
Abstract
Long noncoding RNAs (lncRNAs) were dysregulated in many kinds of cancers, including hepatocellular carcinoma (HCC). AK021443, as a novel lncRNA, was found to be upregulated in HCC, while its potential value and function are still unknown. The pathological changes of liver tissues were observed by hematoxylin and eosin staining. The expression levels of AK021443 in HCC tissues and cell lines were examined by quantitative real-time polymerase chain reaction (qRT-PCR). The proliferation ability of AK021443 on HepG2 and Bel-7402 cells was assessed by CCK8 and EdU staining assays. The role of AK021443 in HepG2 and Bel-7402 cell invasion and migration was measured by Transwell and wound healing assays. Finally, the expression of epithelial–mesenchymal transition (EMT) markers, including E-cadherin, N-cadherin, vimentin, and snail, was investigated by qRT-PCR, Western blot, and immunofluorescence. The role of AK021443 in vivo was also analyzed. Hepatoma cell nucleus increased in HCC tissues compared with normal liver tissues. AK021443 expression was increased in HCC tissues and cell lines. Knockdown of AK021443 significantly reduced HepG2 and Bel-7402 cell proliferation, invasion, and migration. Furthermore, inhibition of AK021443 in HepG2 and Bel-7402 cells significantly repressed EMT ability. Knockdown of AK021443 in vivo also significantly inhibited tumor growth with decreased Ki-67 levels and EMT phenotype in tumor tissues. However, these functions could be reversed by overexpression of AK021443. AK021443 significantly controlled HepG2 and Bel-7402 cell proliferation, colony formation, invasion, and migration by repressing EMT, which might provide a potential therapeutic target for HCC diagnosis.
Introduction
H
Recent evidence has suggested that long noncoding RNAs (lncRNAs) are implicated in developing HCC as key regulators. lncRNAs are important kinds of transcripts, which are longer than 200 nucleotides. A large investigation has revealed that lncRNAs participate in various biological processes, including cell proliferation, apoptosis, invasion, and migration, especially tumorigenesis and pathogenesis (Birgani et al., 2017; Zeng et al., 2017). The lncRNA RP1130-1 has been found to have low expression in patients with HCCs; it may be a powerful tumor biomarker of HCC (Xiao et al., 2016). Highly up-regulated in liver cancer (HULC) as a new lncRNA is upregulated in HCC tissues compared with normal liver tissues, the high level of HCC is associated with HBV infection status (Li et al., 2016). Downregulated HOXA-AS2 expression could significantly inhibit proliferation and invasion of SMMC-7721 and Bel-7402 cells, provided the candidate undergoes HCC patients' target therapy (Wang et al., 2016a). Furthermore, lncRNA BAIAP2-AS1 has been identified as a ceRNA, containing several miRNA binding sites, and functions as an miRNA sponge to promote HBV infection in HCC (Gong et al., 2016). lncRNA MALAT1 promotes HCC progression through regulation of ZEB1 expression by sponging miR-143-3p (Chen et al., 2017). lncRNA ROR is reported as a mediator of the chemotherapeutic response, the target therapy of lncRNA ROR could enhance chemosensitivity in HCC (Takahashi et al., 2014). Therefore, identification of HCC-involved lncRNAs and their functions might provide an effective strategy for diagnosis and prognosis of HCC patients. AK021443 is a novel lncRNA with an increasing expression significantly in advanced-stage HCC (Zhang et al., 2015), but the mechanisms of lncRNA AK021443 in HCC still need more study.
Epithelial–mesenchymal transition (EMT) is one of the most important cellular programs in tumor migration, invasion, and metastasis (Huang et al., 2017). During this process, tumor cells lose epithelial characteristics, E-cadherin, occludin, and other cellular tight junctions and transform into mesenchymal cells accompanying with increasing mesenchymal cell biomarkers, N-cadherin, vimentin, and snail. The transformed cells have been demonstrated to promote tumor cells to invade, metastasize, and in therapeutic resistance (Zhang et al., 2015). Growing evidences show that lncRNAs were involved in HCC invasion and metastasis by regulating EMT. lncRNA CPS1-IT1 served a tumor suppressor role by suppressing EMT in HCC carcinogenesis (Wang et al., 2016b). lncRNA AOC4P also exerted a tumor-suppressive effect on HCC tumor progression (Wang et al., 2015). lncTCF7 as an IL-6-inducible lncRNA exerted an important role in IL-6-induced EMT and invasion of HCC cells (Wu et al., 2015). Although several lncRNAs have been demonstrated in HCC tumor progression by regulating EMT, the specific roles of novel lncRNA AK021443 are not well studied.
Based on existing evidence and understanding, in this study, we assessed whether lncRNA AK021443 regulates EMT by influencing the proliferation and migration levels in HCC.
Materials and Methods
Clinical specimens
A total of 20 pairs of HCC and adjacent nontumor liver tissues were obtained from HCC patients undergoing surgery in the Affiliated Hospital of Hebei University (Hebei, China). This research was approved by the Ethics Committee of the People's Hospital of Hebei University. All the tissue specimens were collected and tested with informed consent of the patients.
Hematoxylin and eosin staining
The HCC and adjacent nontumor liver tissue samples were excised from normal or HCC patients after operation and fixed with 4% paraformaldehyde immediately for 24 h. Then, the fixed sample was washed and gradient dehydrated by different levels of ethanol. After hyalinization with xylene, the sample was embedded in paraffin overnight and sliced into 4-μm-thick consecutive sections. Sections were baked, deparaffinized, and hydrated. Eventually, they were stained with hematoxylin and eosin (HE) for histological observations.
Cell culture
The human hepatic carcinoma Bel-7402, Sk-Hep1, HepG2, Huh-7, and Bel-7404 cell lines and normal human liver cell line LO2 used in this study were purchased from the cell bank of Shanghai Institute for Biological Sciences. All the cell lines were cultured in complete high-glucose DMEM (Hyclone); Huh-7, Bel-7404, and LO2 cells were cultured in Eagle's minimum essential medium (Gibco) in 5% CO2 in a humidified incubator at 37°C, and the medium was supplemented with 10% FBS (Gibco) and 100 U/mL penicillin and 100 mg/mL streptomycin (Invitrogen) for cell culture.
Transfection
pcDNA3.1-AK021443 and its negative control (NC), si-AK021443, and NC siRNA (NC RNA) were purchased from GenePharma (Shanghai, China) and transfected into HepG2 and Bel-7402 cells using Lipofectamine 2000 reagent (Invitrogen); 2 × 105 HepG2 and Bel-7402 cells were seeded into six-well plates per well and incubated for 24 h. Then, HepG2 and Bel-7402 cells were transfected with pcDNA3.1-AK021443, AK021443 siRNA, or NC siRNA that was mixed with Lipofectamine 2000 transfection reagent.
Quantitative real-time polymerase chain reaction
Total RNA from tissues and cells was extracted using TRIzol reagent (Invitrogen). Real-time polymerase chain reaction (RT-PCR) was performed by DBI PrimeScript RT Reagent Kit (DBI Bioscience, Germany). Quantitative RT-PCR (qRT-PCR) was performed using DBI SYBR Premix Ex. The specific primers were as follows: AK021443 forward: 5′-CTTGAACCCAGAAGACAGG-3′ and reverse: 5′-ATGGAACATTAGAGGTAGCAC-3′; and β-actin forward: 5′-ATCGTGCGTGACATTAAGGAGAAG-3′ and reverse: 5′-AGGAAGGAAGGCTGGAAGAGTG-3′. The amplification profile was 94°C for 2 min, followed by 40 cycles of denaturation (94°C for 20 s, 58°C for 20 s, 72°C for 20 s). β-Actin was used as the reference gene. Relative quantification was determined using the 2−▵▵Ct method.
Western blot
Total protein samples harvested from HepG2 and Bel-7402 cells were prepared by RIPA buffer (Solarbio, China) and quantified by the BCA protein assay kit (Thermo Scientific). Protein lysates were separated by 10% polyacrylamide gels and transferred onto PVDF membranes (Millipore); the membranes were blocked with TBST containing 5% nonfat milk at room temperature for 2 h and then incubated with the antibodies (E-cadherin, N-cadherin, vimentin, snail, and GAPDH) at 4°C overnight. All the antibodies were purchased from Abcam (Cambrige, UK). After washing, the membranes were incubated with goat antirabbit horseradish peroxidase-conjugated antibody (BOSTER, China) for 2 h at room temperature. ECL system was used to visualize the signals of immune complexes (Millipore). GAPDH was used as an internal loading control.
CCK8 assay
First, 1 × 104 HepG2 and Bel-7402 cells were seeded into 96-well plates per well. Then, the cells were transfected as indicated by Lipofectamine 2000 transfection reagent. After incubation for 24, 48, and 72 h, respectively, cell growth viability was detected with a CCK8 Kit (Beyotime, China). The absorbance at 450 nm was recorded using a BioTek SYNFRGY4 microplate reader (Thermo Fisher Scientific).
EdU staining
After incubation of the transfected (as indicated) HepG2 and Bel-7402 cells for 48 h, cells were harvested and fixed with 70% alcohol overnight at 4°C, and then incubated with the EdU cell proliferation kit (RIBOBIO, China) according to the instructions; the fluorescence intensity of EdU+-formed compounds was detected by FACSCalibur (BD) at the excitation wavelength of 550 nm and emission wavelength of 565 nm.
Colony formation assay
Cell proliferation ability was also detected by colony formation assay. Eight hundred transfected HepG2 and Bel-7402 cells were seeded into a six-well plate per well. The medium was replaced with fresh medium every 3 days. After incubation for 21 days, cells were fixed with 4% paraformaldehyde and stained with 0.1% crystal violet solution (Sigma-Aldrich). The visible colonies were counted and photographed.
Transwell assay
The invasion abilities of HepG2 and Bel-7402 cells were assessed using Transwell cell culture inserts (Costar Corporation); 2 × 104 HepG2 and Bel-7402 cells transfected with NC or si-AK021443 were plated into each Transwell cell culture insert in serum-free DMEM, the lower chambers were placed with 700 μL of DMEM with 10% FBS. After incubation for 48 h, the cells in the upper surface of the upper chambers were removed with cotton swabs, and the inner migrated cells were fixed with 500 μL of 4% paraformaldehyde for 15 min. The membrane was washed with PBS and stained with crystal violet solution for 20 min. The migrated cells were counted and photographed by a microscope (Olympus, Japan).
Wound healing assay
Cell migration ability was measured by using scratch wound healing assay; after the HepG2 and Bel-7402 cells (transfected as indicated) were grown to 90% confluence in a six-well plate, a micropipette tip was used to scratch a wound. The cellular debris was washed with serum-free medium three times. The wound area was photographed under an Upright Metallurgical microscope (Olympus CX41) at 0 and 48 h.
Immunofluorescence
First, the transfected HepG2 and Bel-7402 cells were plated in a six-well tissue culture plate for 24 h. Next, the cells were fixed with 4% paraformaldehyde and incubated with E-cadherin, N-cadherin, vimentin, and snail overnight at 4°C. Then, cells were incubated with Goat Anti-Human IgG H&L (Alexa Fluor555) for 2 h at 37°C. All the antibodies were purchased from Abcam. Nuclei were counterstained with DAPI (0.5 μg/mL) for 15 min. Images of cells were obtained using an Olympus confocal microscope (Olympus).
Tumor model
First, human HepG2 cells and Bel-7402 cells were transfected with lentivirus vector of pcDNA3.1-AK021443 or shRNA-AK021443 and NC. Then, 3 × 106 HepG2 and Bel-7402 cells were subcutaneously injected in the rear flank of nude mice (male BALB/c-nu/nu mice, 6 per group), which were purchased from the Center of Laboratory Animal Science, Hubei Province, China. The tumor sizes were measured 3 days apart and the tumor volumes were calculated as follows: V (cm3) = width2 (cm2) * length (cm)/2. The animal study was approved by the institutional animal research committee of the Affiliated Hospital of Hebei University and animals were cared for following the guidelines for use and care of laboratory animals.
Immunohistochemistry
Immunohistochemistry of Ki-67 in tumor tissues was performed on 2-μm-thick formalin-fixed and paraffin-embedded specimen sections, which were incubated in xylene for 5 min, followed by 100% ethanol for 10 min, and 95% ethanol for 10 min. After antigen unmasking, the slides were blocked with 3% BSA for 30 min at room temperature. Then, the primary antibody for Ki-67 was incubated with the FFPE specimen sections at 4°C overnight, and then the second antibody was incubated for 2 h. The EnVision Detection System kit (DAKO, Denmark) was used for the DAB chromogen, followed by nuclear staining using hematoxylin.
Statistical analysis
Student's test was carried out for the analysis of data in this research. All the statistical tests were performed using GraphPad Prism 6. Values are expressed as the mean ± standard deviation. p < 0.05 was considered statistically significant. All the experiments were repeated three times at least.
Results
The level of AK021443 expression increased in human hepatoma tissues and cell lines
To investigate the potential clinical significance of AK021443 in HCC, firstly, HCC tissues and corresponding adjacent normal tissues were subjected to HE staining. As shown in Figure 1A, hepatoma cell nucleus increased greatly, the atypia and split image elevated in clinical HCC tissues compared with the adjacent normal liver tissues. Then, 20 pairs of clinical HCC tissues and corresponding adjacent normal tissues were collected to analyze the expression of AK021443 by qRT-PCR. As shown in Figure 1B, AK021443 expression increased significantly relative to that in adjacent normal tissues (p < 0.01). Additionally, AK021443 expression was also examined with qRT-PCR in the normal human liver cell line (LO2) and five HCC cell lines (Bel-7402, Sk-Hep1, HepG2, Huh-7, and Bel-7404); LO2 was used as a control cell line. Results indicated that AK021443 expression was upregulated in the HCC cell lines compared with LO2 (p < 0.01, Fig. 1C), especially with highest expression in HepG2 and Bel-7402 cells; therefore, further research were carried out in HepG2 and Bel-7402 cells. Taken together, these results demonstrated that AK021443 expression overexpressed in HCC tissue and suggested that AK021443 may play an important role in HCC.
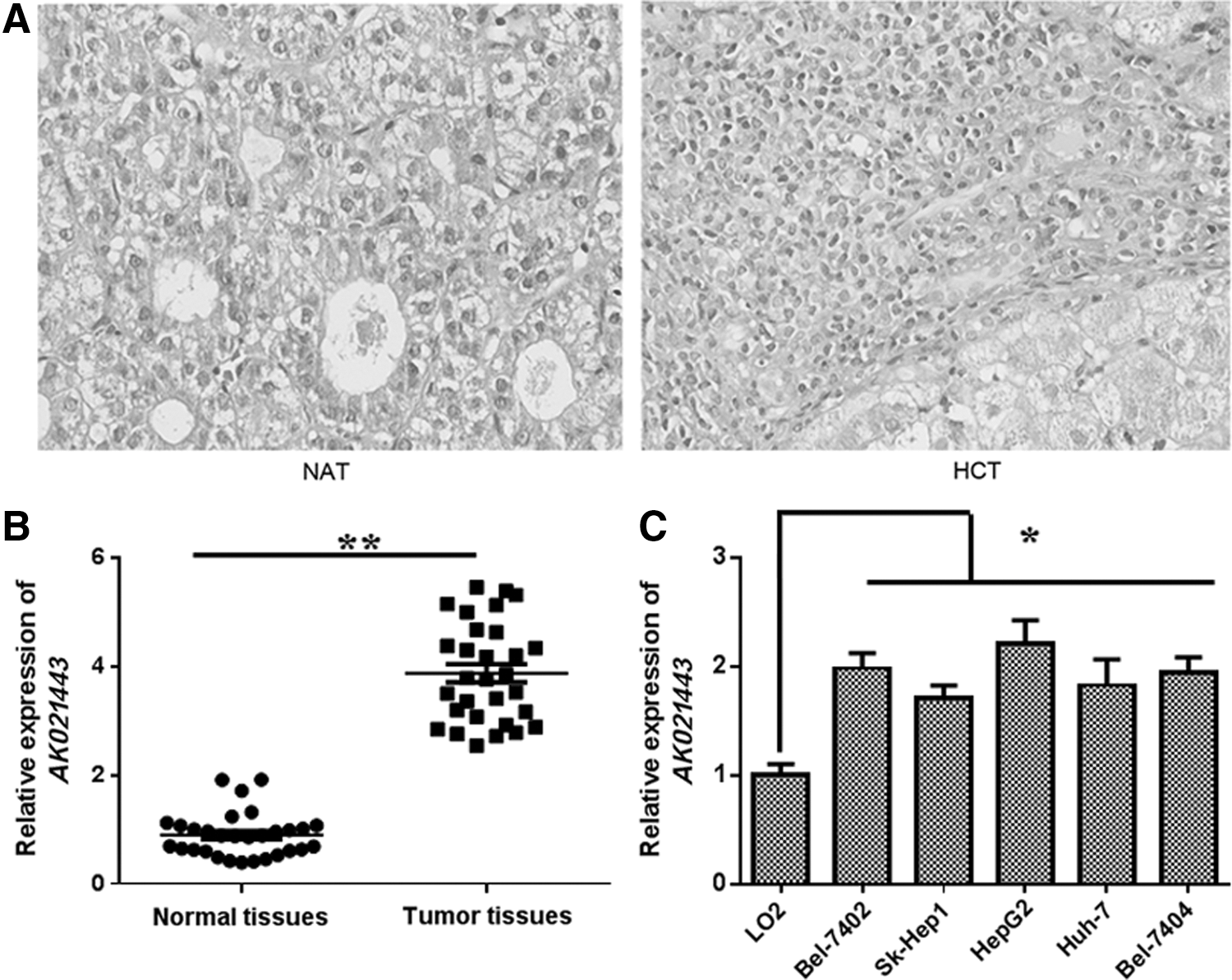
Expression of AK021443 in HCC tissues and HCC cell lines.
AK021443 siRNA suppressed proliferation, colony formation, invasion, and migration of HCC cells
To explore the biological function of AK021443 in the development process of HCC, first, HepG2 and Bel-7402 cells were transfected with AK021443 siRNA or the control. The results showed that AK021443 was significantly decreased in HepG2 and Bel-7402 cells transfected with AK021443 siRNA compared with the NC control (p < 0.01, Fig. 2A). Second, the function of AK021443 on cell ability of HCC was measured by CCK8 assays, as shown in Figure 2B; the downexpression of AK021443 significantly suppressed proliferation of HepG2 and Bel-7402 cells compared with the NC control (p < 0.05). EdU staining confirmed that inhibition of AK021443 resulted in fewer positive EdU+ cells compared with the NC group (Fig. 2C). To further investigate the function of AK021443, plate clone formation assay showed that HepG2 and Bel-7402 cells transfected with AK021443 siRNA had a significantly lower clone formation ability compared with cells transfected with the NC control (p < 0.01, Fig. 2D). Finally, we detected the effect of AK021443 on the ability of HCC cell invasion and migration by Transwell migration assay and wound healing assay, respectively. As shown in Figure 2E and F, the number of invasive and migratory HepG2 and Bel-7402 cells markedly decreased when cells were transfected with AK021443 siRNA (p < 0.01).
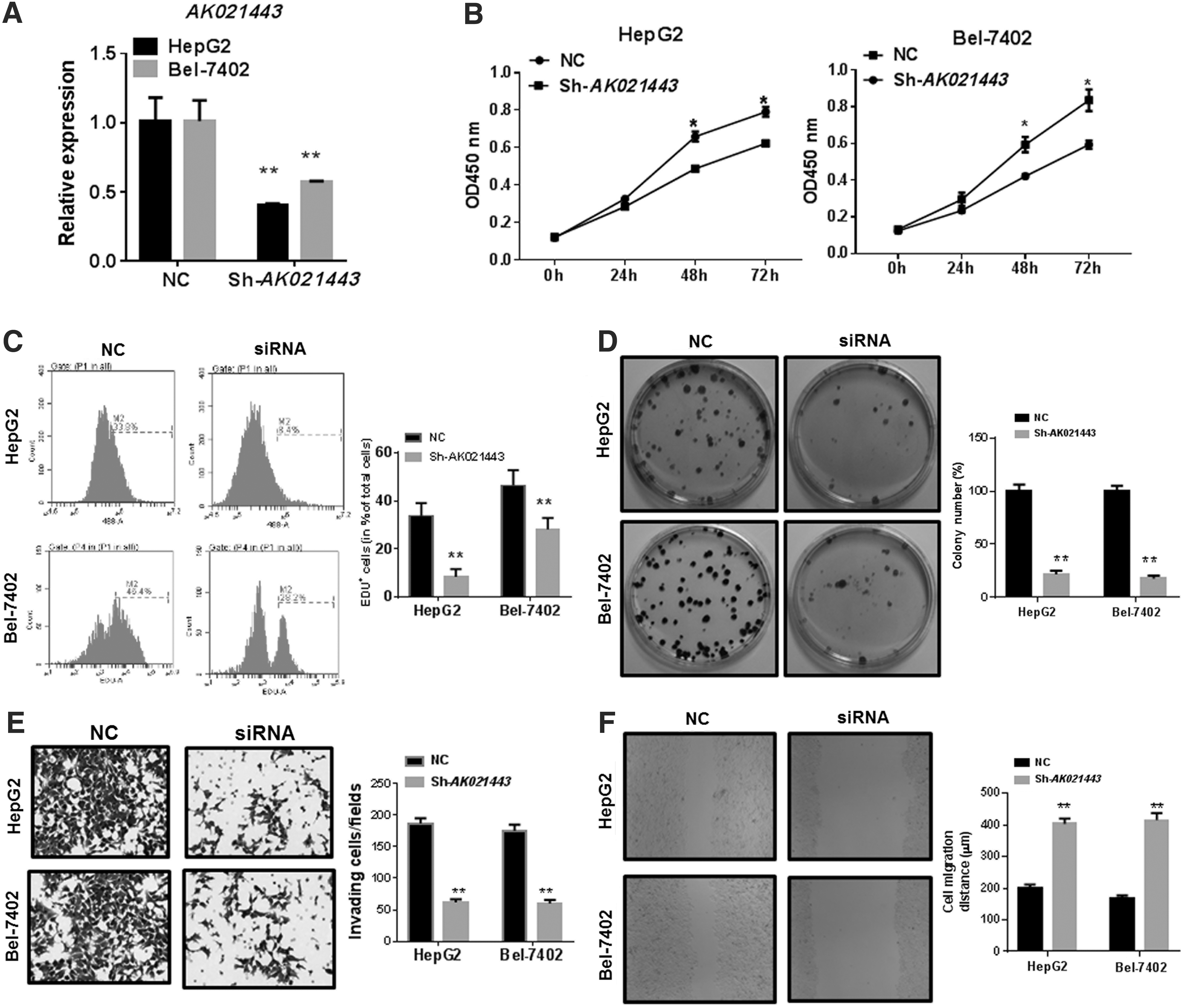
Effects of AK021443 inhibition suppressed the proliferation, colony formation, invasion and migration.
AK021443 siRNA regulated EMT-related proteins in HCC cells
According to previous reports, EMT plays an important role in the occurrence and development of induced HCC; therefore, we also detected whether AK021443 participates in the EMT process; qRT-PCR, Western blot, and immunofluorescence experiments were used to investigate the mRNA and protein expression levels of E-cadherin, N-cadherin, vimentin, and snail, which are the biomarkers of EMT. As shown in Figure 3A–E, the qRT-PCR and Western blot results indicated that mRNA and protein expression levels of E-cadherin increased, whereas N-cadherin, vimentin, and snail expression levels decreased in HepG2 and Bel-7402 cells transfected with AK021443 siRNA compared with the control (p < 0.01). Meanwhile, we also observed an increasing expression level of E-cadherin and a decreasing expression of N-cadherin, vimentin, and snail in HepG2 and Bel-7402 cells transfected with AK021443 siRNA (Fig. 3F). The result demonstrated that AK021443 may have an effect on EMT.
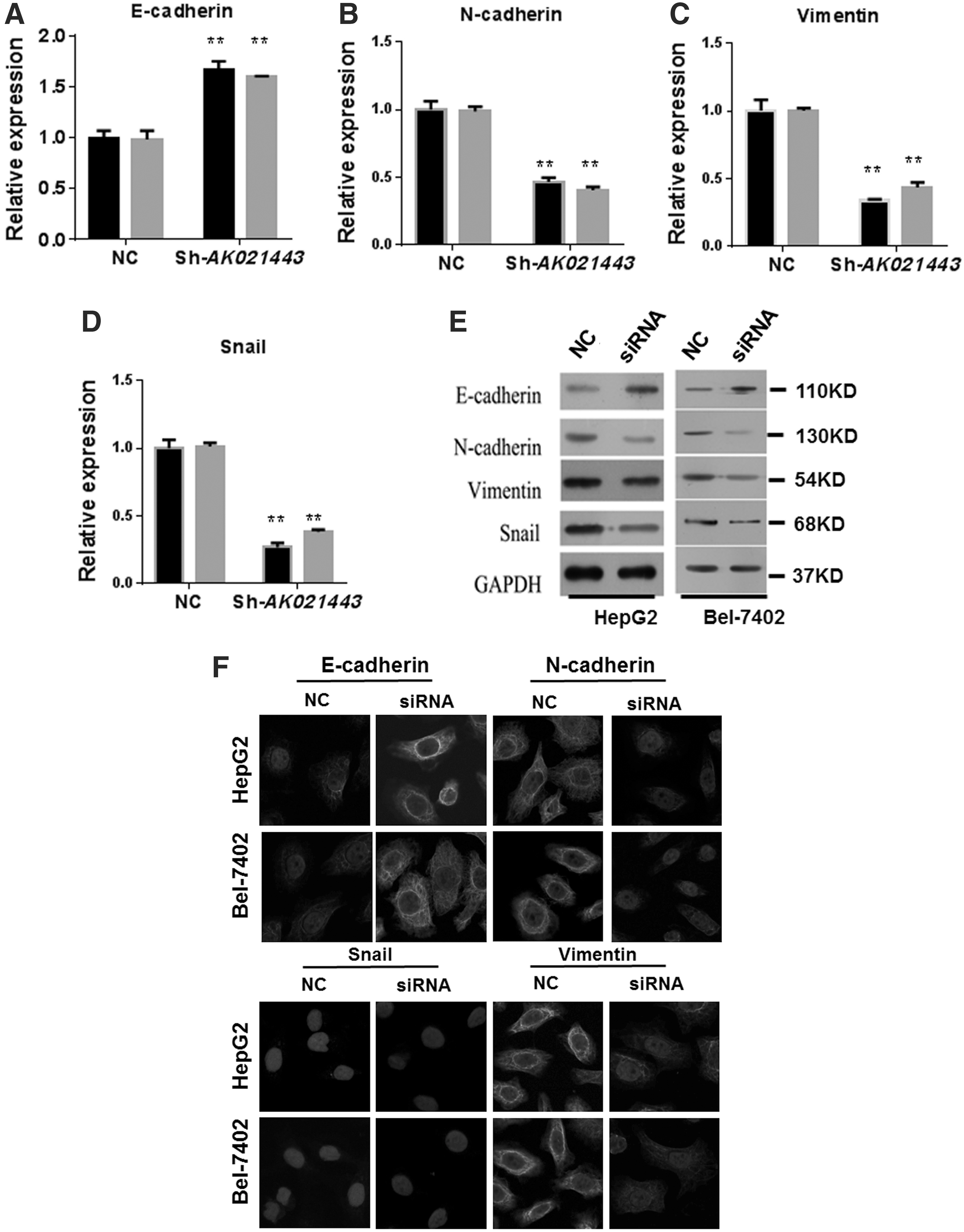
Inhibition of AK021443 regulated EMT-related proteins.
AK021443 overexpression promotes proliferation, colony formation, invasion, and migration of HCC cells
To further demonstrate the role of AK021443 in HCC, we performed the overexpression of AK021443 in two cell lines. We found that overexpression of AK021443 upregulated cell vitality, proliferation, clone formation, invasion, migration (Fig. 4), and the EMT phenotype (Fig. 5) in vitro.
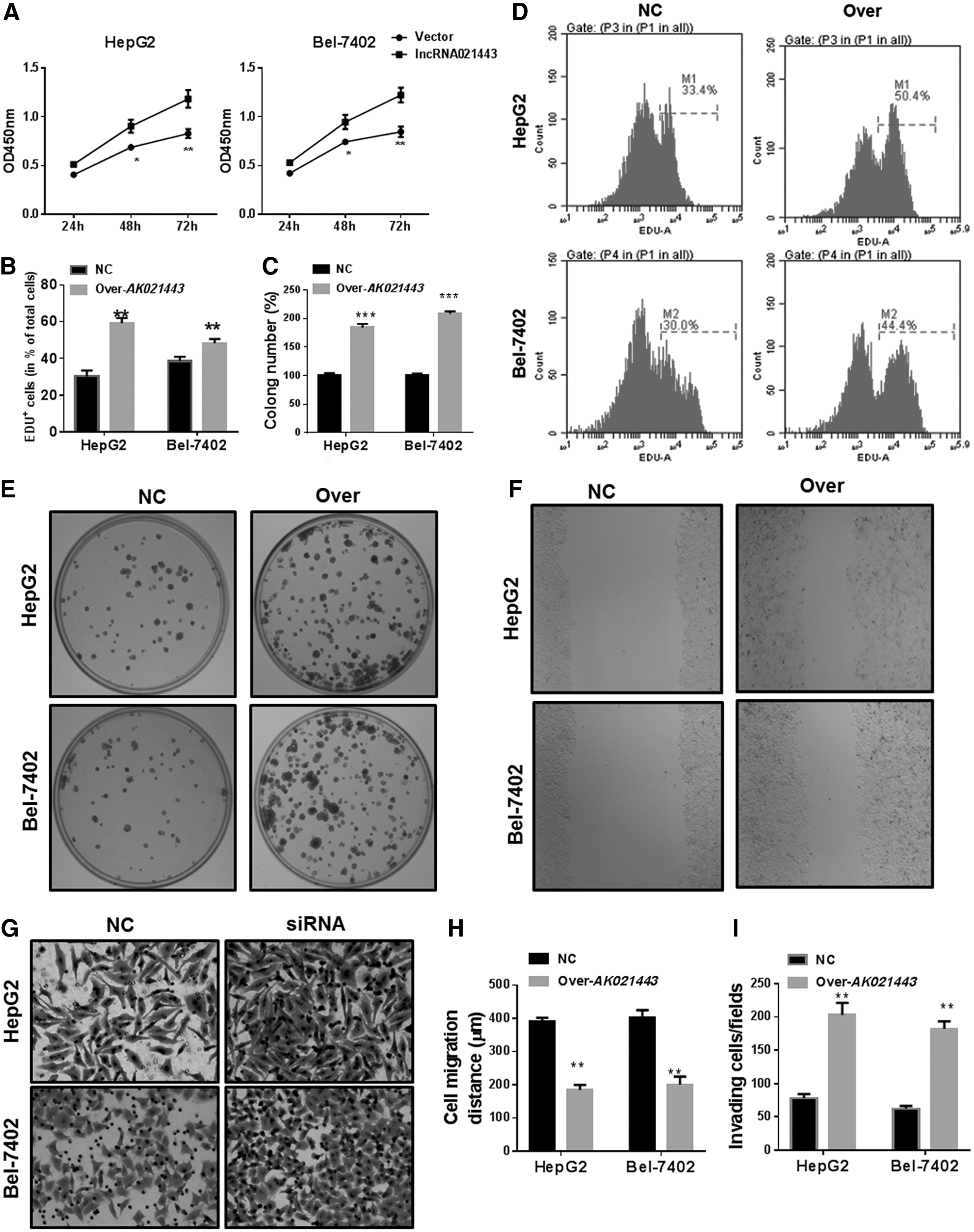
Effects of AK021443 overexpression promoted the proliferation, colony formation, invasion, and migration of HepG2 and Bel-7402 cells.
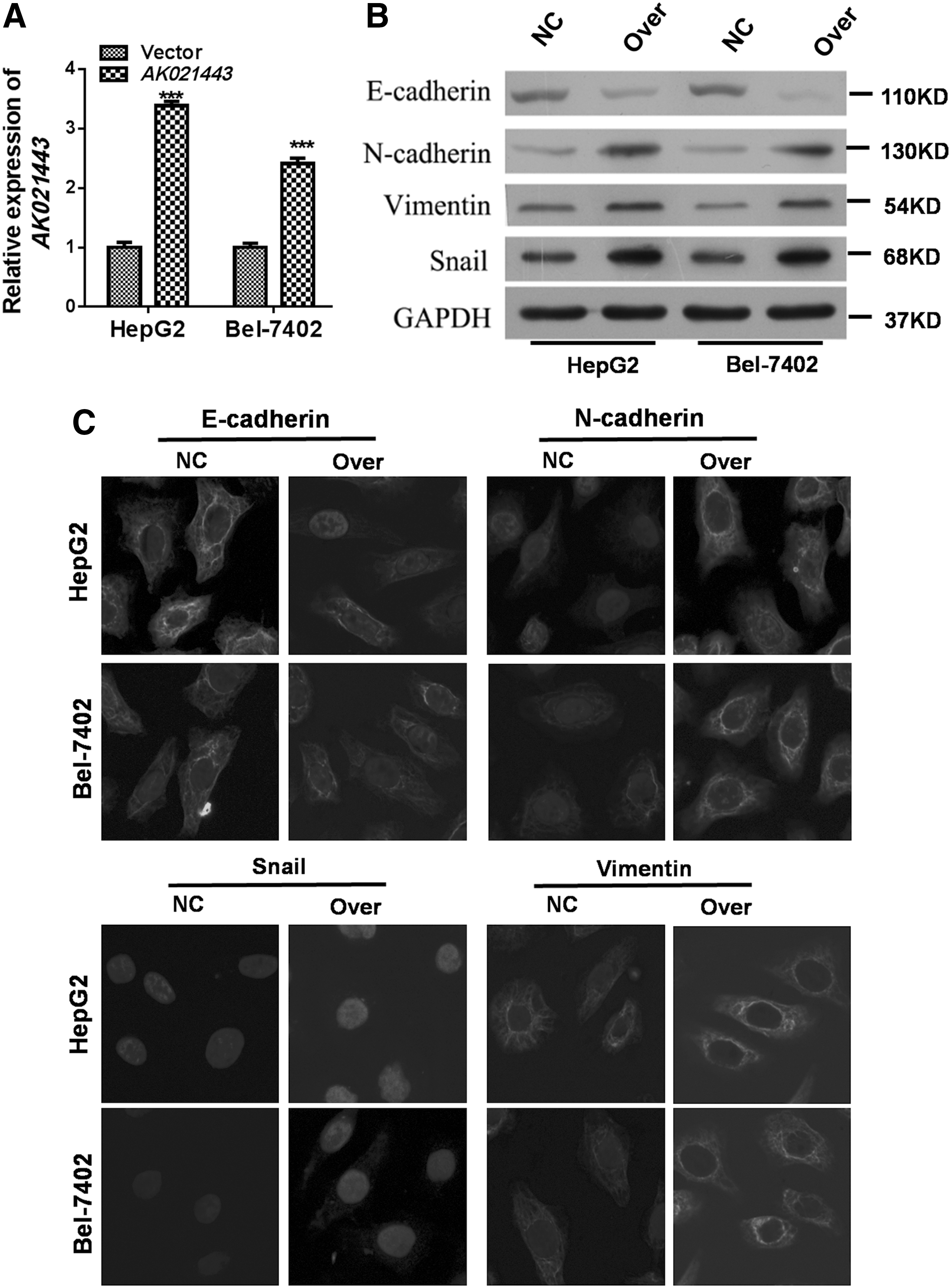
Overexpression of AK021443 regulated EMT-related proteins.
Knockdown of AK021443 restricts tumor growth with impaired EMT phenotype in vivo
To determine the tumorigenic role of AK021443 in vivo, the xenograft model of conditional human HepG2 cells and Bel-7402 cells with AK021443 overexpression or knockdown was established. The results indicated that AK021443 overexpression promoted tumor growth with a higher tumor volume (Fig. 6 A, B) and high Ki-67 levels (Fig. 6C) in tumor tissues than NC, and AK021443 knockdown showed the converse effects. In addition, the expression of AK021443 was confirmed in tumor tissues (Fig. 6D) and the EMT phenotype of tumor cells in vivo was estimated and it was found that overexpression of AK021443 promotes the formation of EMT, which was abrogated by inhibition of AK021443 (Fig. 6E). These effects were consistent with the findings in vitro.
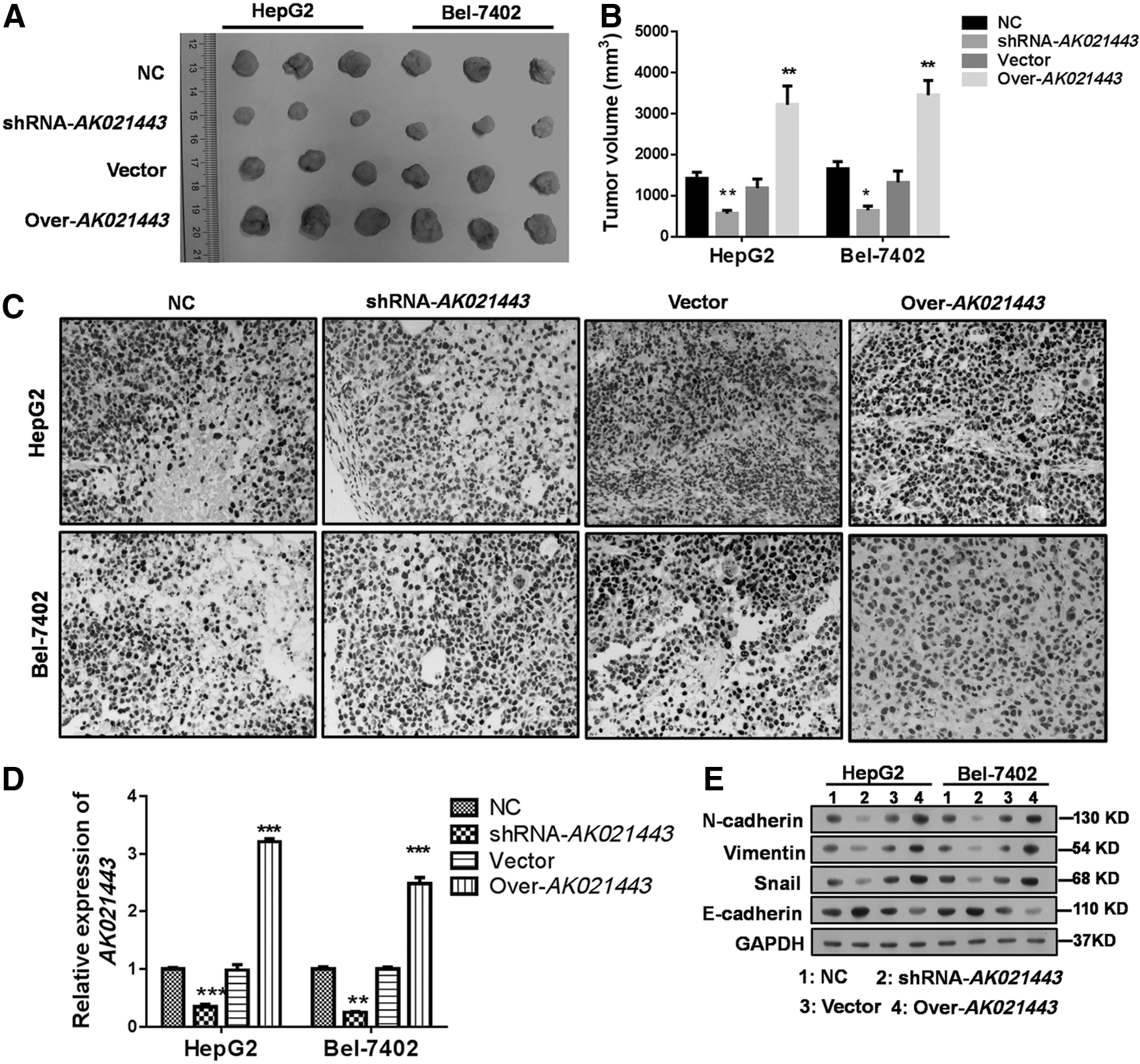
Inhibition of AK021443 restricts tumor growth in vivo.
Discussion
Due to the rapid infiltrating growth and high metastatic capabilities of HCC cells, primary HCC has a poor prognosis and high mortality (Saito et al., 2015; Roth et al., 2017). Therefore, it is of great importance to prevent and treat the migration and invasion of HCC cells for improving the diagnoses and treatment of HCC patients. AK021443 is a newfound lncRNA with an increasing expression significantly in advanced-stage HCC when compared with the nontumor liver tissue, and AK021443 was mainly found to be involved in cell cycle progression by gene ontology analysis (Zhang et al., 2015). In this study, 20 pairs of human clinical hepatoma tissues and five HCC cell lines were used to detect AK021443; we demonstrated that lncRNA AK021443 displayed higher endogenous expression in HCC tissues and HCC cells compared with the adjacent nontumor liver tissues and normal liver cells. Moreover, inhibition of AK021443 using siRNA could significantly reduce proliferation, colony formation, invasion, and migration capacities of HCC cells compared with normal liver cells, which were restored by overexpression of AK021443. Together, these results indicated that lncRNA AK021443 might function as a cancerigenic factor for HCC.
Invasion and metastasis are the primary causes of high mortality of cancer, and EMT is closely related to the high invasiveness and metastasis of cancer cells (Yuan et al., 2014). Epithelial cells lose junction proteins, such as E-cadherin and occludin, and obtain mesenchymal makers, such as N-cadherin, vimentin, and snail, which are crucial for promoting the EMT (Peng et al., 2017; Wang et al., 2017). There is mounting evidence that suggested that lncRNAs were involved in HCC invasion and metastasis by regulating EMT. Inhibition of lncRNA SNHG20 reduced cell proliferation and invasion; moreover, SNHG20 could regulate E-cadherin by binding to the enhancer of EZH2 (Liu et al., 2017). lncRNA UC001kfo could promote proliferation and metastasis by targeting a-SMA in HCC cells (Pan et al., 2017). lncRNA-Unigene56159, as a novel lncRNA, is overexpressed in HBV-related HCC tissues, and Unigene56159 greatly promoted cell migration and invasion in HCC by regulating EMT (Lv et al., 2016). High lncRNA-ATB expression promotes HCC cell invasion and metastasis by upregulating ZEB1 and ZEB2 expression, eventually reducing the EMT process (Yuan et al., 2014; Saito et al., 2015). In the present research, we first observed that AK021443 siRNA greatly inhibits the migration and invasion abilities of HepG2 and Bel-7402 cells. Furthermore, inhibition of AK021443 could significantly downregulate the EMT-promoting proteins (N-cadherin, vimentin, and snail) and upregulate the EMT-inhibiting protein such as E-cadherin in vitro and in vivo. However, this inhibition could be restored by the overexpression of AK021443. These results demonstrated that AK021443 may increase HCC invasion and migration by promoting EMT in HCC.
Conclusion
In summary, our research might provide a comprehensive understanding of AK021443 in HCC, and AK021443 exerts the important role of promoting proliferation, invasion, and migration by regulating EMT processes in HCC. Our findings provide new insights into the role of AK021443 in the development of HCC and it might be a potential therapeutic target for HCC diagnosis.
Footnotes
Disclosure Statement
No competing financial interests exist.