
Abstract
Maternal vitamin D deficiency in pregnancy has been associated with an increased risk of preeclampsia. Vascular endothelial dysfunction is a major phenotype of pregnancies with preeclampsia, contributing to increased maternal hypertension and proteinuria. We sought to determine whether vitamin D supplementation would alleviate preeclampsia associated endothelial dysfunction and explore the underlying mechanism using the reduced uterine perfusion pressure (RUPP) rat model. RUPP operated rats were supplemented with 1,25(OH)2D (RUPP+VD) on day 1, 7, and 14 of pregnancy by subcutaneous injection. On day 19 of pregnancy, after the measurement of blood pressure and urine collection, maternal blood serum and placenta samples were collected. 1,25(OH)2D treatment significantly improved endothelial dysfunction by reducing apoptosis and increasing nitric oxide (NO) production in blood vessels of RUPP operated rats compared to untreated RUPP rats. 1,25(OH)2D significantly down-regulated the expression of placental soluble FMS-like tyrosine kinase-1 (sFlt-1) in RUPP rats. Furthermore, the circulating sFlt-1 levels in maternal serum were positively correlated with the expression of placental sFlt-1 and were restored to a normal pregnant level by 1,25(OH)2D treatment in RUPP rats. Incubation of endothelial cell line with rat serum from RUPP+VD group significantly increased NO production and decreased caspase-3 activity compared with serum from untreated RUPP rats. Moreover, neutralization of sFlt-1 using the specific antibody mimicked the effect of 1,25(OH)2D, which abolished the deleterious effect of RUPP rat's serum on NO production and apoptosis. These results suggest that vitamin D supplementation is protective against RUPP induced endothelial dysfunction by downregulating placental sFlt-1, which can possibly alleviate preeclampsia associated symptoms.
Introduction
P
Maternal vascular endothelial dysfunction is a major phenotype of pregnancies with preeclampsia, contributing to increased peripheral vascular resistance, maternal hypertension, and proteinuria. Many studies have shown that the interaction between circulating factors and the maternal endothelium significantly contribute to generalized vascular dysfunction (Powe et al., 2011). Serum from pregnant women with preeclampsia induces endothelial injury and dysfunction in vitro (Myers et al., 2005). The serum circulating factors are partly of placental origin, providing a link between placental and maternal vascular dysfunction (Goulopoulou and Davidge, 2015).
It was reported that maternal vitamin D level is important during pregnancy. Lack of maternal vitamin D is reported to be associated with preeclampsia (Bodnar et al., 2007), and women with vitamin D deficiency have been found to have a significantly higher chance of developing preeclampsia (Tabesh et al., 2013). In addition, the incidence rate of preeclampsia is highest in winter months due to the less sunlight exposure resulting in a lower production of vitamin D (Magnus and Eskild, 2001).
Low vitamin D status in pregnant mice resulted in symptoms of preeclampsia as elevated blood pressure and impaired placental development (Liu et al., 2013). Furthermore, vitamin D supplementation in preeclampsia model rat showed a beneficial effect on preeclampsia associated symptoms (Tian et al., 2016). However, the underlying mechanism that links vitamin D with preeclampsia related symptoms is still unclear. In the present study, we investigated the effects of vitamin D on preeclampsia associated endothelial dysfunction and explored the underlying mechanism using the reduced uterine perfusion pressure (RUPP) rat model, which may provide experimental evidence for the justification of the clinical use of vitamin D in preeclampsia.
Materials and Methods
Animals
Three-month-old female Sprague-Dawley rats were obtained from Experimental Animal Center of Hebei Province, China. Rats were housed in a temperature controlled room (25°C) with a 12-h light/12-h dark cycle. The protocol for the reduced uterine perfusion pressure (RUPP) rat model was described previously (Granger et al., 2006; Tian et al., 2016). Pregnant rats were divided into three groups as follows: normal pregnant (NP, n = 10), pregnant with reduced uterine perfusion pressure (RUPP, n = 10), and pregnant RUPP plus 1,25(OH)2D (1,25-dihydroxyvitamin D) (RUPP+VD, n = 10). On day 1, 7, and 14 of pregnancy, RUPP+VD group of rats received 120 ng/100 g body weight of 1,25(OH)2D (injectable Calcijex, 1 mg/mL; Abbott Laboratories) by subcutaneous injection. The NP group and RUPP group received vitamin D vehicle (saline solution) subcutaneously (Masszi et al., 2013). On day 14 of pregnancy, the rats from the RUPP and RUPP+VD groups had RUPP surgeries performed as described previously (Tian et al., 2016), while rats in the NP group were sham operated controls. RUPP surgeries that resulted in maternal paraplegia or death were excluded from data analyses.
Twenty-four hour urine samples were collected at day 18 of gestation as described previously (Tian et al., 2016). On day 19 of gestation, arterial pressure was recorded continuously for 2 h with a pressure transducer (Cobe III Transducer) (Gilbert et al., 2007) after pressure stabilization. Rats were euthanized for blood collection through carotid artery catheter and tissue removal. Serum samples were frozen at −20°C until assayed. Rat tissues, including the placenta and maternal aorta, were excised immediately and weighed. The aorta was used to measure nitric oxide (NO) production, apoptosis, or stored at −80°C for western blot analysis. The placenta was used for enzyme-linked immunosorbent assay (ELISA). All procedures were carried out in accordance with the approval of the ethics committee of Hebei Medical University. Adequate steps were taken to ensure that animals did not suffer unnecessarily during the experiment.
Proteinuria assay
Proteinuria assay was performed as described previously (Tian et al., 2016). On day 18 of pregnancy, rats were placed into metabolic cages for the collection of 24-h urine samples. The volume of urine samples was recorded, and urine samples were stored at −20°C until analysis. Total urinary protein excretion was measured using coomassie brilliant blue G250 (n = 10 per group).
Enzyme-linked immunosorbent assay
Circulating and placental levels of vascular endothelial growth factor (VEGF), placental growth factor (PGF), soluble FMS-like tyrosine kinase-1 (sFlt-1), and soluble endoglin (sEng) were measured using commercial ELISA Kits available from R&D Systems (Quantikine) according to the manufacturer's directions. All assays were done in duplicate, and the protein levels were calculated using a standard curve derived from known concentrations of the respective recombinant proteins. The intra-assay variation for VEGF, PGF, sFlt-1, and sEng assay was 3.5%, 4.7%, 4.2%, and 5.90%, and the interassay variation for VEGF, PGF, sFlt-1, and sEng assay was 4.9%, 6.1%, 5.4%, and 7.3%, respectively.
Western blot
The protocol for western blot was described previously (Tian et al., 2016). Snap-frozen rat aorta was homogenized in a radio immunoprecipitation assay buffer (150 mM NaCl, 25 mM Tris-HCl, 1% Nonidet P-40, 0.5% sodium deoxycholate, 0.1% sodium dodecyl sulfate, pH 7.6) containing protease inhibitors and then centrifugated at 10,000 g for 20 min to collect the lysate. The lysates were probed with rabbit anti-eNOS (endothelial nitric oxide synthase) antibody (sc-8311; Santa Cruz), rabbit anti-phospho-eNOS (p-eNOS) antibody (sc-21871-R; Santa Cruz), mouse anti-caspase-3 antibody (sc-56053; Santa Cruz), and mouse anti-β-actin (A5441; Sigma). The images were quantitatively analyzed using ImageJ software (NIH).
Endothelial cell culture and treatments
We used the human endothelial cell line (EAhy926) as a model of endothelial cells within blood vessels. EAhy926 cells were cultured in low glucose Dulbecco's modified Eagle's medium (Wako Pure Chemicals) supplemented with 10% fetal bovine serum and 50 U/mL penicillin–streptomycin. After 24 h incubation with 20% rat serum from three groups (NP, RUPP, and RUPP+VD) and 40 μg/mL sFlt-1 neutralizing antibody (Tab-009cq-s; Creative Biolabs), endothelial cells were assessed for apoptosis and NO production.
NO measurement
NO production was estimated from the amounts of nitrite measured using NO2/NO3 Assay Kit-FX (Fluorometric), 2,3-Diaminonaphthalene Kit. For EAhy926 cells, culture medium was replaced with Krebs Ringer Buffer (was composed of 120 mM NaCl, 4.8 mM KCl, 0.54 mM CaCl2, 1.2 mM MgSO4, 15.9 mM Na2HPO4, 11 mM glucose, pH 7.4). After incubation for 30 min, aliquots were removed and prepared for the measurements according to the instructions provided by the manufacturer. Fluorescence was quantitated at an excitation wavelength of 360 nm and an emission wavelength of 460 nm. For blood vessels of NP, RUPP, and RUPP+VD rats, aorta segments were incubated in Krebs and stimulated with 10−7 M acetylcholine (ACh) for 30 min to measure NO production.
Caspase-3 activity assay
The activity of caspase-3 was determined by the Caspase Colorimetric Assay Kit (BioVision) according to the manufacturer's protocol. Briefly, 200 μg of total protein from aorta tissues or endothelial cells were loaded to each well of the 96-well plate and incubated with the caspase substrate at 37°C for 4 h. Samples were read at 405 nm in microplate spectrophotometer (BioTek). Caspase-3 activity was calculated against the mean value of caspase-3 activity from corresponding control.
Statistical analysis
Data are expressed as the mean ± standard deviation. Comparisons between three groups were made with ANOVA followed by post hoc Newman–Keuls multiple comparison test. Differences were considered statistically significant at p < 0.05. Statistical calculations were made with SPSS 17.0.
Results
1,25(OH)2D alleviates RUPP-induced hypertension and proteinuria in rats
To analyze the effects of 1,25(OH)2D on preeclampsia associated symptoms, we measured the blood pressure changes initially. We observed that the RUPP operation significantly increased the systolic blood pressure compared with NP group (134.30 ± 12.60 mmHg vs. 114.80 ± 8.60 mmHg, p < 0.01). In contrast, vitamin D treatment attenuated the elevation of systolic blood pressure induced by RUPP operation (120.10 ± 11.40 mmHg vs. 134.30 ± 12.60 mmHg, p = 0.015; Supplementary Fig. S1A; Supplementary Data are available online at
The RUPP operation caused an increase of 24 h urinary protein in rats (5.83 ± 2.30 mg vs. 2.90 ± 0.87 mg, p < 0.01), which is an important symptom in preeclampsia. Treatment with vitamin D decreased the amount of urinary protein excretion to 3.15 ± 1.22 mg (p < 0.01; Supplementary Fig. S1B). These data indicate that 1,25(OH)2D treatment attenuates the symptoms of preeclampsia.
1,25(OH)2D treatment increases vascular NO production and eNOS/p-eNOS levels in RUPP rats
NO production was measured in aortic tissue homogenate from NP, RUPP, and RUPP+VD rats after stimulation with 10−7 M ACh. NO production was significantly lower in RUPP group than the NP group (1010 ± 241 pmol/mg tissue vs. 1776 ± 178 pmol/mg tissue, p < 0.01) and higher in RUPP+VD group than RUPP group (1598 ± 206 pmol/mg tissue vs. 1010 ± 241 pmol/mg tissue, p < 0.01; Fig. 1A). Western blot analysis revealed that total eNOS and activated p-eNOS levels were affected by the RUPP operation and vitamin D treatment (Fig. 1B). Further analysis showed a significantly enhanced level of both eNOS and p-eNOS in RUPP+VD group versus the RUPP group, while these levels were reduced in RUPP versus NP rats (Fig. 1C, D). These data suggest that vitamin D treatment affects the expression of eNOS and p-eNOS, which in turn affects the NO production.
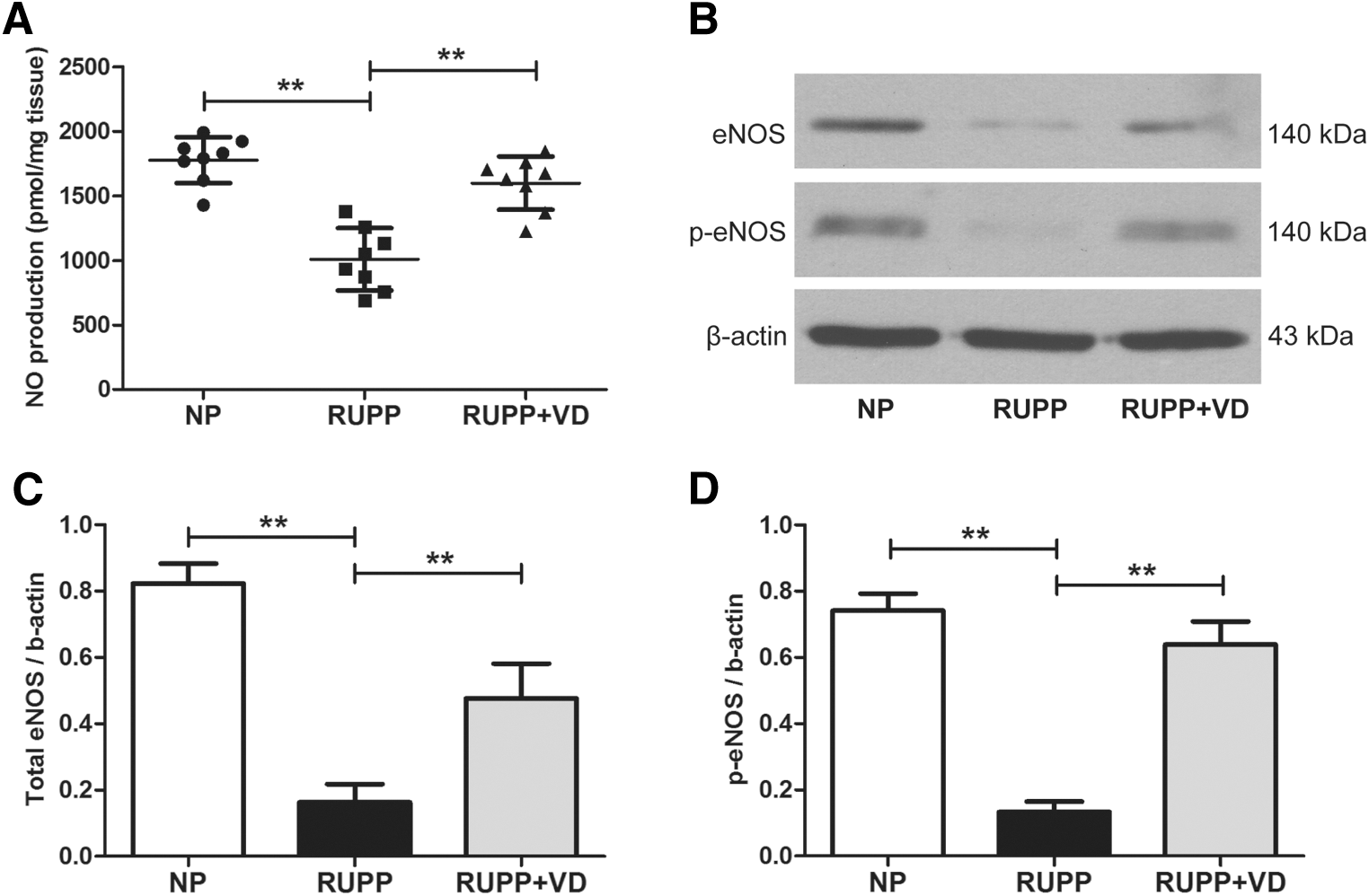
1,25(OH)2D treatment increases NO production in blood vessels of RUPP rats. ACh (10−7 M) induced NO production was measured in aortic tissue homogenate from NP, RUPP, and RUPP+VD rats
1,25(OH)2D inhibits caspase-3 mediated apoptosis in blood vessels of RUPP rats
Caspase-3 activity assay was performed to investigate whether 1,25(OH)2D could alleviate the vascular apoptosis induced by RUPP. Significantly increased caspase-3 activity was observed in RUPP group compared with NP group (54.2%, p < 0.01). In contrast, treating with vitamin D significantly reduced the caspase-3 activity by 36.9% (p < 0.01; Fig. 2A). Furthermore, western blot analysis showed that RUPP induced a significant increase of the cleavage of caspase-3, while vitamin D treatment significantly decreased the cleavage in RUPP group, which is a commonly used biomarker of apoptosis (p < 0.01; Fig. 2B, C).
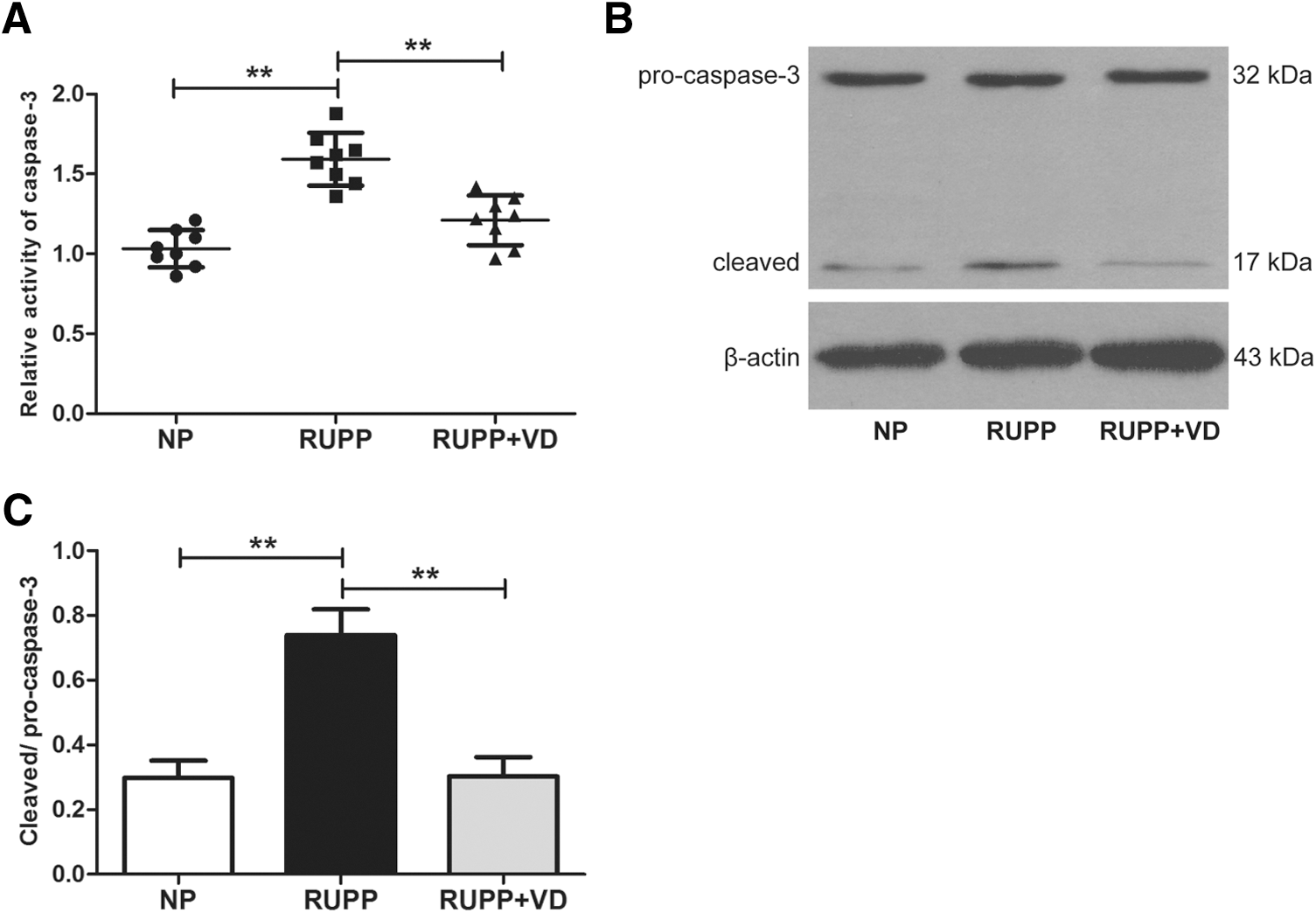
1,25(OH)2D treatment inhibits caspase-3 mediated apoptosis in blood vessels of RUPP rats. Aorta tissue lysates from NP, RUPP, and RUPP+VD rats were prepared for caspase-3 activity and western blot analysis.
1,25(OH)2D restores the increased expression of placenta-derived sFlt-1 in RUPP rats
The RUPP rats had significantly reduced placental weight (0.65 ± 0.06 mg vs. 0.52 ± 0.08 mg, p < 0.01) compared with NP rats. Vitamin D treatment improved this decrease of placental weight induced by the RUPP operation (0.61 ± 0.09 mg, p = 0.028; Fig. 3A).
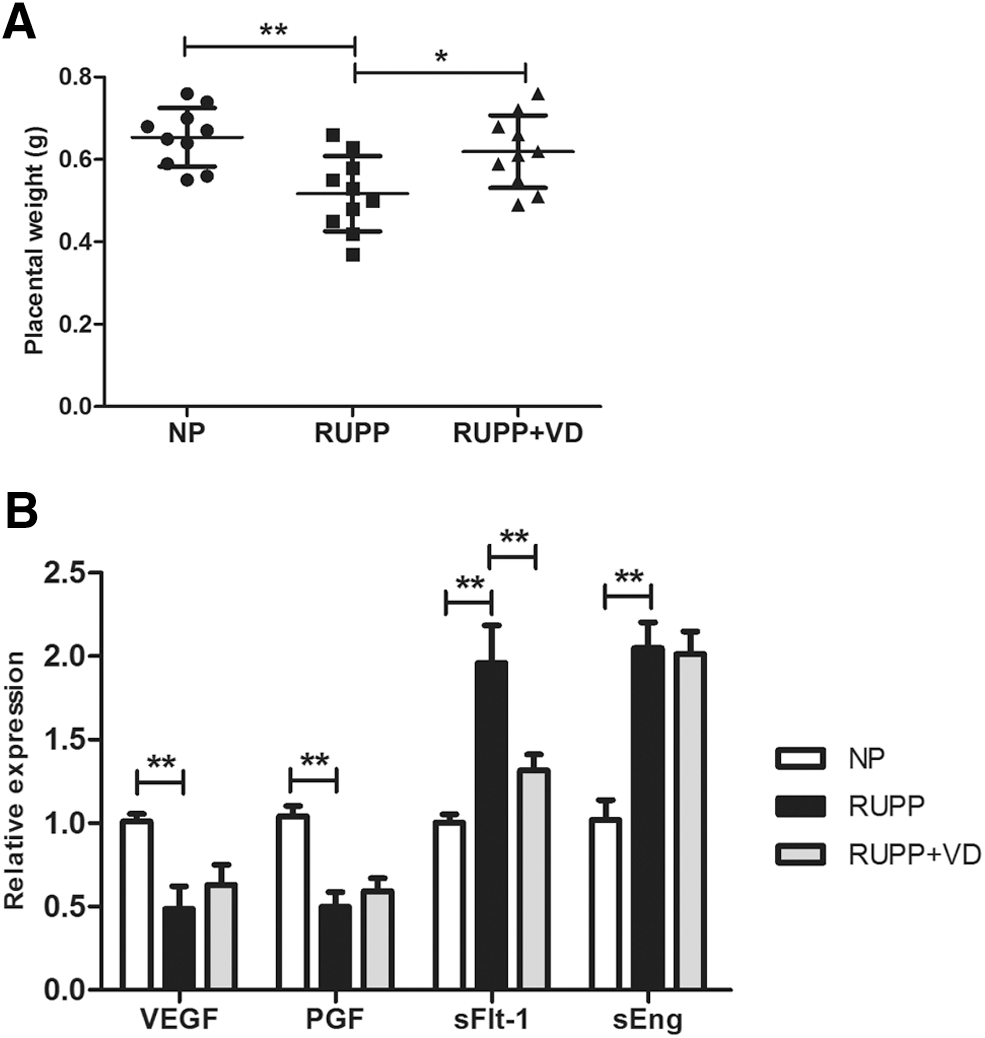
Effect of 1,25(OH)2D on placental weight and expression of placenta-derived circulating factors in RUPP rats. On day 19 of gestation, the placenta was excised and weighed in pregnant RUPP, RUPP+VD, and NP rats
Due to the strong association with placenta-derived circulating factors in vascular dysfunction of pregnancies with preeclampsia, we investigated the effect of 1,25(OH)2D on the expression of VEGF, PGF, sFlt-1, and sEng in the placenta of RUPP rats. The results were shown as fold change for each of these factors of the RUPP and RUPP+VD compared to the NP group. The RUPP operation caused a significantly decreased expression of VEGF and PGF by 2.05-fold (p < 0.01) and 2.08-fold (p < 0.01), respectively, compared to the NP group. On the contrary, the expression of sFlt-1 and sEng was significantly increased by 1.95-fold (p < 0.01) and 2.02-fold (p < 0.01), respectively, in placenta of RUPP rats compared with NP rats. Furthermore, the high expression of sFlt-1 induced in the RUPP group was significantly attenuated by vitamin D treatment (1.49-fold, p < 0.01; Fig. 3B).
1,25(OH)2D treatment decreases circulating sFlt-1 and increases circulating VEGF in RUPP rats
We further investigated the levels of circulating sFlt-1 and VEGF in rat serum. Serum sFlt-1 was significantly increased in RUPP rats compared with NP rats (211 ± 49 pg/mL vs. 70 ± 29 pg/mL, p < 0.01), and this was attenuated by vitamin D treatment (112 ± 40 pg/mL, p < 0.01; Fig. 4A). In contrast, serum VEGF level was significantly decreased in RUPP group (266 ± 39 pg/mL vs. 391 ± 41 pg/mL, p < 0.01) and rescued by vitamin D supplementation (330 ± 57 pg/mL, p = 0.019; Fig. 4B). The correlation between placenta-derived sFlt-1 and serum sFlt-1 was investigated. The expression level of placental sFlt-1 was positively correlated with serum sFlt-1 (r = 0.78, p < 0.001; Fig. 4C). Furthermore, the expression level of serum sFlt-1 was negatively correlated with serum VEGF (r = −0.69, p < 0.001; Fig. 4D). The results indicated that increased expression of placental sFlt-1 may contribute to the elevated circulating sFlt-1 level in RUPP rats.
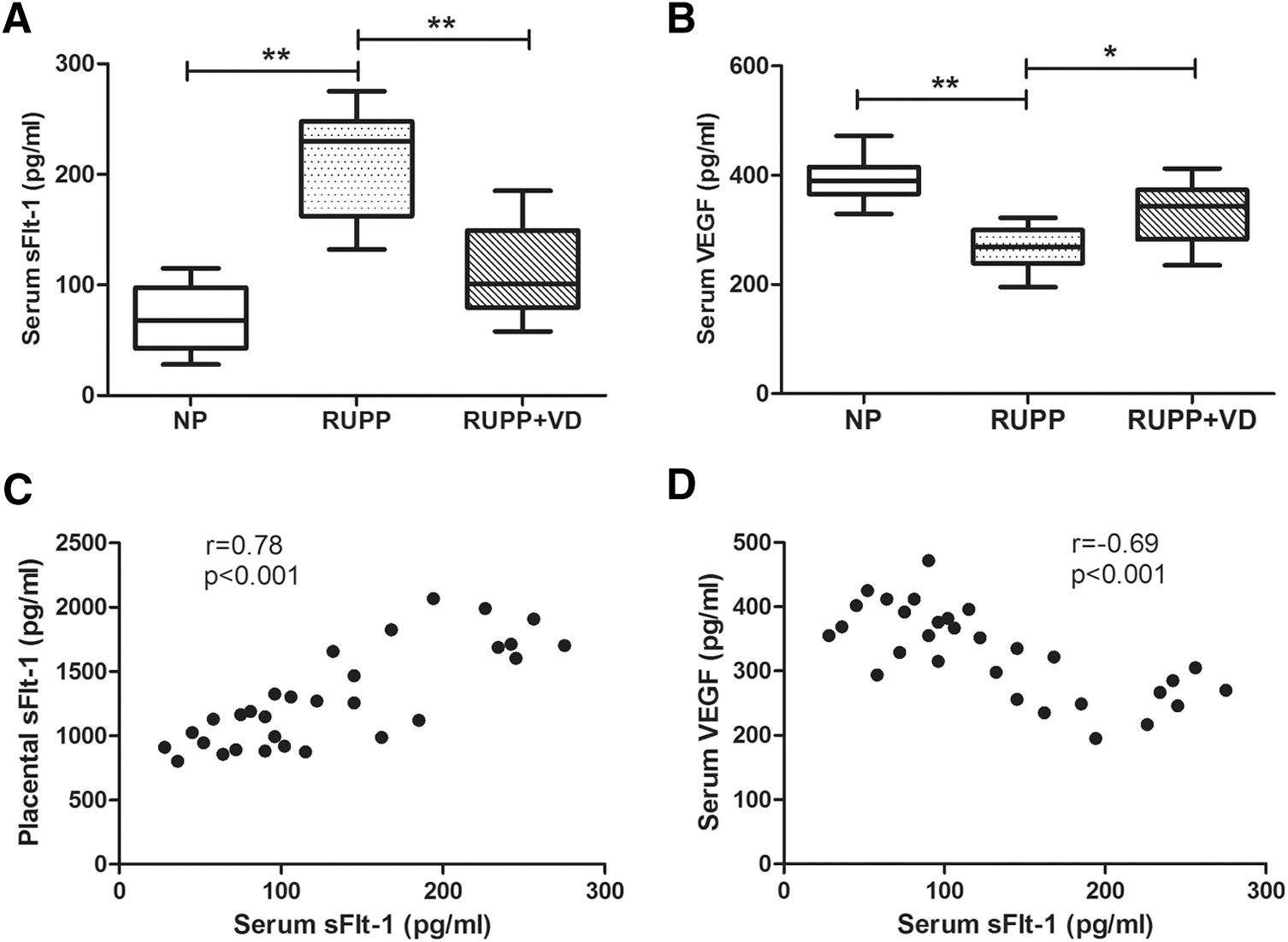
Effect of 1,25(OH)2D on circulating sFlt-1 and VEGF in RUPP rats. Serum of pregnant RUPP, RUPP+VD, and NP rats was collected and followed by ELISA measurement of sFlt-1
Neutralization of serum sFlt-1 mimics the effect of 1,25(OH)2D on endothelial dysfunction
To investigate the underlying mechanism of how vitamin D can alleviate preeclampsia related endothelial dysfunction, we further analyzed the effect of rat serum containing the circulating factors previously described on an endothelial cell line in culture. Rat serum from the RUPP group decreased NO production of the EAhy926 cell compared with the NP group (9.05 ± 1.04 pmol/mg protein vs. 11.97 ± 0.69 pmol/mg protein, p < 0.01), while serum from the RUPP+VD group significantly increased NO production compared with the RUPP group (10.92 ± 0.96 pmol/mg protein vs. 9.05 ± 1.04 pmol/mg protein, p = 0.012). Furthermore, neutralization of sFlt-1 using specific blocking antibody significantly abolished the deleterious effect of the rat serum from the RUPP group on NO production (11.32 ± 0.87 pmol/mg protein, p < 0.01; Fig. 5A and Supplementary Fig. S2).

Neutralization of serum sFlt-1 mimics the effect of 1,25(OH)2D on endothelial dysfunction. NO production
Caspase-3 activity was measured after incubation of endothelial cell line with rat serum. The results showed that the serum from RUPP group significantly increased apoptosis by 81.2% compared with NP group (p < 0.01). In contrast, vitamin D treatment rescued the RUPP induced apoptosis by 37.6% (p < 0.01). Similarly, neutralization of sFlt-1 mimicked the effect of vitamin D, which decreased the RUPP induced high caspase-3 activity by 43.2% (p < 0.01; Fig. 5B).
Taken together, these results indicated that 1,25(OH)2D alleviates RUPP induced preeclampsia related symptoms and endothelial dysfunction in rats, which could possibly work by regulating the expression of placenta-derived sFlt-1.
Discussion
The exact etiology of preeclampsia is currently unknown; therefore, it lacks effective prevention and prediction for preeclampsia. Maternal circulating concentrations of the active form of vitamin D, 1,25(OH)2D, increase significantly during early gestation. Multiple studies have shown associations between low maternal 1,25(OH)2D concentrations and the risk of preeclampsia (Bodnar et al., 2007; Robinson et al., 2010), whereas higher maternal circulating vitamin D concentrations were associated with a lower risk of preeclampsia (Mirzakhani et al., 2016). Insufficient placental perfusion in the RUPP model induces the same symptoms as human preeclampsia (McCarthy et al., 2011). In our study, the RUPP rats showed elevated blood pressure and increased urinary protein, which can be alleviated by 1,25(OH)2D treatment. These data confirm the findings from similar studies (Faulkner et al., 2016; Tian et al., 2016) and provide experimental evidence for the clinical use of vitamin D in preeclampsia.
Maternal endothelial dysfunction is a hallmark of preeclampsia, which significantly contributes to generalized vascular dysfunction, hypertension, and proteinuria (Hayman et al., 2000; Powe et al., 2011). Intracellular NO generation and apoptosis are surrogate markers of endothelial function (Di Marco et al., 2009). We investigated the effect of vitamin D treatment on NO production and apoptosis. Although Faulkner et al. (2016) showed that there is no change of circulating NO levels in RUPP rats treated with vitamin D compared with RUPP rats, our results indicated that 1,25(OH)2D supplementation significantly increased eNOS and p-eNOS level and NO production in aortic endothelium of RUPP rats. The increased NO production in endothelium may contribute to improving the local vascular function rather than circulating NO level. We also observed a significant decrease of RUPP induced apoptosis in rats treated with vitamin D. Our previous research showed that 1,25(OH)2D inhibited apoptosis in placenta of RUPP rats (Tian et al., 2016). The present study indicated that 1,25(OH)2D significantly prevented the blood vessels from RUPP induced apoptosis. These results suggest that the inhibition of endothelial cell apoptosis by vitamin D may be one common mechanism for improving placenta and vascular function in RUPP rats. We propose that understanding the mechanism underlying how vitamin D improved the preeclampsia associated endothelial dysfunction will provide insight into the clinical treatment of preeclampsia.
Many studies have shown that the interaction between circulating factors and maternal endothelium significantly contribute to generalized vascular dysfunction (Myers et al., 2005; Kao et al., 2016). In addition, the placenta plays a cardinal role in the pathogenesis of preeclampsia because delivery of the placenta resolves the clinical symptoms (Pratt et al., 2015). In response to reduced perfusion, the placenta produces abnormal levels of circulating factors that act upon the mother to bring about the clinical symptoms of preeclampsia (Roberts and Escudero, 2012). There is also evidence that maternal vitamin D supplementation may impact gene expression in placenta (Schulz et al., 2017). We found significantly decreased placental weights in RUPP rats and which were rescued by vitamin D treatment that confirms what was reported in previous studies (Faulkner et al., 2016; Tian et al., 2016). Moreover, there were significant changes of placental-derived circulating factors (VEGF, PGF, sFlt-1, and sEng) in placenta of RUPP rats compared with NP rats. Furthermore, with vitamin D treatment, the expression of sFlt-1 was obviously restored in RUPP rats. Interestingly, we did not observe changes in placental expression of sEng, VEGF, and PGF in RUPP rats treated with vitamin D, which suggests a different regulatory mechanism of these factors. In addition, unlike sFlt-1, there is evidence that the placenta may be not a major source of the elevated level of sEng in human preeclampsia (Paasche Roland et al., 2012).
sFlt-1 is a soluble splice variant of VEGF receptor and negatively regulates the actions of VEGF by binding free circulating VEGF and occupying the VEGF receptor (Zhai et al., 2014). It is well documented that elevated circulating level of sFlt-1 was strongly correlated with preeclampsia (George et al., 2011). The elevated sFlt-1 in preeclampsia patients leads to widespread endothelial dysfunction and the clinical manifestations of proteinuria and hypertension (Maynard et al., 2003; Cindrova-Davies et al., 2011). Furthermore, overexpression of exogenous sFlt-1 induces a condition mimicking preeclampsia in pregnant rats (Murphy et al., 2010; Holwerda et al., 2014). We found that the serum sFlt-1 level positively correlated with the expression of placental sFlt-1 and was significantly elevated in RUPP group and was rescued by vitamin D supplementation. Furthermore, neutralizing the excess serum sFlt-1 of RUPP rats rescued the endothelial dysfunction in these rats. We also observed that the circulating VEGF level was negatively correlated with serum sFlt-1, which suggested the possible antagonistic effect of sFlt-1.
Conclusion
In conclusion, we reported that vitamin D treatment attenuated RUPP induced maternal vascular dysfunction. In addition, for the first time, we showed that vitamin D could downregulate the placental expression of sFlt-1, which in turn contributes to restoring the excess production of serum sFlt-1. We also demonstrated that neutralizing serum sFlt-1 rescued RUPP induced endothelial injury, which mimicked the effect of vitamin D in this study. These findings suggest that vitamin D could alleviate preeclampsia associated symptoms by restoring the excess sFlt-1 in preeclampsia. Further studies are needed to combine vitamin D supplementation with antagonizing sFlt-1 in maternal circulation for preeclampsia treatment.
Footnotes
Acknowledgments
This work was supported by grants from the Health and Family Planning Commission of Hebei (ZD20140388) and the State Administration of Traditional Chinese Medicine of Hebei (2014141).
Disclosure Statement
No competing financial interests exist.
References
Supplementary Material
Please find the following supplemental material available below.
For Open Access articles published under a Creative Commons License, all supplemental material carries the same license as the article it is associated with.
For non-Open Access articles published, all supplemental material carries a non-exclusive license, and permission requests for re-use of supplemental material or any part of supplemental material shall be sent directly to the copyright owner as specified in the copyright notice associated with the article.