
Abstract
microRNAs have been reported to play important roles in the pathogenesis of rheumatoid arthritis (RA). This study examined the effects of miR-522 on the biological behaviors of RA synovial fibroblasts. The expression levels of miR-522 and relevant genes were measured by quantitative real-time PCR. The protein levels of cytokines were determined by ELISA assay. The protein levels of matrix metalloproteinases (MMPs) and suppressor of cytokine signaling 3 (SOCS3) were determined by western blot assay. Luciferase reporter assay was used to confirm the potential target of miR-522. Our results showed that miR-522 was upregulated in synovial fibroblasts from RA patients, and miR-522 expression level was significantly associated with the RA-associated clinical parameters. miR-522 overexpression increased the mRNA and protein expression levels of tumor necrosis factor-α (TNF-α), interleukin-1β (IL-1β) and MMPs (MMP-1, MMP-3, and MMP-13) in RA synovial fibroblasts. Lipopolysaccharide induced the upregulation of TNF-α, IL-1β, and MMPs in RA synovial fibroblasts, which was reversed by miR-522 knockdown. Bioinformatics analysis identified SOCS3 as a potential target of miR-522, and this target of miR-522 was confirmed by luciferase reporter assay, and miR-522 overexpression suppressed the mRNA and protein expression levels of SOCS3. The enforced expression of SOCS3 attenuated the enhanced effects of miR-522 on mRNA expression levels of TNF-α, IL-1β, and MMPs. Collectively, our results suggested that miR-522 regulated the expression of proinflammatory cytokines and MMPs partly via targeting SOCS3 in RA synovial fibroblasts, which may contribute to pathogenesis of RA.
Introduction
R
Recently, evidence from various studies indicated that RA was associated with both genetic and environmental factors. RA synovial fibroblasts were found to be involved in RA aggressiveness by production abundant cytokines and matrix metalloproteinases (MMPs), and a large number of studies reported that RA synovial fibroblasts play crucial roles in RA (Philippe et al., 2012). In this regard, identifying the novel targets to regulate the abnormal biological behaviors of RA synovial fibroblasts may provide us with new strategy to treat RA.
microRNAs (miRNAs) are short noncoding RNAs and mediate mRNA translational repression or degradation through base pairing to 3′ untranslated region (3′UTR) of mRNA (Ambros et al., 2003), and functional studies demonstrated that miRNAs are involved in the regulation of epigenetic anomalies (Fromm et al., 2015). Several studies have detected the aberrant expression of miRNAs in RA, including miR-203, miR-155, miR-346, and so on, and the aberrant expression of miRNAs was found to be presented in peripheral blood mononuclear cells, synovial fibroblasts and synovial tissue from patients with RA. These miRNAs were shown to exert their functional roles via regulating specific signaling pathways, which in turn contribute to the pathogenesis of RA (Stanczyk et al., 2008, 2011; Alsaleh et al., 2009). In addition, in vivo studies by using arthritic mice showed that intra-articular injection of miR-15a could induce cell apoptosis by repressing the translation of Bcl-2 in arthritic synovium (Nagata et al., 2009).
Recent studies have shown that some common miRNAs differently expressed in Kashin–Beck disease, osteoarthritis and RA, and miR-522 were found to be upregulated in the blood specimens from the patients with RA (Wu et al., 2017), which suggests that dysregulation of miR-522 may contribute to the pathogenesis of miR-522. miR-522 belongs to the family of the chromosome 19 miRNA cluster, and is the largest miRNA cluster in the human genome (Tan et al., 2014). Although the functional role of miR-522 was well documented in different types of cancers, its role in the pathogenesis of RA is largely unknown, thus it is of great scientific interest to further elucidate the potential role of miR-522 in RA.
Suppressor of cytokine signaling 3 (SOCS3) belongs to the suppressor of cytokine signaling family, and SOCS3 was found to be key negative intracellular regulators of cytokine and Toll-like receptor responses (Veenbergen et al., 2011), and accumulating evidence suggested that SOCS3 plays an important role in the regulation of immune cell development and function. Recent studies have demonstrated that intravenously induced overexpression of SOCS3 is effective in preventing collagen-induced arthritis (Veenbergen et al., 2008). In addition, SOCS3 was shown to take part in the cholinergic pathway regulation of synovitis in RA (Li et al., 2017), indicating the potential functional role of SOCS3 in RA. Studies have identified that several miRNAs target SOCS3 to regulate the inflammatory process. For example, miR-19a-mediated reduction of SOCS3 enhanced interferon-α and interleukin (IL)-6 signal transduction through STAT3 (Collins et al., 2013). However, whether SOCS3 is regulated by miRNAs that involved in RA remains unclear.
In the present study, we examined the expression of miR-522 in the RA synovial fibroblasts from RA patients, and we also performed a series of in vitro functional studies to elucidate the molecular mechanisms of miR-522-mediated biological behaviors of RA synovial fibroblasts.
Materials and Methods
Synovial tissue collection
Synovial tissues from 30 RA patients were obtained during joint surgery at Shaoxing People's Hospital from 2014 to 2016. All the RA patients meet the American College of Rheumatology criteria for classification of disease. All patients had Health assessment questionnaire [HAQ score, a questionnaire for the assessment of
Healthy control tissues were obtained from 25 patients with joint trauma undergoing joint replacement surgery at Shaoxing People's Hospital from 2014 to 2016. The control patients were free of other diseases such as infectious disease, autoimmune disease, and cancer. The study was approved by the Ethics Committee of Shaoxing People's Hospital, and all the recruited patients signed the written informed consent. The clinical characteristics of the all recruited patients were shown in Table 1. The collected samples were then processed for further experimentation.
CCP, cyclic citrullinated peptide; DAS 28, disease activity score of 28 joints; ESR, erythrocyte sedimentation rate; HAQ, health assessment questionnaire; IL-1β, interleukin-1β; RA, rheumatoid arthritis; TNF-α, tumor necrosis factor alpha.
Synovial fibroblasts culture and transfection
The synovial tissues of knee joint were collected during the joint replacement surgery of RA patients and control patients. The synovial tissues were washed with sterile PBS and cut into pieces. Then, it was digested using collagenase (Wako, Osaka, Japan) and DNAase (Sigma, St. Louis, MO) at 37°C for 2 h. After centrifugation, the synovial fibroblasts were collected and cultured in DMEM medium (Thermo Fisher Scientific, Waltham, MA) supplemented with 5 mM glucose and 10% fetal bovine serum (FBS; Thermo Fisher Scientific) in a humidified atmosphere of 5% CO2 at 37°C. All the in vitro experiments were performed at least in triplicates.
Oligonucleotides, cell transfection, and lipopolysaccharide stimulation
miR-522 mimics, miR-522 inhibitor, and their respective controls were purchased from RiboBio (Guangzhou, China). The pcDNA3.1 vector and SOCS3-overexpressing vector (pcDNA3.1-SOCS3) were purchased from GenePharma (Shanghai, China). Transfection was performed in Opti-MEM medium with Lipofectamine 2000 following manufacturer's instructions. The cells were collected at 24 h posttransfection. Synovial fibroblasts were treated with lipopolysaccharide (LPS) (100 ng/mL) alone, LPS (100 ng/mL)+miR-522 inhibitor, or LPS (100 ng/mL)+inhibitor control for 24 h. After the treatment, cells were collected for further experimentation.
RNA extraction and quantitative real-time PCR
Total of 100 ng RNA were extracted from cells using RNAiso Plus reagent (Takara, Dalian, China) following the manufacturer's instructions. Then, the RNA was reverse-transcribed to cDNA, using PrimeScript 1st strand cDNA Synthesis Kit (for miRNA) or PrimeScript RT Master Mix (for mRNA) (Takara). The real-time PCR was performed using SYBR Green Real-Time PCR Master Mixes (Takara) on an ABI 7900 PCR system (Applied Biosystems, Foster City, CA). U6 was used as an internal control for miRNA expression, and GAPDH was used as an internal control for mRNA expression. The primers were shown in Supplementary Table S1 (Supplementary Data are available online at
Western blotting
Proteins were extracted from synovial fibroblasts by using the lysis buffer and the proteins from whole cell lysates were prepared and quantified by BCA Protein Assay Kit. The proteins (30 μg) were separated by sodium dodecyl sulfate polyacrylamide gel electrophoresis (SDS-PAGE) and transferred onto PVDF membranes. Subsequently, the membranes were blocked with 5% skimmed milk at room temperature for 1 h and then incubated with MMP-1 (Abcam), MMP-3 (Cell signaling Technology, Danvers, MA), MMP-13 (Cell Signaling Technology), SOCS3 (Abcam), or β-actin primary antibodies (Abcam) at 4°C overnight, followed by washing three times with PBST. The membrane was then incubated with HRP-conjugated secondary antibodies at room temperature for 2 h. Protein was visualized by the enhanced chemiluminescence system (Roche, Basel, Switzerland). β-Actin was used as the internal control.
ELISA for TNF-α and IL-1β
The protein levels of secreted TNF-α and IL-1β in RA synovial fibroblasts after different treatments were measured by an enzyme immunoassay, using the human TNF-α and IL-1β ELISA Kits (Abcam).
Luciferase reporter assay
For the luciferase reporter vector construction, a wild-type (SOCS3-WT) or mutated (SOCS3-MUT) fragment of SOCS3 3′UTR that contained the putative miR-522 binding sites was cloned into a pMIR-Report vector (Thermo Fisher Scientific). Synovial fibroblasts were cotransfected with pMIR constructs, along with miR-522 mimics or miR-522 inhibitor or their respective controls. At 48 h posttransfection, firefly and Renilla luciferase activities were measured by Dual Luciferase Assay (Promega, Madison, WI) to determine the luciferase reporter activities.
Data analyses
Statistical analyses were performed using GraphPad Prism software. Correlations between miR-522 expression and patients' clinical data were analyzed by the Spearman's rank correlation test. The data are presented as mean ± SD. Student's t-test or one-way ANOVA was used for the comparison of significant differences among treatment groups. p < 0.05 was considered statistically significant.
Results
miR-522 was upregulated in synovial fibroblasts from RA patients and correlates with DAS28, anti-CCP, TNF-α, IL-1β, and ESR in RA patients
The clinical characteristics of all the RA patients and control subjects are summarized in Table 1. For the DAS28 score, the mean value was 5.6, which implies high disease activity in the RA patients. Anti-CCP is an antibody present in most RA patients, and the RA group showed a markedly increased anti-CCP (485.5 ± 264.5 U/mL) compared with normal group (12.6 ± 4.8 U/mL) (Table 1). For the ESR, the levels of ESR were significantly increased in the RA group (46.5 ± 31.2 mm/h) compared with that in normal group (8.5 ± 2.7 mm/h) (Table 1), suggesting higher degree of inflammation in the joints from RA patients. TNF-α and IL-1β were found to play key roles in the pathogenesis of RA and were significantly higher in the RA patients than that of normal controls (Table 1). For the age and gender, no significant differences were found between groups (Table 1).
The expression levels of miR-522 in synovial fibroblasts were determined by quantitative real-time PCR (qRT-PCR), and the expression levels of miR-522 in synovial fibroblasts from RA patients were markedly higher than that from normal group (Fig. 1A). The correlations between miR-522 expression levels and the clinical parameters were also examined. In the RA patients, miR-522 expression levels positively correlated with DAS 28 (r = 0.1589, p = 0.0291), anti-CCP (r = 0.4651, p = 0.0096), TNF-α (r = 0.3772, p = 0.0399), IL-1β (r = 0.4651, p = 0.0096), and ESR (r = 0.4634, p = 0.0099) (Fig. 1B–F and Table 2). There was no significant correlation between miR-522 expression and the other clinical parameters, including age, gender, and duration of disease (Table 2).
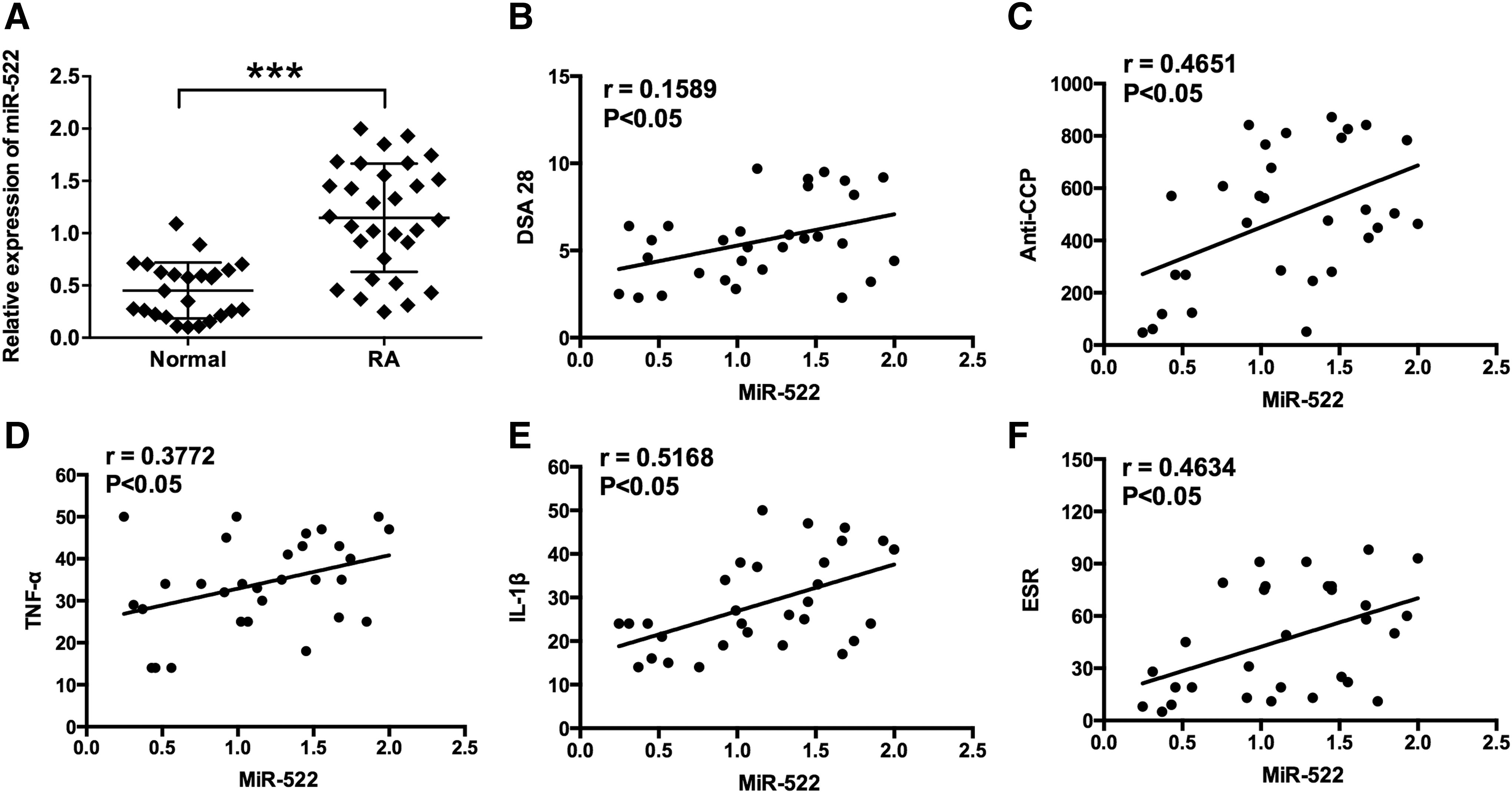
miR-522 was upregulated in fibroblast-like synoviocytes from patients with RA and its correlation with clinical characteristics.
Upregulation of miR-522 increased the expression of TNF-α and IL-1β in RA synovial fibroblasts
The effect of miR-522 overexpression on the expression of TNF-α and IL-1β was investigated in RA synovial fibroblasts. The relative expression levels of miR-522 were upregulated after transfecting miR-522 mimics in RA synovial fibroblasts (Fig. 2A). miR-522 mimics increased the mRNA expression levels of TNF-α and IL-1β in RA synovial fibroblasts comparing to mimics control (Fig. 2B, C). At protein levels, the ELISA assay revealed that the secreted protein levels of TNF-α and IL-1β were significantly increased in RA synovial fibroblasts transfected with miR-522 mimics comparing with that of mimics control (Fig. 2D, E).
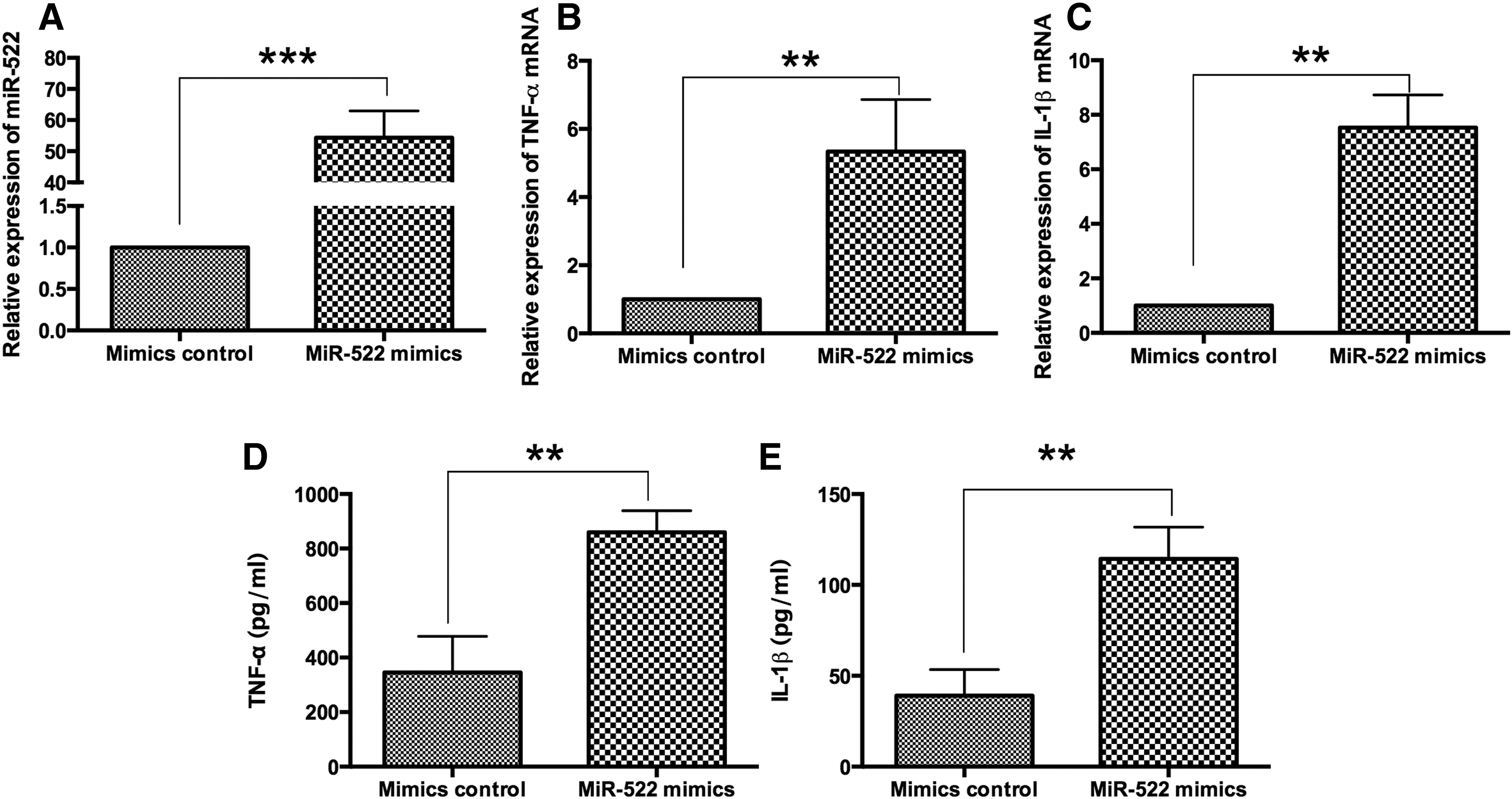
The effects of miR-522 overexpression on the expression levels of TNF-α and IL-1β in RA synovial fibroblasts.
Upregulation of miR-522 promoted the expression of MMPs in RA synovial fibroblasts
The mRNA and protein expression levels of MMPs, including MMP-1, MMP-3, and MMP-13, were examined by qRT-PCR and western blot assay, respectively, in RA synovial fibroblasts that received different treatments. The qRT-PCR results showed that the mRNA expression levels of MMP-1, MMP-3, and MMP-13 were significantly upregulated in RA synovial fibroblasts transfected with miR-522 mimics when compared with control group (Fig. 3A, B). In consistent with the qRT-PCR results, the protein expression levels of MMP-1, MMP-3, and MMP-13 were markedly upregulated in RA synovial fibroblasts transfected with miR-522 mimics when compared with control group (Fig. 3C, D).
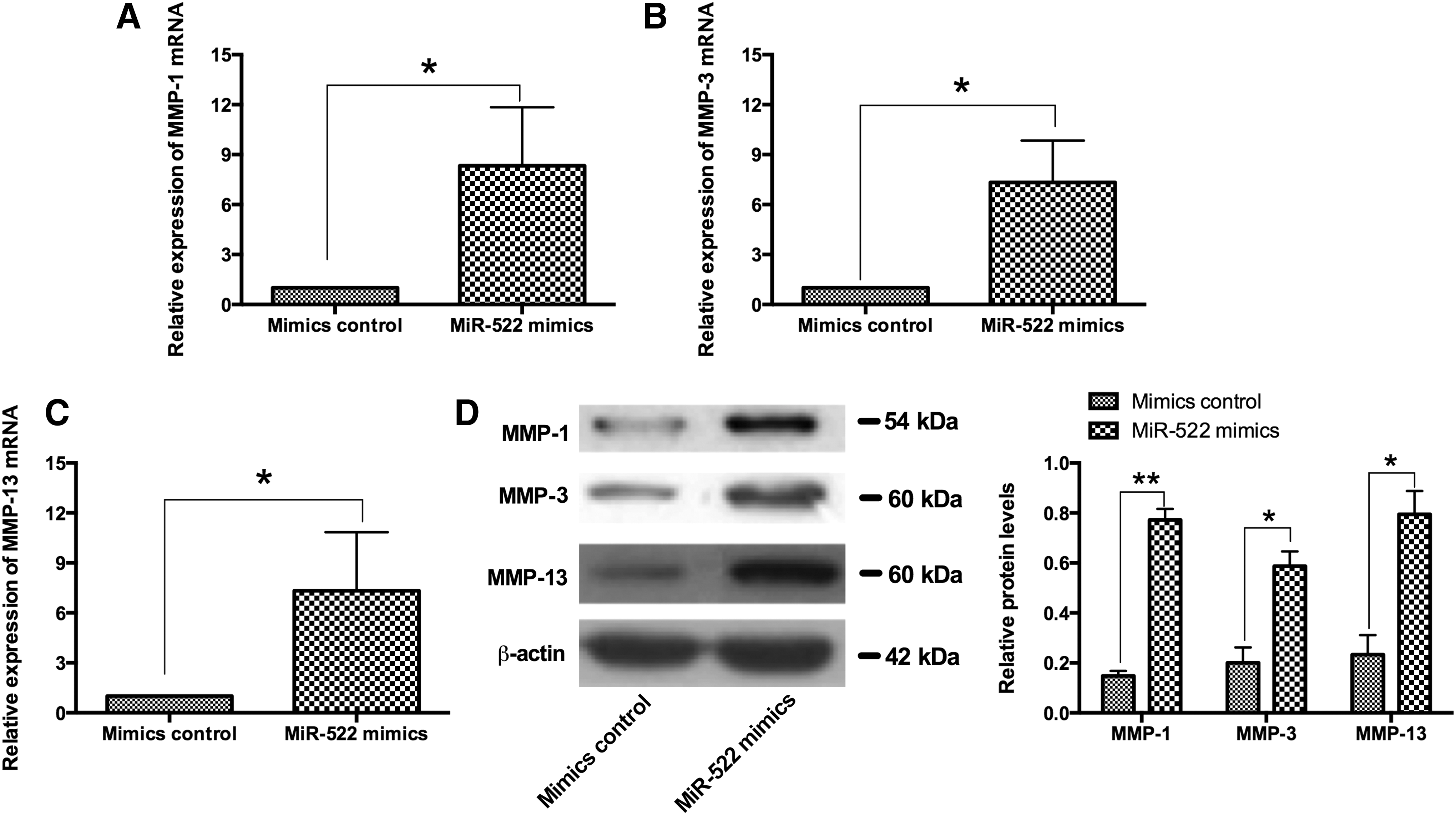
The effects of miR-522 overexpression and the expression levels of MMP-1, MMP-3, and MMP-13 in RA synovial fibroblasts. The relative mRNA expression levels of
Knockdown of miR-522 suppressed the expression of TNF-α and IL-1β induced by LPS in RA synovial fibroblasts
To address the effects of miR-522 knockdown on the proinflammatory cytokines production, the LPS-stimulated inflammatory response model was used. Knockdown of miR-522 was achieved efficiently by transfection of miR-522 inhibitor in RA synovial fibroblasts (Fig. 4A). Furthermore, the mRNA and protein expression levels of TNF-α and IL-1β were measured by qRT-PCR and western blot assay, respectively, and LPS treatment increased the mRNA and protein expression levels of TNF-α and IL-1β in RA synovial fibroblasts when compared to negative control (NC) group (Fig. 4B–E). The LPS-induced upregulation of the mRNA and protein expression levels of TNF-α and IL-1β were suppressed by the transfection of miR-522 inhibitor in RA synovial fibroblasts when compared with transfection of miR-522 inhibitor control (Fig. 4B–E).
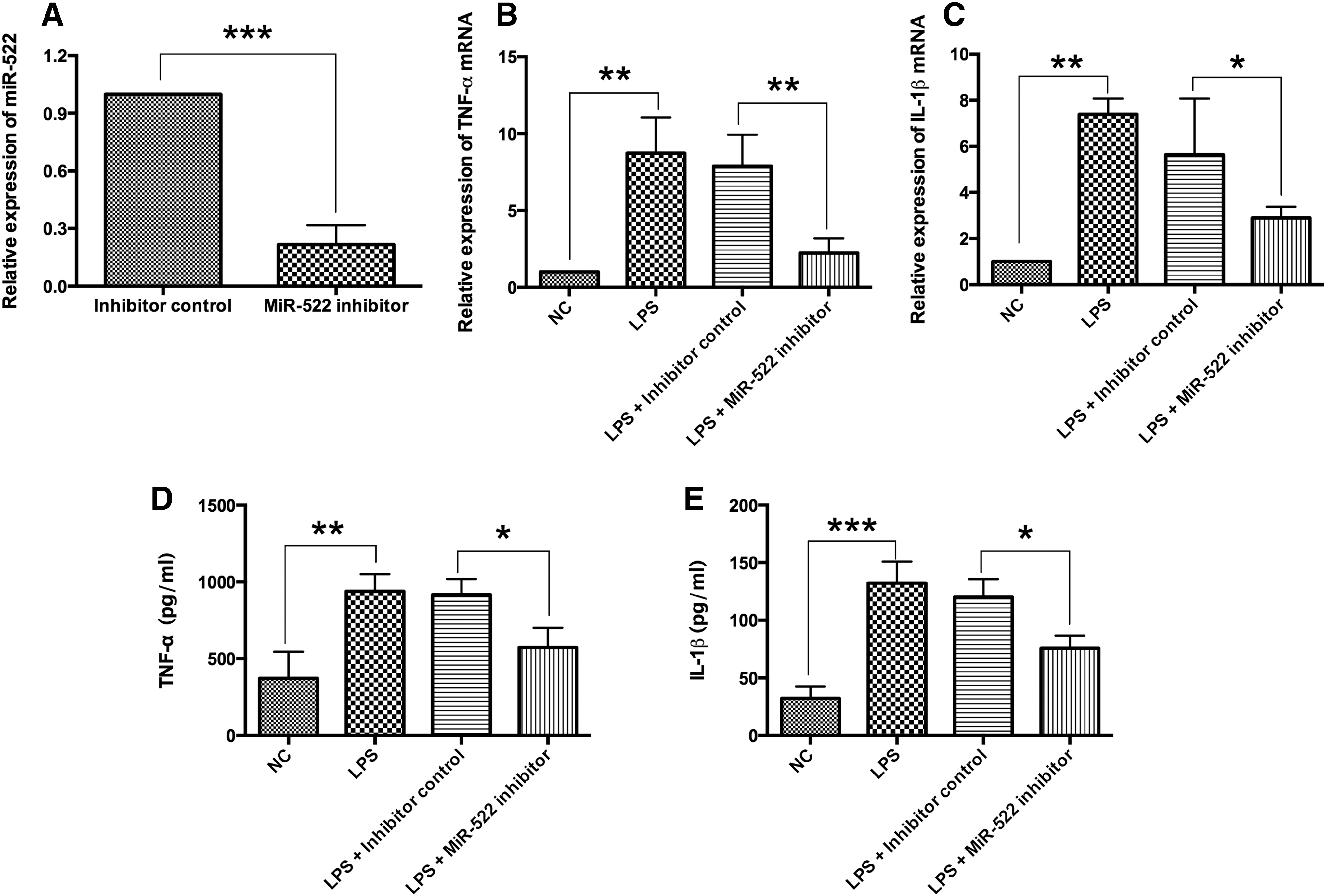
The effects of miR-522 silencing on the expression levels of TNF-α and IL-1β in LPS-treated RA synovial fibroblasts.
Knockdown of miR-522 suppressed the expression of MMPs stimulated by LPS in RA synovial fibroblasts
Similarly, overexpression of MMP-1, MMP-3, and MMP-13 mRNAs and proteins were observed in LPS-treated RA synovial fibroblasts. Compared with the NC group, LPS significantly increased the mRNA and protein expression levels of MMP-1, MMP-3, and MMP-13 (Fig. 5A–D). The LPS-induced upregulation of the mRNA and protein expression levels of MMP-1, MMP-3, and MMP-13 were reversed in RA synovial fibroblasts when transfecting with miR-522 inhibitor (Fig. 5A–D).
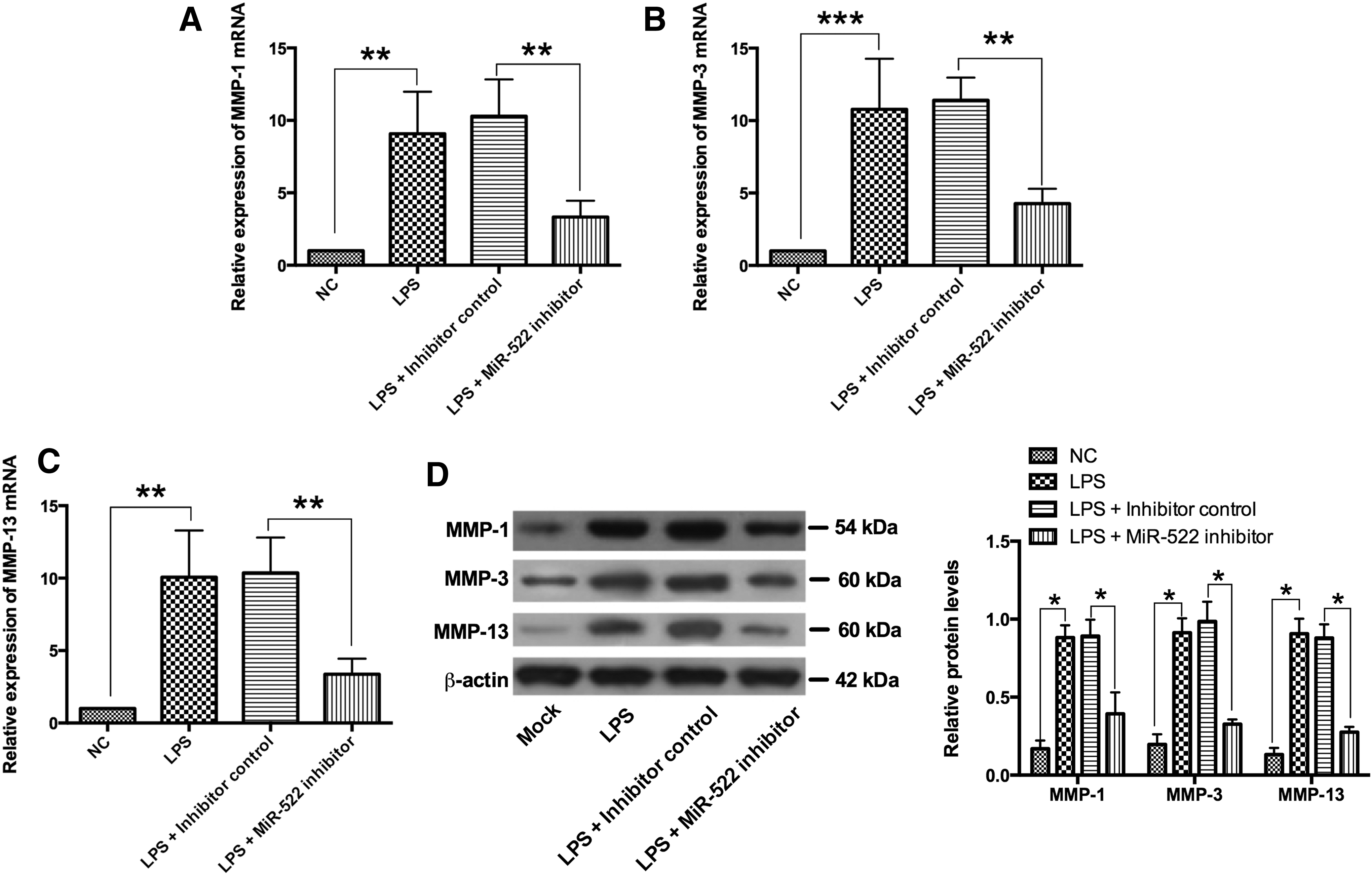
The effects of miR-522 silencing the expression levels of MMP-1, MMP-3, and MMP-13 in LPS-treated RA synovial fibroblasts. The relative mRNA expression levels of
miR-522 repressed SOCS3 expression by targeting its 3′TUR
We screened the potential targets of miR-522 by TargetScan, and SOCS3 was on the top of the list. The 3′UTR of SOCS3 contains a binding site of miR-522. A luciferase reporter vector containing wild-type 3′UTR of SOCS3 (WT) or the mutant one (MUT) was constructed to confirm the binding of SOCS3 and miR-522 (Fig. 6A). The luciferase activity was decreased significantly when transfecting with miR-522 mimics and luciferase reporter vector containing wild-type 3′UTR of SOCS3 (WT) in RA synovial fibroblasts (Fig. 6B). The luciferase activity of mutant 3′UTR vector was unaffected (Fig. 6C). Furthermore, the effects of ectopic overexpression of miR-522 on the expression of SOSC3 were examined in RA synovial fibroblasts. Overexpression of miR-522 caused dramatically reduction of SOCS3 at both mRNA and protein levels comparing with mimics control group (Fig. 6D, E).
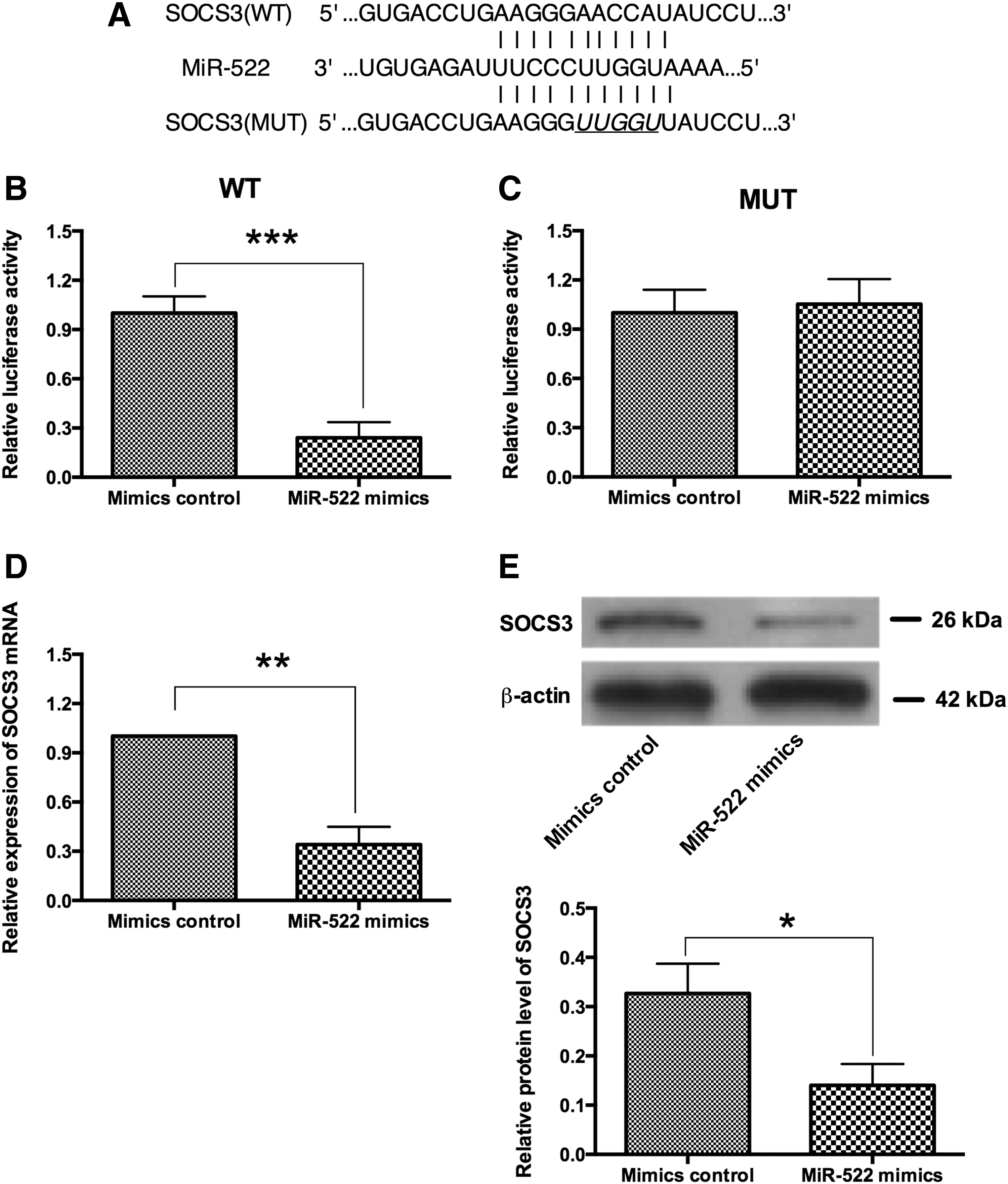
miR-522 targets 3′UTR of SOCS3 gene and suppresses its expression.
SOCS3 attenuated the effects of miR-522 in RA synovial fibroblasts
The mRNA and protein expression levels of SOCS3 were significantly increased in RA synovial fibroblasts after transfection with pcDNA3.1-SOCS3 comparing with pcDNA3.1 group (Fig. 7A, B). As expected, cotransfection with pcDNA3.1+miR-522 mimics significantly increased the mRNA expression levels of TNF-α, IL-1β, MMP-1, MMP-3, and MMP-13 in RA synovial fibroblasts comparing with pcDNA3.1+mimics control group (Fig. 7C, D); and cotransfection with pcDNA3.1-SOCS3+miR-522 mimics partially attenuated the enhanced effects of miR-522 overexpression on the mRNA expression levels of TNF-α, IL-1β, MMP-1, MMP-3, and MMP-13 in RA synovial fibroblasts comparing with pcDNA3.1+miR-522 mimics group (Fig. 7C, D).
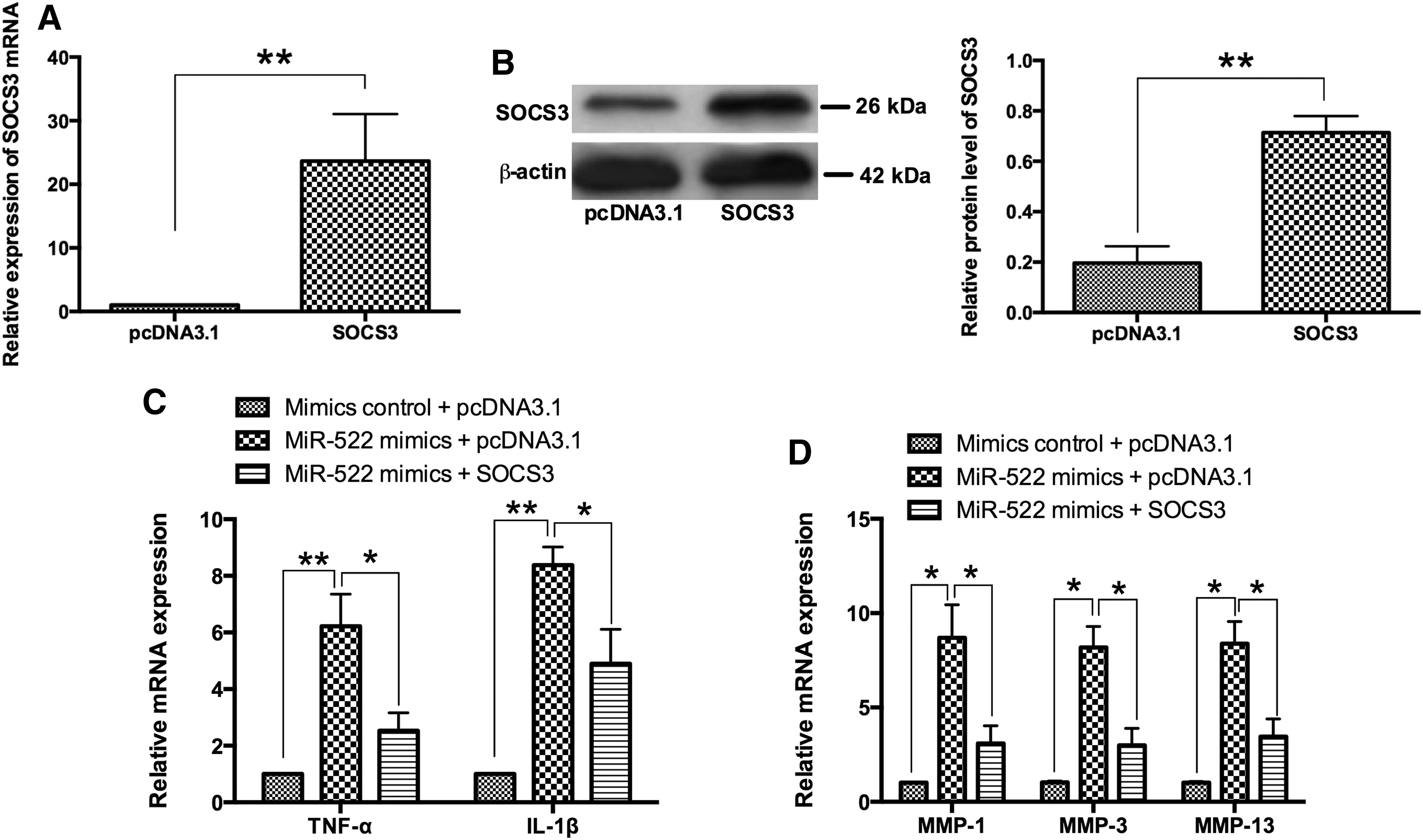
SOCS3 attenuated the effects of miR-522 in RA synovial fibroblasts.
Discussion
Due to the complexity in the pathogenesis of RA, RA usually developed with various manifestations and can be easily misdiagnosed. In addition, the existing criteria are unsuitable for the early detection in RA and cannot predict the aggravations of this disease. As a result, discovering new biomarkers for early detection is one of the challenges. Extracellular RNA consists mainly of rRNA, together with mRNAs, miRNA and small modified RNAs and oligonucleotide RNAs, and extracellular RNA in association with microparticles or exosomes, can be released by vital cells actively or cell death (Wissler, 2004).
Recently, extracellular RNA was found to participate in RA pathophysiology and RA synovial fibroblasts are influenced by extracellular RNA in a prodestructive manner (Zimmermann-Geller et al., 2016). Lots of studies have shown the aberrantly expressed miRNAs in the development of RA (Pauley et al., 2008; Nagata et al., 2009; Nakamachi et al., 2009), which indicated that the change of miRNAs expression level contributed to the mechanisms and may be suitable for predicting the RA course. On the contrary, targeting miRNA will enable a new medication toward RA treatment.
Studies have shown the implication of miR-522 in the cancer cell proliferation and drug resistance. MiR-522 is involved in promoting cell proliferation and cell cycle progression. miR-522 enhanced the level of cyclin D1 and Rb phosphorylation and inhibited cell cycle inhibitors expression, accelerating G1/S transition (Zhang et al., 2016). miR-522 also mediated glioblastoma cells proliferation by suppressing PH domain leucine-rich repeats protein phosphatase-1 (Zhang et al., 2015). Recently, miR-522 has been reported to reverse doxorubicin-induced drug resistance of colon cancer cell by targeting ABCB5 (Yang et al., 2015). Further study by Wu et al. (2017) demonstrated that some common miRNAs differently expressed in Kashin–Beck disease, osteoarthritis and RA and miR-522 were found to be upregulated in the blood specimens from the patients with RA.
The role of cytokines such as TNF-α and IL-1β has been well documented in RA, and miRNAs were found to regulate these cytokines release in RA synovial fibroblasts. For example, miR-26b inhibited the cell proliferation and cytokine secretion in human RA synovial fibroblasts via the Wnt/GSK-3 3β/β-catenin pathway (Sun et al., 2015); downregulation of miR-211 was reported to suppress the expression of proinflammatory cytokines and the chemokine in the RA synovial fibroblasts (Yang and Yang, 2015); and miR-573 overexpression suppressed the expression of IL-6 and cyclooxygenase 2 in RA synovial fibroblasts (Wang et al., 2016). In various inflammatory diseases, the MMPs were overexpressed and were shown to promote the progression of the disease (Itoh, 2015).
In the RA, RA synovial fibroblasts facilitated the productions of MMPs, which in turn promoted the cartilage destruction in the affected joints of RA patients (Araki and Mimura, 2017). However, the expression profile and functional details of miR-522 in the RA remain scant. In addition, it is unclear about the relationship among miR-522 and TNF-α, IL-1β, or MMPs, which were associated with synovial inflammation. In this study, the miR-522 expression levels in the synovial fibroblasts of patients with RA were significantly higher than that in control subjects. Meanwhile, markedly high levels of TNF-α and IL-1β were found in peripheral blood of patients with RA. Furthermore, miR-522 expression levels positively correlated with TNF-α and IL-1β expression.
In RA synovial fibroblasts, overexpression of miR-522 significantly increased the expression of TNF-α, IL-1β, and MMPs (MMP-1, MMP-3, and MMP-13) at both mRNA and protein levels. On the contrary, knockdown of miR-522 suppressed the TNF-α, IL-1β, and MMPs production stimulated by LPS. Therefore, the aforesaid results suggested that upregulation of miR-522 contributes to the enhanced expression of TNF-α, IL-1β, and MMPs in RA synovial fibroblasts.
A single miRNA can regulate multiple genes and how miR-522 regulated the expression of proinflammatory cytokines and MMPs in RA needs to be uncovered. SOCS3 was identified as one of target genes of miR-522 through TargetScan. In the aspect of SOCS3-involved in the regulation of RA, studies showed that overexpression of SOCS3 exhibited the anti-inflammatory property, which subsequently suppressed the induction and development of RA in mice (Shouda et al., 2001). Hematopoietic cell SOCS3 knockout mice showed significant elevation of IL-6 levels during experimental arthritis (Ushiki et al., 2016). In addition, silencing SOCS3 promoted IL-1-induced inflammatory joint disease (Carow and Rottenberg, 2014).
Our study has determined the regulatory role of miR-522 in the SOCS3 expression in RA synovial fibroblasts. miR-522 targets the 3′UTR region of SOCS3 gene and suppresses its expression at both protein and mRNA levels in RA synovial fibroblasts. Consequently, the higher level of miR-522 leads to a decreased expression of SOCS3. More importantly, the enforced expression of SOCS3 attenuated the enhanced effects of miR-522 on mRNA expression of TNF-α, IL-1β, MMP-1, MMP-3, and MMP-13.
In addition, overexpression of SOCS3 could inhibit the LPS-induced TNF-α in macrophages (Dai et al., 2013), and overexpression of SOCS3 suppressed the expression of TNF-α and IL-1β in the macrophages (Zhang et al., 2011). SOCS3 was also found to have inhibitory effects on the JAK-STAT pathway (Yoshimura et al., 2007) and MMPs expression could be induced via the activation of JAK-STAT signaling pathway (Li et al., 2001). In the chondrocytes, downregulation of SOCS3 enhanced leptin-induced expression of MMP-1, MMP-3, and MMP-13 (Koskinen-Kolasa et al., 2016), and SOCS3 overexpression caused a significant decreased of LPS-induced MMP-13 expression in osteoblasts (Gao et al., 2013). Thus, the effects of SOCS3 on the MMPs may involve the JAK-STAT pathway in the present study. Collectively, these results may suggest that miR-522-mediated expression of proinflammatory cytokines and MMPs is likely via regulating the SOCS3 in RA synovial fibroblasts.
In summary, our study for the first time assessed the effect of miR-522 and its target gene SOCS3 on the biological behaviors of RA synovial fibroblasts and elucidated the possible mechanism underlying RA progression. Our results suggested that the miR-522 regulated the expression of proinflammatory cytokines and MMPs partly via targeting SOCS3 in RA synovial fibroblasts, which may contribute to pathogenesis of RA. However, further investigations are required to fully understand the mechanism of miR-522 and its target genes in the pathogenesis of RA.
Footnotes
Acknowledgment
Thank you for the grant support by the Young Scholar Fund of Shaoxing People's Hospital (No. 2017B16).
Disclosure Statement
No competing financial interests exist.
References
Supplementary Material
Please find the following supplemental material available below.
For Open Access articles published under a Creative Commons License, all supplemental material carries the same license as the article it is associated with.
For non-Open Access articles published, all supplemental material carries a non-exclusive license, and permission requests for re-use of supplemental material or any part of supplemental material shall be sent directly to the copyright owner as specified in the copyright notice associated with the article.