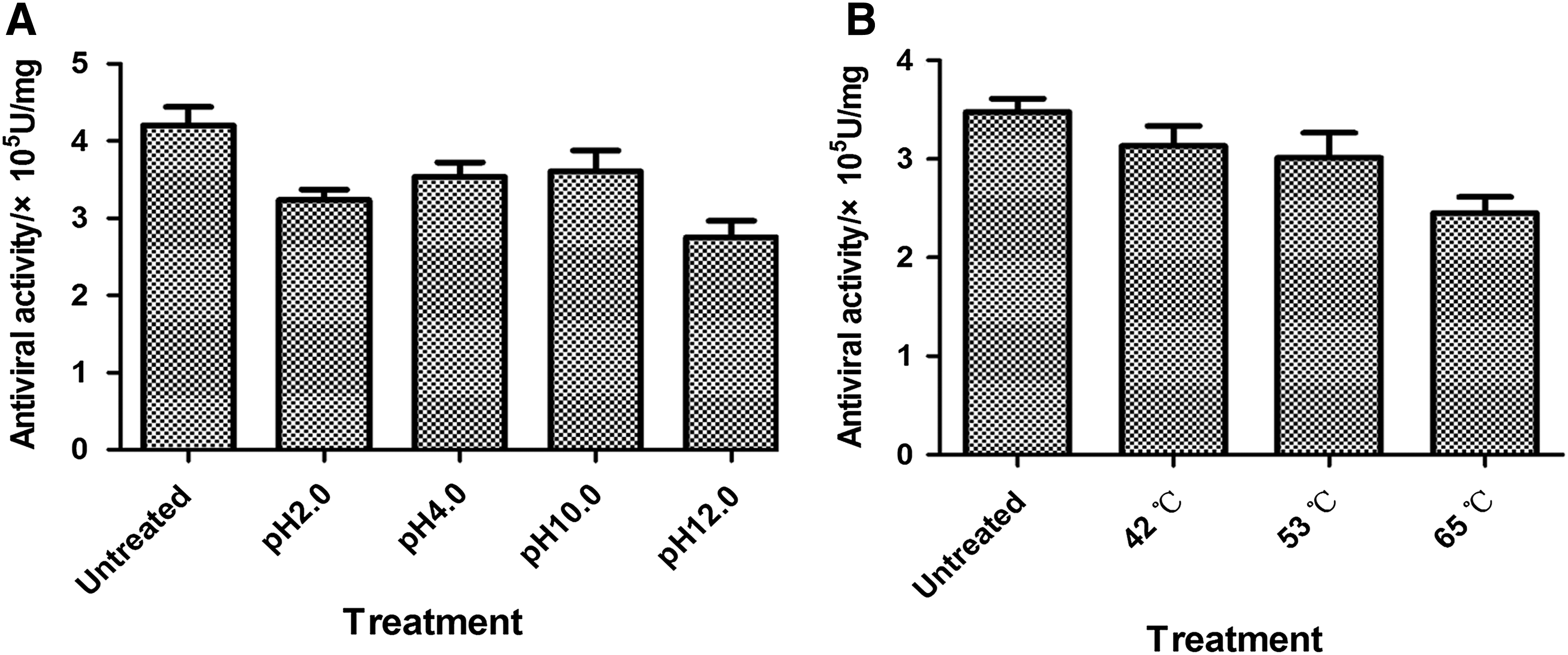
The physicochemical characteristics of PcIFN-α were analyzed using the CEFs/VSV system, and these characterizations included trypsin, pH, and temperature sensitivity as previously described (Shao et al., 2015). Detailedly, the PcIFN-α samples underwent the following processing: (1) combined with 1% trypsin to a final concentration of 0.25% trypsin and placed in a water bath for 1 h at 37°C, (2) combined with hydrogen chloride or sodium hydroxide to adjust pH levels of 2.0, 4.0, 10.0, and 12.0 for 24 h at 4°C, after which they were adjusted back to original pH 7.0, (3) placed in a 42°C, 53°C, and 65°C water bath for 4 h, then they were rapidly placed in an icebox for cooling, after which the antiviral activity was determined by the CEFs/VSV system. The antiviral activities of the treated and untreated samples were compared.
PcIFN-α induced the transcription of Mx1, ISG15, and ISG56
CEFs were seeded in six-well plates and treated with PcIFN-α for 0, 3, 6, 9, 12, and 24 h. Subsequently, the cells' total RNA was prepared using RNAiso Plus reagent (Takara), and then reverse transcribed into cDNA using a PrimeScript RT Reagent Kit (Takara). Quantitative real-time PCR (qRT-PCR) was carried out on a LightCycler® 480 (Roche, Switzerland) and determined with the FastStart Universal SYBR Green Master reagents (Roche). The PCR was set up under the following thermal cycling conditions: the qRT-PCR was performed with the following primers: 95°C for 10 min, 40 cycles of 95°C for 15 s, 60°C for 30 s, and 72°C for 20 s. PcGAPDH-A and PcGAPDH-B, PcMx-1-A and PcMx-1-B, PcISG15-A and PcISG15-B, and PcISG56-A and PcISG56-B (Table 1). GAPDH was used as the internal reference, the transcription of Mx1, ISG15, and ISG56 mRNA was analyzed with the method 2−ΔΔCT (Livak and Schmittgen, 2001).
PcIFN-α induced the protein expression of Mx1
CEFs were seeded in six-well plates and treated with PcIFN-α for 0, 3, 6, 9, 12, and 24 h. Subsequently, cells were lysed in radio immunoprecipitation assay lysis buffer (Beyotime, Beijing, China). Lysates were collected and separated by SDS-PAGE; PAb against Mx-1 (1:5000 dilution) (Proteintech, Wuhan, China) and GAPDH (1:8000 dilution) were used as the primary antibodies, and specific reaction products were detected with HRP-conjugated goat antirabbit IgG.
Statistical analyses
Differences between the mean values of data were assessed with a Student's t-test using SPSS (SPSS, Inc., Chicago, IL). p-Values of 0.05 or less were considered to be statistically significant. All values are expressed as the means ± standard deviations.
Results
Characterization of PcIFN-α gene
The product of recombinant PcIFN-α cDNA was 582 bp DNA sequences, which encoded signal peptide of the N-terminal 31 amino acids and a mature protein of 162 amino acids (Fig. 1A). The open reading frame of the PcIFN-α sequence has been submitted to the GenBank databases with the accession number KJ001188.1.
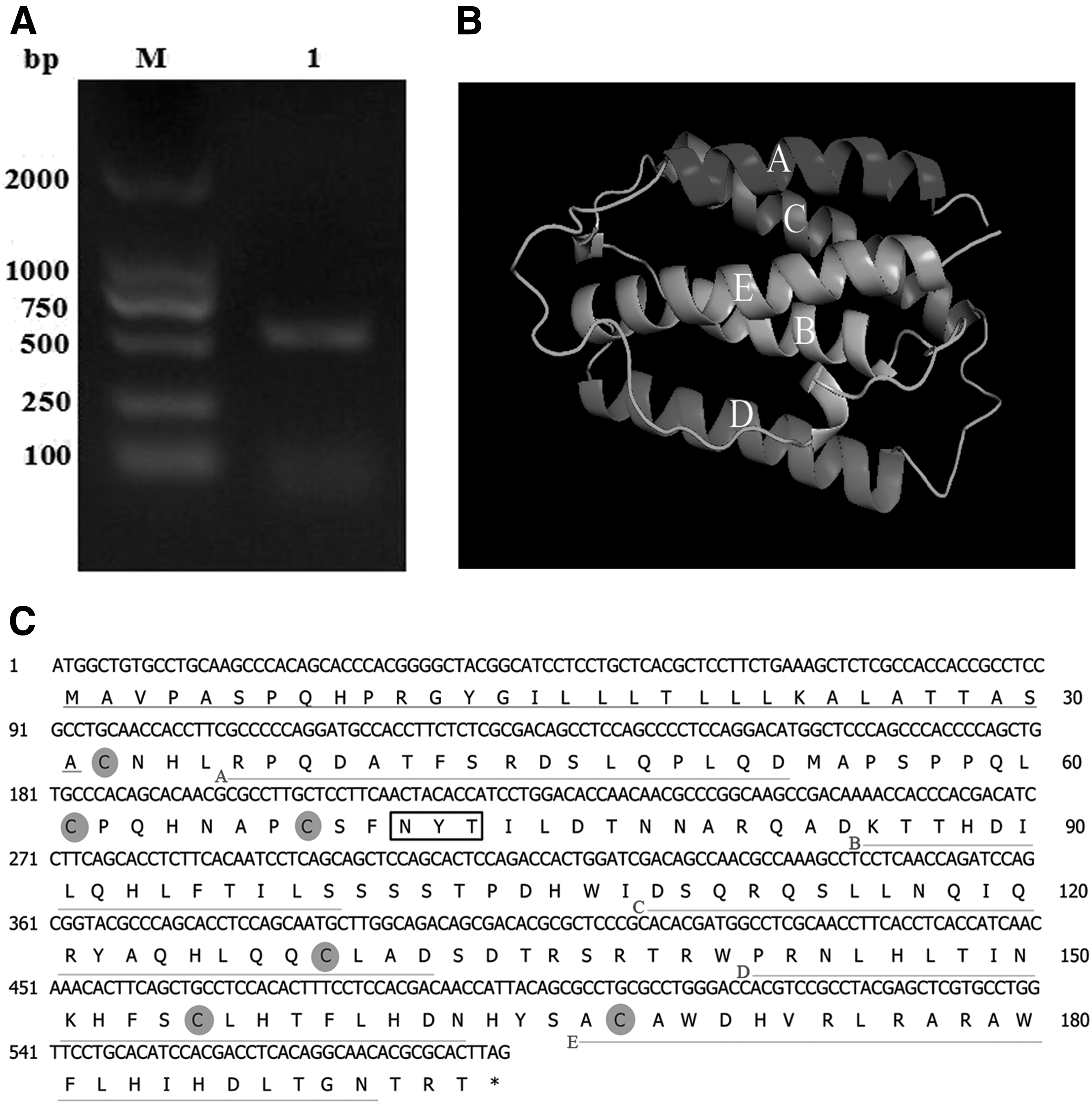
Sequence cloning, analysis, and 3D prediction of PcIFN-α.
SignalP 4.1 server software analysis predicted that the N-terminal 31 amino acids represented the signal peptide (Fig. 1B). Three-dimensional structure prediction website revealed that PcIFN-α contained five putative alpha helices, which was consistent with the IFN molecules analyzed by NMR spectroscopy (Ghasriani et al., 2013) (Fig. 1C). Most secreted proteins in eukaryotes are modified on the amino acid Asn located in the consensus sequence Asn.Xaa.Ser/Thr (NXS/T) by an N-glycan, a process known as N-glycosylation (Song et al., 2013). Analysis using NetNGlyc Server 1.0 predicted that PcIFN-α contained one glycosylation site at 71NYT73 (Fig. 1B). Further analysis demonstrated that PcIFN-α contained six cysteine residues at the positions of 32, 61, 68, 100, 129, 155, and 168. These data demonstrated the existence of a single nucleotide polymorphism (SNP) in PcIFN-α, which was in agreement with previous reports on SNPs of IFN-α in other species (Song et al., 2013). Nucleotide evolution tree analysis showed significant differences in the nucleotide sequences among the PcIFN-α and those of other species of animals and human IFN-αs (Fig. 2A). PcIFN-α clustered with
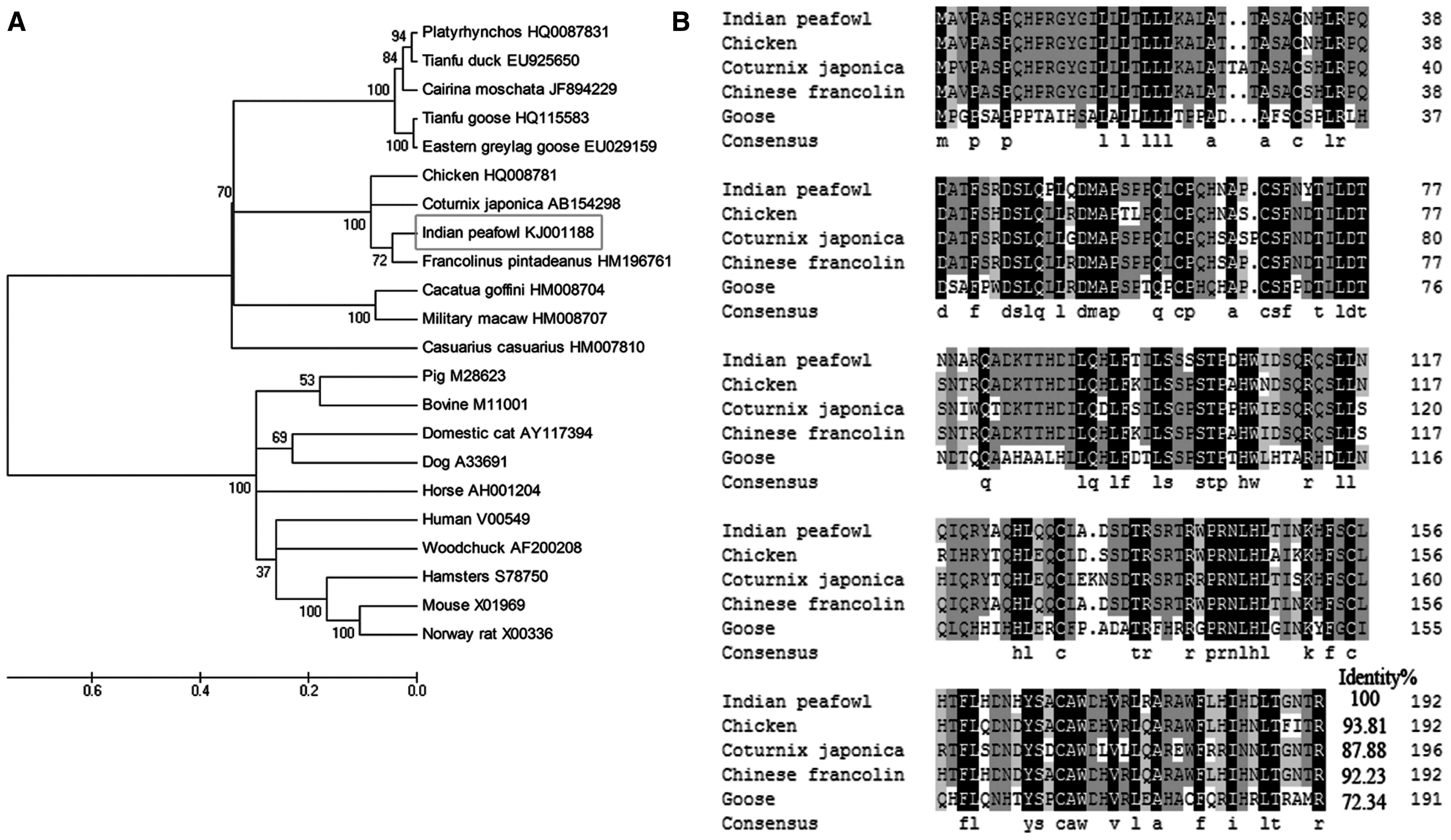
Phylogenetic tree construction of the PcIFN-α and IFN-α amino acid alignment
Protein expression and verification of recombinant PcIFN-α
The gene fragment encoding PcIFN-α protein (without signal peptide) was cloned into the pET32a expression vector. SDS-PAGE analysis revealed that the recombinant protein PcIFN-α was mainly expressed as the inclusion body form with an apparent molecular weight of 38 kDa after induction with 1 mM isopropyl β-
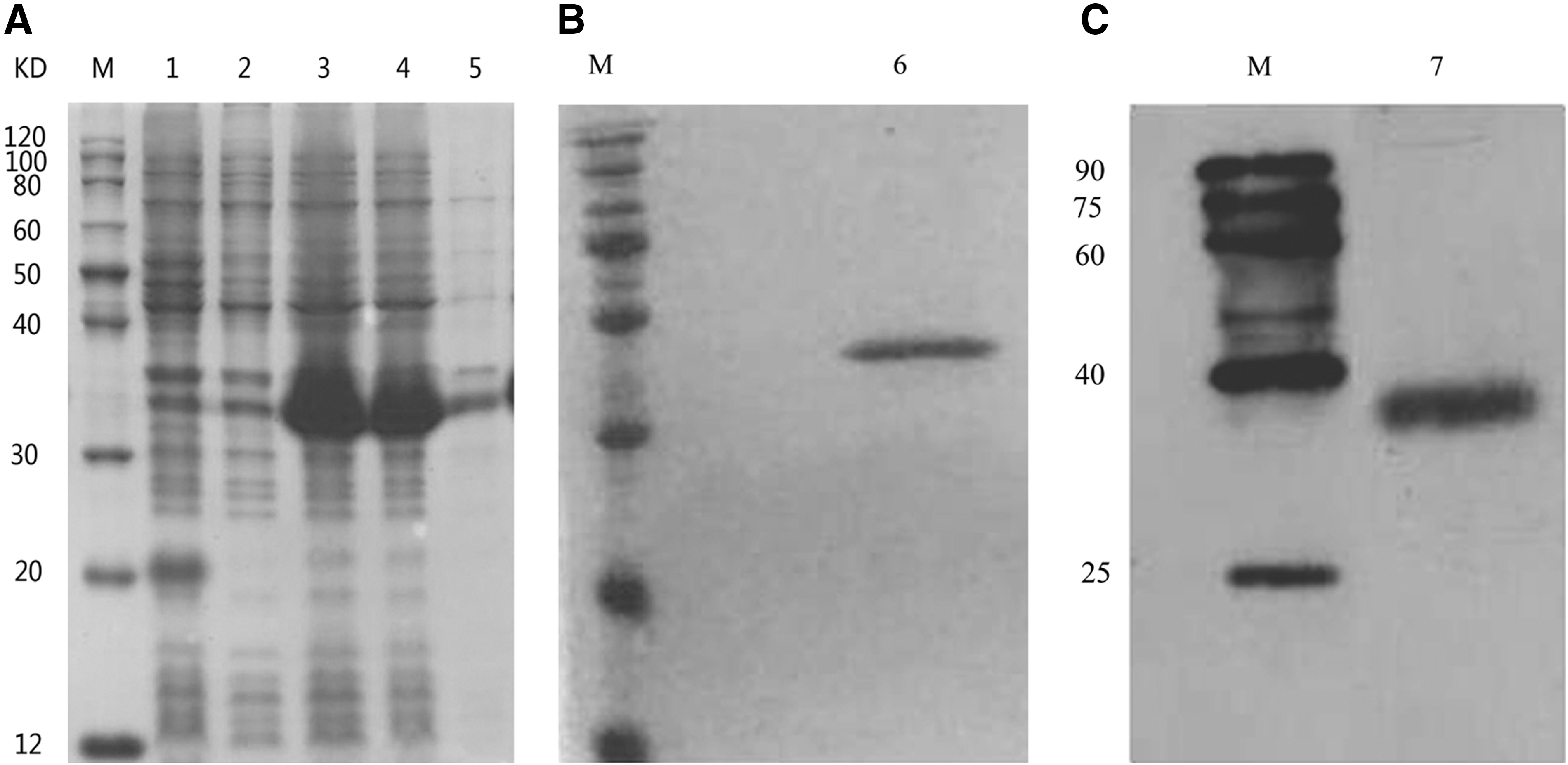
Protein expression and verification of recombinant PcIFN-α.
Antiviral activity and antibody blocking assay in vitro
Endpoint dilution assay showed that TCID50 of VSV, NDV, and AIV titers was 10−7.26/100 μL, 10−4.75/100 μL, and 10−6.86/100 μL, respectively. To see whether PcIFN-α possesses biological function, we detected its antivirus activity and made a comparison with chicken IFN-α using cytopathic inhibition test in vitro. Renatured IFN-αs were diluted fourfold and preincubated in CEFs; after 24 h of VSV, NDV, and AIV incubation, the positive control displayed completely 100% cytopathic, and the dilution could confer a protective effect on CEFs in different levels. Negative controls were normal in CEFs. Moreover, PcIFN-α exerted superior antivirus activity when CEFs suffered from VSV and NDV compared with ChIFN-α. Neutralization assay of PcIFN-α antiviral protective activity results demonstrated that PcIFN-α preincubated with rabbit anti-PcIFN-α could not inhibit the replication of VSV. These antiviral activities were abrogated by rabbit anti-PcIFN-α antibodies in vitro (Fig. 4).
Neutralization of PcIFN-α antiviral protective activity.
Primary physicochemical characteristics of PcIFN-α
We tested the stability of PcIFN-α at 0.25% trypsin, pH 2, 4, 10, and 12 as well as 42°C, 56°C, and 65°C, respectively. Therefore, PcIFN-α was treated at different situations, then antiviral assay was conducted in CEFs/VSV system in parallel with untreated groups. Results showed that PcIFN-α was stable at changing pH and temperature for 2 h because no loss in antiviral activity was detected in treated and untreated samples (p > 0.05) (Fig. 5). On the contrary, after treatment with 0.25% trypsin, the antiviral activity was completely absent (data not shown), which is a common characteristic of
PcIFN-α induced the production of ISGs
To assess the effect of PcIFN-α on type I IFN response, the transcriptions of ISGs (including Mx-1, ISG15, and ISG56) were quantified. qRT-PCR analysis showed that the transcription of Mx-1, ISG15, and ISG56 increased sharply at 12 h in PcIFN-α-treated CEFs and sustained high expression levels until 24 h, and the expression of 24 h was significant (p < 0.01) than that of 3 h (Fig. 6A). Immunoblotting analysis showed that the Mx-1 protein was detected as a single major protein band of ∼76 kDa, whereas the amounts of Mx-1 protein gradually increased from 6 to 24 h after treatment. In addition, Mx-1 protein expression was much higher at 24 h than at 6 h (Fig. 6B). GAPDH was used as an endogenous control, and a 36 kDa protein band was detected in all the samples.

Analysis of ISGs transcription and Mx-1 protein expression in CEFs after PcIFN-α treatment.
Discussion
Dwelling in cultivated areas and stream sides, Indian peafowl is also known to forage around human habitats and villages for scraps. The movement of this species, no matter in the zoo or wild, is in proximity to domestic birds, most of which are always threatened to the point of becoming a disease outbreak (Khulape et al., 2014). IFNs, pleiotropic cytokines that regulate resistance to viral infections, have recently been reinvestigated intensively, as the interdependence of innate and adaptive immune responses became more evident (Gonzalez-Navajas et al., 2012). Interest in IFNs of birds has recently sparked from increasing problems with viral diseases in the poultry industry. Recombinant ChIFN-α has been found to be able to inhibit the replication of various viruses in many cell lines (Qu et al., 2013). However, information about Indian peafowl IFN-α is available. To determine its biological characteristics and potential protective effects on Indian peafowl, we characterized PcIFN-α in prokaryotic expression systems and determined their biological activities using bioinformatics analysis, Western blot, physicochemical property analysis, and cytopathic inhibition test.
In general, type I IFNs in birds and mammals have the typical intron lacking feature (Xu et al., 2013), which makes the method we cloned the PcIFN-α gene from genomic DNA feasible. Using genomic DNA, we successfully amplified the predicted 582 bp Indian peafowl IFN-α gene coding 193 amino acids. The molecular characteristics of PcIFN-α were analyzed using bioinformatics software. There was one potential N-glycosylation site in PcIFN-α mature protein. Besides, Cys-1, 30, 37, 98, 124, and 137 of mature PcIFN-α-formed disulfide bonds were crucial for its activity, which was conserved in some type I IFNs. Phylogenetic analysis revealed that IFN-αs were subdivided into two monophyletic lineages: avian and mammalian. The mammalian branch could be divided into carnivores and herbivores. The IFN-αs similarity of P. cristatus, Francolinus pintadeanus, C. japonica, G. gallus, were consistent with their grouping within the herbivorous monophyletic group (which was distinct from other carnivorous). Indeed, both gene conversion and gene duplication have shaped the evolution of the IFN-α gene family in eutherian species (Hughes, 1995). The features of PcIFN-α sequences provided more support for this view. As expected from the close phylogenetic relationship, the highest homology was observed with G. gallus and F. pintadeanus IFN-α sequences on the DNA as well as the protein level (>90%), suggesting that the IFN-α gene may have originated from gene duplication in a common ancestor during evolution.
Cytopathic inhibition test was carried out to compare the biological activities of PcIFN-α with ChIFN-α, which is an effective therapeutic agent used for viral infections clinically. In contrast, it was found that PcIFN-α protected CEFs from challenge with VSV, AIV, and NDV, indicating that PcIFN-α protein showed certain degrees of cross-species activity. This finding was not surprising considering the close relationship in evolutionary terms between peafowl and chicken. In contrast, by comparison, PcIFN-α exerted a considerable antiviral activity against VSV and NDV infections in CEFs, whereas ChIFN-α demonstrated superior antiviral activity against AIV infection in the same system (Table 2). Similar to ChIFN-α, PcIFN-α displayed low cytotoxicity in CEFs (Fig. 4A). In addition, PcIFN-α was verified to be highly sensitive to trypsin, insensitive to temperature, and stable despite changes in pH. Given the substantial antiviral activity and low cytotoxicity of PcIFN-α, it is a potential candidate for a novel, effective therapeutic agent.
AIV, avian influenza virus; CEF, chicken embryo fibroblasts; IFN-α, interferon-α; NDV, Newcastle disease virus; VSV, vesicular stomatitis virus.
ISGs (including Mx-1, ISG15, and ISG56) were obviously induced by PcIFN-α, exerting antiviral activity of IFNs and inhibiting viral replication, such as VSV, AIV, and NDV described in this study. The transcription of Mx-1 increased 6 h after PcIFN-α treatment, and peaked 24 h thereafter. This event was consistent with the expression of Mx-1 protein. The transcription of ISG15 and ISG56 showed a trend similar to that of Mx-1. Given that ISGs were downstream in the IFN signaling pathways, PcIFN-α can obviously induce ISGs expression in a time-dependent manner, suggesting that PcIFN-α can further inhibit viral replication and exert antiviral activity.
Conclusions
In this study, the characterization and biological activity of PcIFN-α were analyzed. PcIFN-α presented moderate biological activity, low cytotoxicity, and stable physicochemical characteristics, as well as exerted antiviral activity on CEF cells and a protective effect against VSV, AIV, and NDV. In addition, PcIFN-α induced the transcription of Mx1, ISG15, and ISG56 genes, as well as the expression of Mx1 protein in a time-dependent manner. Overall, our study on PcIFN-α enriched the current knowledge about the avian IFN-α family. All the consequences in our study provide a basis data for further research on PcIFN-α in future and these might provide clues to new preventative and therapeutic treatments to the peafowl.
Footnotes
Acknowledgments
The authors thank Gao Mingchun (College of Animal Medicine, Northeast Agricultural University, Harbin, China) for critical reading of the article and for expert technical assistance on the study. This work was supported by the National Key Research and Development Program of China (Grant No. 2017YFD0501702), the National Natural Science Foundation of China (Grant No. 31672619), the Fundamental Research Funds for the Central Universities (Grant No. 2572016EAJ5), and the Natural Science Foundation of Heilongjiang Province (Grant No. C2015061).
Disclosure Statement
No competing financial interests exist.