
Abstract
To date, in vitro culture systems able to sufficiently expand the small population of spermatogonial stem cells (SSCs), a tool for the development of sperm-mediated gene transfer techniques in transgenic pigs, in the porcine seminiferous tubule have not been reported. Therefore, as a step toward engineering a noncellular niche to support the in vitro maintenance of porcine SSC self-renewal, we investigated the types of integrin heterodimers that are expressed and functional on their membrane. The α and β integrin subunit protein expressions were analyzed using immunocytochemistry and fluorescence immunoassay, and the function of integrin heterodimers was confirmed by attachment and antibody inhibition assays. The integrin subunits, α3, α4, α5, α6, α8, α9, αV, and β1, were identified on the surface of them. Moreover, they showed significantly increased adhesion to fibronectin, laminin, and vitronectin, and functional blocking of integrin α4β1, α6β1, or αVβ1 significantly inhibited adhesion to these molecules. They showed significantly decreased adhesion to tenascin C and functional blocking of integrin α5β1 did not significantly inhibit adhesion to fibronectin. Accordingly, we confirmed that the integrin heterodimers α4β1, α6β1, and αVβ1 actively function on the surface of undifferentiated porcine SSCs, whereas α3, α5, α8, and α9 are present in inactive forms.
Introduction
R
There are several methods for generating transgenic animals: DNA microinjection (Niemann and Kues, 2000; Wheeler and Walters, 2001; Houdebine, 2002, 2009; Wheeler et al., 2003; Niemann et al., 2005), somatic cell nuclear transfer (Niemann and Kues, 2000; Wheeler and Walters, 2001; Houdebine, 2002; Wheeler et al., 2003; Niemann et al., 2005; Robl et al., 2007; Vajta and Callesen, 2012), and retrovirus- (Niemann and Kues, 2000; Houdebine, 2002, 2009; Wheeler et al., 2003; Niemann et al., 2005; Robl et al., 2007), biopolymer- (Wheeler et al., 2003; Nitta and Numata, 2013), and embryonic stem cell-mediated gene transfer (Niemann and Kues, 2000; Wheeler and Walters, 2001; Houdebine, 2002, 2009). However, these methods have shown low efficiency in the generation of transgenic progeny (Niemann and Kues, 2000; Houdebine, 2002, 2009; Niemann et al., 2005; Robl et al., 2007) and in transgene integration into the genome (Houdebine, 2002; Wheeler et al., 2003; Niemann et al., 2005). Therefore, sperm-mediated gene transfer has been considered (Maione et al., 1998; Smith and Spadafora, 2005; Lavitrano et al., 2006, 2013; Kurome et al., 2007; García-Vázquez et al., 2011). However, the extremely condensed chromosomes of the sperm head make it difficult to integrate foreign DNA into the genome (Lavitrano et al., 2006; Kurome et al., 2007; García-Vázquez et al., 2011). As a result, scientific interest in the generation of transgenic sperm using spermatogonial stem cells (SSCs) with easy transgene integration and sperm differentiation has gradually increased.
Despite a number of trials, development of an in vitro system capable of sufficiently expanding the small population of SSCs located in the seminiferous tubule of the porcine testis has not yet been successful. Previously, porcine SSCs have been cultured on a feeder cell-based cellular niche, which is difficult to manipulate, but cellular niche-dependent culture systems have not shown effective supports in maintaining self-renewal of porcine SSCs. Therefore, noncellular niches, which are easier to define and manipulate than cellular niches, have taken the spotlight and the development of noncellular niche-dependent culture systems mimicking the seminiferous tubule basement membrane (STBM) in direct contact with porcine SSCs in vivo has been requested.
The STBM, which plays an important role in maintaining the self-renewal of SSCs (Olie et al., 1995; de Rooij et al., 2008; Campos-Junior et al., 2012; Chen et al., 2013), is composed of fibronectin (Skinner et al., 1985; Oatley and Brinster, 2012), laminin (Siu and Cheng, 2004a, 2008; Mazaud Guittot et al., 2008; Cheng et al., 2010; Kanatsu-Shinohara and Shinohara, 2013), collagen IV (Enders et al., 1995; Siu and Cheng, 2004a, 2008; Mazaud Guittot et al., 2008; Cheng et al., 2010; Kim et al., 2011), entactin (Siu and Cheng, 2004b, 2008; Mazaud Guittot et al., 2008), and perlecan (Siu and Cheng, 2004b, 2008; Mazaud Guittot et al., 2008). Structurally, the presence of laminin in the STBM enhances the strength of the entire network by interacting with nidogen and perlecan (Kim et al., 2011), and collagen IV improves the stability by interacting with laminin (Enders et al., 1995; Siu and Cheng, 2004b). Moreover, laminin induces cellular responses by interacting with specific binding sites on membrane proteins embedded in laminin (Olie et al., 1995; Siu and Cheng, 2004b; Oatley and Brinster, 2012), and collagen IV influences adhesion, migration, and cell differentiation (Olie et al., 1995; Siu and Cheng, 2004b; Oatley and Brinster, 2012). The combined stimulation of signals derived from these extracellular matrix (ECM) proteins can regulate the destiny of SSCs through integrins that directly recognize the ECM proteins (Siu and Cheng, 2004b). Integrins are a group of transmembrane cell adhesion molecules constructed as a heterodimer of α and β subunits (Kim et al., 2011; Brizzi et al., 2012; Chen et al., 2013; Silván et al., 2013). A total of 18 α and 8 β subunits can be assembled together, generating at least 24 different heterodimers (Siu and Cheng, 2008; Kim et al., 2011; Brizzi et al., 2012; Silván et al., 2013). These assemblies maintain tissue integrity through cell-to-cell contact (Virtanen et al., 1997; Xi, 2009; Chen et al., 2013; Silván et al., 2013) and communicate extracellular signals to the cytoplasm (Ellis and Tanentzapf, 2010; Brizzi et al., 2012; Kanatsu-Shinohara and Shinohara, 2013; Silván et al., 2013). These heterodimeric receptors interact directly with binding sites in ECM proteins and induce numerous biological processes, including cell attachment (Virtanen et al., 1997; Siu and Cheng, 2004b; Ellis and Tanentzapf, 2010; Chen et al., 2013), spreading (Kim et al., 2011; Brizzi et al., 2012), proliferation (Olie et al., 1995; Ellis and Tanentzapf, 2010; Kim et al., 2011; Brizzi et al., 2012; Silván et al., 2013), survival (Xi, 2009; Ellis and Tanentzapf, 2010; Brizzi et al., 2012; Silván et al., 2013), morphogenesis (Xi, 2009; Geiger and Yamada, 2011), and differentiation (Virtanen et al., 1997; Xi, 2009; Ellis and Tanentzapf, 2010; Kim et al., 2011; Brizzi et al., 2012; Silván et al., 2013).
Despite knowledge of the ECM proteins that make up the STBM in the testis, the integrin family members expressed on the surface of SSCs have remained unknown in domestic animals, with the exception of rodents, making it difficult to define a noncellular niche for in vitro culture of SSCs derived from domestic animals. In this study, we identified the integrin heterodimers expressed on the plasma membrane of SSCs in pigs. The integrin subunits expressed in porcine SSCs were analyzed at the translational level and the functions of integrin heterodimer candidates based on the expressed subunits were identified by attachment and antibody inhibition assays.
Materials and Methods
Animals
One- to 4-day-old crossbred (Landrace × Yorkshire) or purebred (Yorkshire × Yorkshire) male neonatal piglets were generously supplied from Gumbo, Inc. (Wonju, Korea) and collection of testes from them were conducted through routine castration surgery. The Institutional Animal Care and Use Committee (IACUC) of Kangwon National University (IACUC approval No. KW-131106-1) approved all animal experimental procedures, which were performed according to the Animal Care and Use Guideline of Kangwon National University.
Harvest of SSCs from porcine testes
Male neonatal testes were transported from a local farm (Gumbo, Inc.) to our laboratory in ice-cold Dulbecco's phosphate-buffered saline (DPBS; Welgene, Inc., Daegu, Korea) supplemented with 1% (v/v) antibiotic–antimycotic solution (Welgene) within 1 h. Subsequently, to isolate testicular cells from testes, the tunica albuginea and epididymis were removed from testes and the seminiferous tubules were digested by 0.1% (w/v) type IV collagenase (Worthington Biochemical, Lakewood, NJ) in high glucose Dulbecco's modified Eagle's medium (DMEM; Welgene) at 37°C for 15 min. The fragmented seminiferous tubules were dissociated sequentially and separately by 0.1% (w/v) hyaluronidase (Sigma-Aldrich, St. Louis, MO) in high glucose DMEM and 0.25% trypsin-EDTA (Welgene) at 37°C for 10 min. The dissociated testicular cells were washed twice with DMEM containing 10% (v/v) heat-inactivated fetal bovine serum (FBS; Gibco, Carlsbad, CA) and filtered using a 70-μm nylon strainer (SPL, Pocheon, Korea) for eliminating myoid and Sertoli cells. Moreover, red blood cell lysis buffer (Sigma-Aldrich) was used for removing erythrocytes included in the dispersed cells. Then, SSCs were sorted from the isolated testicular cells by Petri dish plating postdifferential plating method described previously (Park et al., 2014). In brief, 5 × 106 testicular cells were resuspended in high glucose DMEM supplemented with 10% (v/v) heat-inactivated FBS and 1% (v/v) antibiotic–antimycotic solution and these cells were plated on 100-mm Petri dish (SPL) coated with 0.1% (w/v) gelatin (Sigma-Aldrich). After incubating for 16 h at 37°C, 1 × 106 cells suspended in medium were plated on 35-mm Petri dish in high glucose DMEM supplemented with 15% (v/v) heat-inactivated FBS, 0.1 mM β-mercaptoethanol (Gibco), 1% (v/v) nonessential amino acid (NEAA; Gibco), 2 mM
Immunocytochemistry
The SSCs derived from porcine testes were fixed with 4% (v/v) paraformaldehyde (Junsei Chemincal Co., Ltd., Chuo-ku, Japan) for 10 min and washed twice with DPBS. Subsequently, the fixed cells in DPBS were stained with fluorescent-unconjugated GFRα1 primary antibody at 4°C overnight and the detection of GFRα1 primary antibody was conducted by incubating Alexa Fluor® 546- or fluorescein isothiocyanate (FITC)-conjugated secondary antibodies diluted in DPBS at 4°C for 2 h. After rinsing twice with DPBS, the stained cells were double stained by FITC-conjugated anti-mouse integrin α4, PE-conjugated anti-mouse integrin α9, and fluorescent-unconjugated anti-human integrin α3, α5, α6, α8, αv, and β1 primary antibodies diluted in DPBS at 4°C overnight, and fluorescent-unconjugated anti-integrin primary antibodies were detected by incubating the double-stained cells with Alexa Flour® 488- or 546- or FITC-conjugated secondary antibodies at 4°C for 2 h. Table 1 describes the detailed information and dilution rate of the used antibodies. Then, rinsing twice with DPBS was performed and the double-stained cells were counterstained with mounting medium for fluorescence with DAPI (Vector Laboratories, Inc., Burlingame, CA). Finally, the triple-stained cells were monitored under a confocal laser scanning microscope (LSM880; ZEISS, Jena, Germany).
AIA, antibody inhibition assay; FI, fluorescence immunoassay; GFRα1, glial cell-derived neurotrophic factor family receptor alpha-1; ICC, immunocytochemistry; FITC, fluorescein isothiocyanate.
Fluorescence immunoassay
The fixation process was conducted by incubating 1 × 105 cells for 10 min in 4% (v/v) paraformaldehyde. After rinsing twice with DPBS, the fixed cells were stained for 2 h at 4°C with FITC-conjugated anti-mouse integrin α4, α5, α6, α9, αv, and β1 antibodies and fluorescent-unconjugated anti-human integrin α3 and α8 primary antibodies diluted in DPBS supplemented with 2% (v/v) FBS. Subsequently, primary antibodies were detected using Alexa Flour 488-conjugated secondary antibodies diluted in DPBS supplemented with 2% (v/v) FBS. The detailed information and dilution rate of the used antibodies are listed in Table 1. The stained cells were washed twice with DPBS and fluorescence intensity was measured using SoftMax® Pro 6.2.2. (Molecular Devices Corp., Sunnyvale, CA) after adding 100 μL DPBS to the stained cells.
Attachment assay
The attachment of SSCs to ECM ligands was conducted by somewhat modifying previous procedures (Lee et al., 2010). In brief, 96-well tissue culture plates were respectively coated with following concentrations of the purified ECM ligands: 0, 10, 20, 40, 60, and 80 μg/mL fibronectin (Millipore, Billerica, MA); 0, 200, 400, and 600 μg/mL laminin (Sigma-Aldrich); 0, 5, 10, and 20 μg/mL vitronectin (R&D Systems, Inc.); and 0, 1, 5, and 10 μg/mL tenascin C (R&D Systems, Inc.) overnight (minimum 18 h at 4°C). Each well was blocked with 1% (w/v) bovine serum albumin (BSA; Sigma-Aldrich) at 4°C for 1 h and then the wells were washed three times with DPBS. Around 1 × 105 porcine SSCs resuspended in SSC culture medium (pSSCCM) consisting of StemPro-34 medium (Invitrogen) supplemented with insulin–transferrin–selenium (Invitrogen), 60 μM putrescine dihydrochloride (Sigma-Aldrich), 6 mg/mL D-(+)-glucose (Wako Pure Chemical Industries, Ltd., Osaka, Japan), 0.11 mg/mL sodium pyruvate (Sigma-Aldrich), 1 μL/mL DL-lactic acid (Sigma-Aldrich), 5 mg/mL BSA (Sigma-Aldrich), 2 mM
Antibody inhibition assay
Each well of 96-well tissue culture plate was coated with 40 μg/mL fibronectin, 200 μg/mL laminin, and 5 μg/mL vitronectin overnight at 4°C. Blocking of the wells was conducted with 1% (w/v) BSA for 1 h at 4°C. Subsequently, for inhibiting function of integrin heterodimers, 1 × 105 SSCs of porcine neonatal testes were resuspended in pSSCCM, including anti-integrin α4 [9C10 (MFR4.B)], anti-integrin α5 [5H10-27 (MFR5)], anti-integrin α6 (NKI-GoH3), or anti-integrin αV (RMV-7) blocking antibody and incubated at 37°C for 2 h. The detailed information regarding the used antibodies is shown in Table 1. The functionally blocked porcine SSCs were then plated on each well and incubated at 37°C for 8 h. To remove nonadherent cells, the wells were washed extensively with DPBS. The adherent cells were fixed in 4% (v/v) paraformaldehyde for 10 min at room temperature and staining of adherent cells was performed with 0.1% (w/v) Crystal Violet in 20% (v/v) methanol for 5 min. Finally, the wells were washed twice with distilled water and supplemented with 50 μL 0.2% (v/v) Triton X-100 diluted with distilled water. The amount of dye was measured at 570 nm using a microplate reader (Epoch Microplate Spectrophotometer). Poly-
Statistical analyses
Statistical analyses of all numerical data derived from each experiment were conducted using the Statistical Analysis System (SAS) program. Furthermore, in detecting a significance of the main effects in the SAS package, control and treatment groups were compared by the PROC-GLM method. The p-values less than 0.05 was indicative of significant differences.
Results
Identification of integrin subunits expressed on the surface of undifferentiated porcine SSCs
To determine the integrin heterodimers expressed on the surface of undifferentiated porcine SSCs, expression of the individual α and β subunits that make up the integrin heterodimers interacting with each ECM protein observed in the testis (Skinner et al., 1985; Paranko et al., 1995; Ozturk et al., 2003; Siu and Cheng, 2004a, 2004b, 2008; Cheng et al., 2010; Oatley and Brinster, 2012) was monitored at the translational level. As shown in Figure 1, expression of the integrin subunits α5, α6, α9, αV, and β1 on the surface of undifferentiated porcine SSCs was observed, and the quantitative analysis (Fig. 2) showed that the integrin α5 subunit had the highest significant expression. Among the integrin subunits with significantly low expression, α9 and β1 showed intermediate expression compared with α6 and αV. The lowest significant expression was detected with α6 and the highest was found with αV. These results demonstrate that α5, α6, α9, αV, and β1 are localized on the membrane of porcine SSCs in an undifferentiated state.

The expression of the α and β integrin subunits expressed in undifferentiated porcine SSCs. The SSCs were sorted from the testicular cells enzymatically isolated from porcine testis by Petri dish plating post-DP method. Subsequently, the expression of integrin α and β subunit proteins in the sorted porcine SSCs was identified by immunocytochemistry. As the results, integrin α5
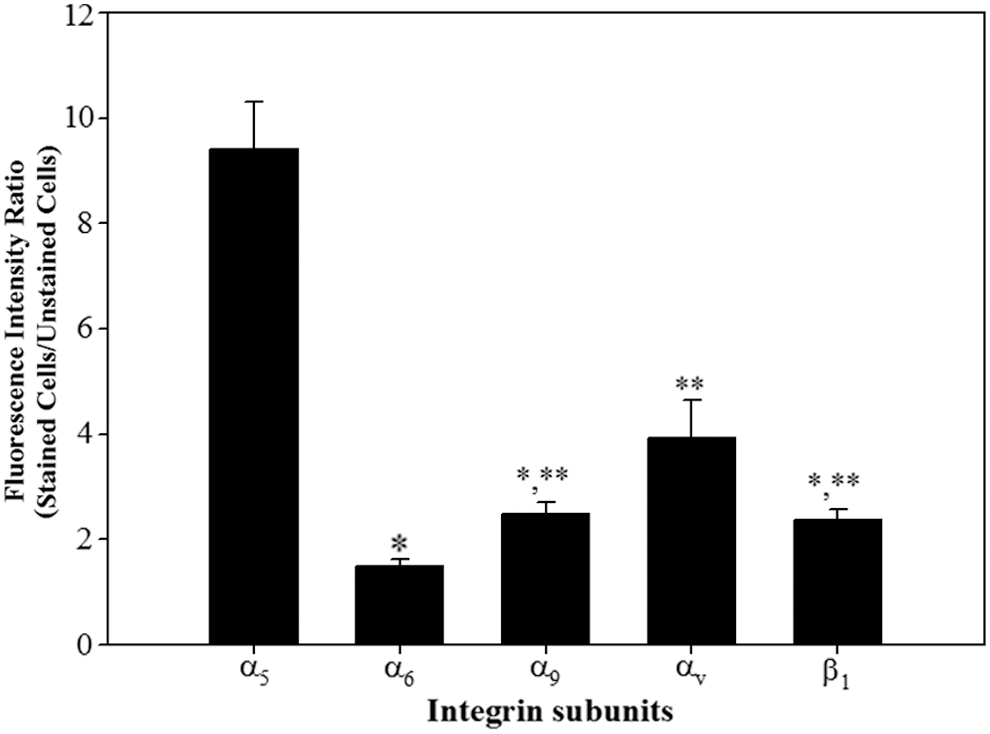
Quantitative analysis of the α and β integrin subunits expressed in undifferentiated porcine SSCs. The SSCs were sorted from the testicular cells enzymatically isolated from porcine testis by Petri dish plating post-DP method. Subsequently, the expression of integrin α and β subunit proteins in the sorted porcine SSCs was measured using fluorescence immunoassay, and then the expression level of each integrin subunit protein was calculated as the ratio of fluorescence intensity of stained cells to fluorescence intensity of unstained cells. The α5 integrin subunit showed the highest significant expression. Among the subunits with relatively weak expression, the lowest significant expression was detected with α6 and the highest significant expression was found with αV. Moreover, α9 and β1 showed intermediate expression. All data shown are mean ± SEM from three independent experiments. *,**p < 0.05. SEM, standard errors of the mean.
Identification of functional integrin heterodimers on the surface of undifferentiated porcine SSCs
Building off of these results, the previously described integrin heterodimers α5β1, α6β1, α9β1, and αVβ1 (Giebel et al., 1997; Shinohara et al., 1999; Siu and Cheng, 2004b, 2008; Ebata et al., 2005; Xi, 2009; Ellis and Tanentzapf, 2010) were hypothesized to be active integrin heterodimers expressed in undifferentiated porcine SSCs. Their presence was investigated by estimating the levels of adherent porcine SSCs cultured on natural ECM proteins interacting specifically with each integrin heterodimer and the level of adherence of porcine SSCs treated with antibodies blocking the function of individual integrins. Compared with those cultured without ECM proteins, porcine SSCs cultured on fibronectin, laminin, and vitronectin, but not those cultured on tenascin C, showed significantly improved adherence (Fig. 3). These results suggest that undifferentiated porcine SSCs express integrins α5β1, α6β1, and αVβ1, which specifically interact with fibronectin, laminin, and vitronectin, respectively, on the cell surface.

Identification of integrin heterodimers interacting with fibronectin, laminin, vitronectin, and tenascin C on the surface of undifferentiated porcine SSCs. Ninety-six-well tissue culture plates were coated with 0, 10, 20, 40, 60, and 80 μg/mL fibronectin; 0, 200, 400, and 600 μg/mL laminin; 0, 5, 10, and 20 μg/mL vitronectin; and 0, 1, 5, and 10 μg/mL tenascin C, respectively, and 1 × 105 porcine SSCs resuspended in pSSCCM were plated to each well. After incubation for 2 h at 37°C, adherent cells were stained with Crystal Violet and adherent level was quantified using a microplate reader. Porcine SSCs cultured on ECM plates with proteins showed significantly more attachment, regardless of protein concentration, than cultures without proteins. The highest significant rates of attachment were detected in cultures with 40 μg/mL fibronectin
Next, we blocked the functions of integrin heterodimers of undifferentiated porcine SSCs, and incubated the cells on 40 μg/mL fibronectin, 200 μg/mL laminin, and 5 μg/mL vitronectin, which showed the highest adherence of porcine SSCs (Fig. 3). SSCs in which integrins α6β1 or αVβ1 were blocked showed significantly less adherence, whereas those in which integrin α5β1 was blocked showed no significant decrease in adherence (Fig. 4). These results confirmed that undifferentiated porcine SSCs exhibit functional expression of the integrins α6β1 and αVβ1 on the cell surface, and that both α5 and α9 are present in inactive subunit forms and not as functional heterodimers.
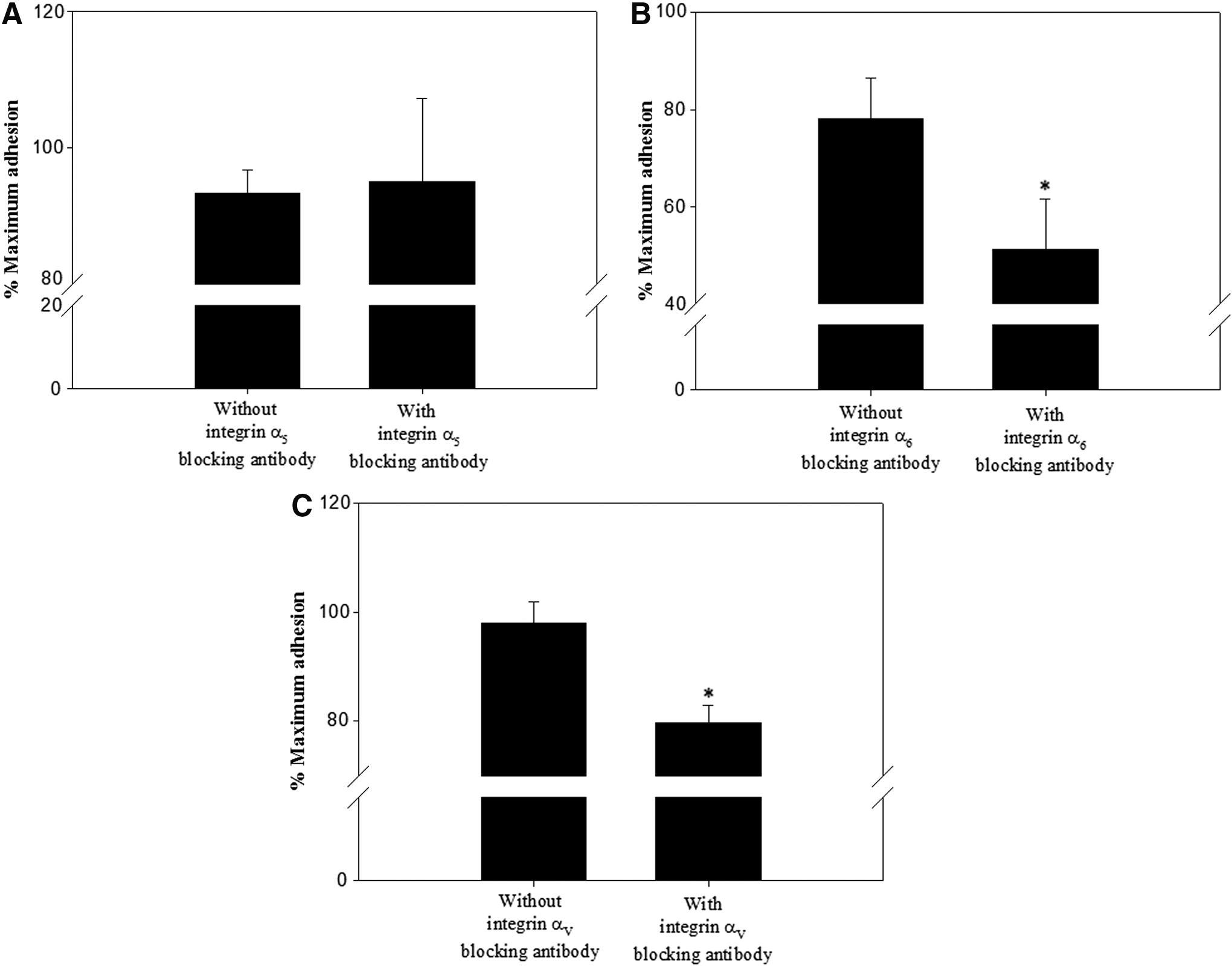
Functional analysis of the integrin heterodimers thought to function on the surface of undifferentiated porcine SSCs. Porcine SSCs incubated in the absence or presence of anti-integrin α5 [5H10-27 (MFR5)]
The adhesion of porcine SSCs to fibronectin in the absence of the functional integrin α5β1 (Fig. 3A) led us to hypothesize that porcine SSCs may express additional integrin heterodimers that bind fibronectin. Given that the integrins α3β1, α4β1, and α8β1 can bind fibronectin (Johansson et al., 1997; Aziz-Seible and Casey, 2011; Xu and Mosher, 2011), we assessed the expression of subunits α3, α4, and α8 on the membrane of undifferentiated porcine SSCs (Figs. 5 and 6). Each was expressed on the membrane, but the expressions of α3 and α4 were significantly higher than that of α8. Moreover, SSCs incubated on 40 μg/mL fibronectin with blockade with integrin α4β1 showed significantly lower adherence than those in which this integrin was not blocked (Fig. 7). These results confirmed the presence of integrin α4β1 on the membrane of undifferentiated porcine SSCs. In this way, we indirectly identified the presence of α3 and α8 as individual integrin subunits on the membrane of undifferentiated porcine SSCs.
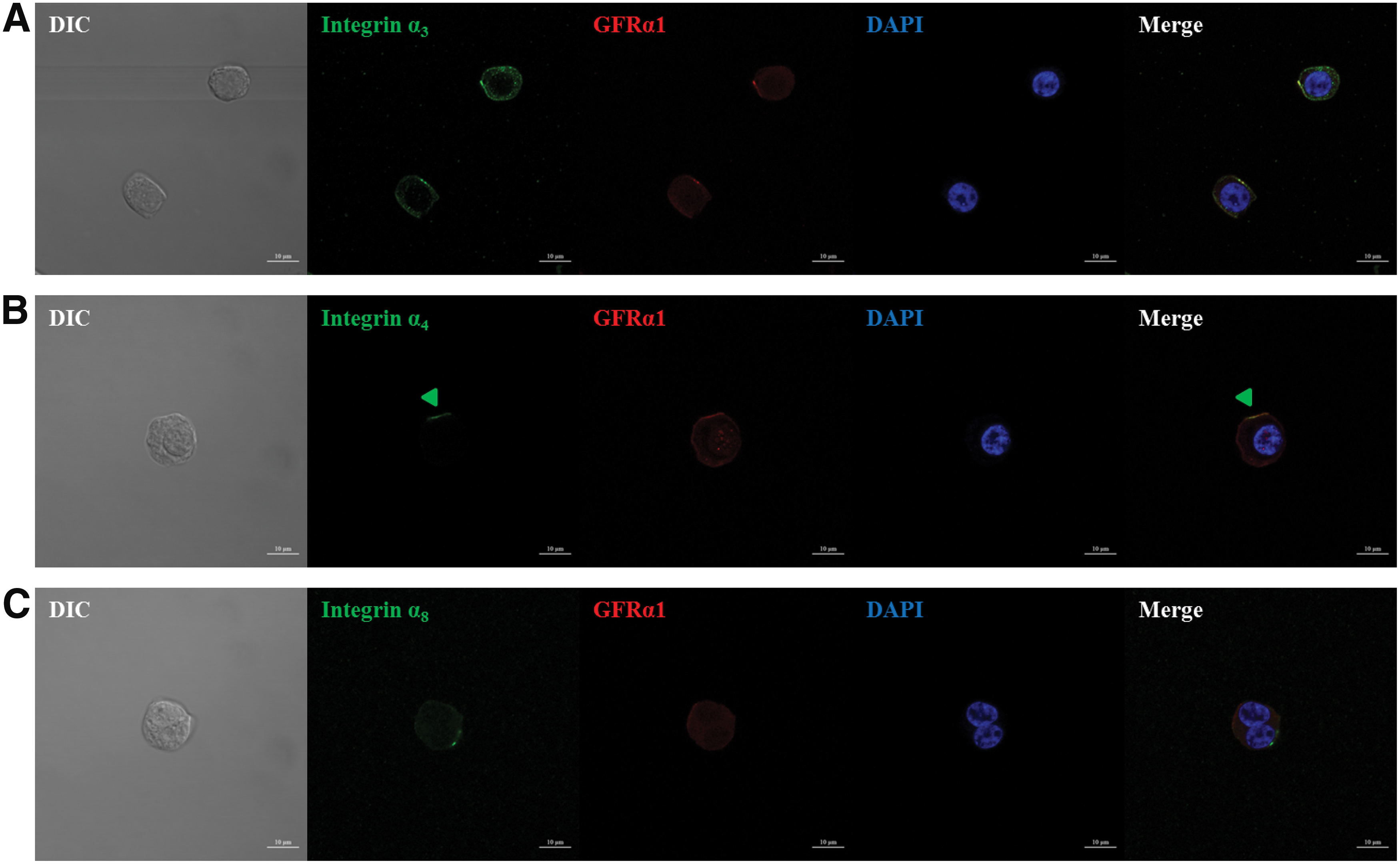
The expression of the integrin subunits α3, α4, and α8 in undifferentiated porcine SSCs. The isolation of SSCs from the testicular cells enzymatically isolated from porcine testis was conducted by Petri dish plating post-DP method. Then, the expression of integrin α3, α4, and α8 subunit proteins associated with binding domains within fibronectin in the sorted porcine SSCs was identified by immunocytochemistry. Integrin subunit α3
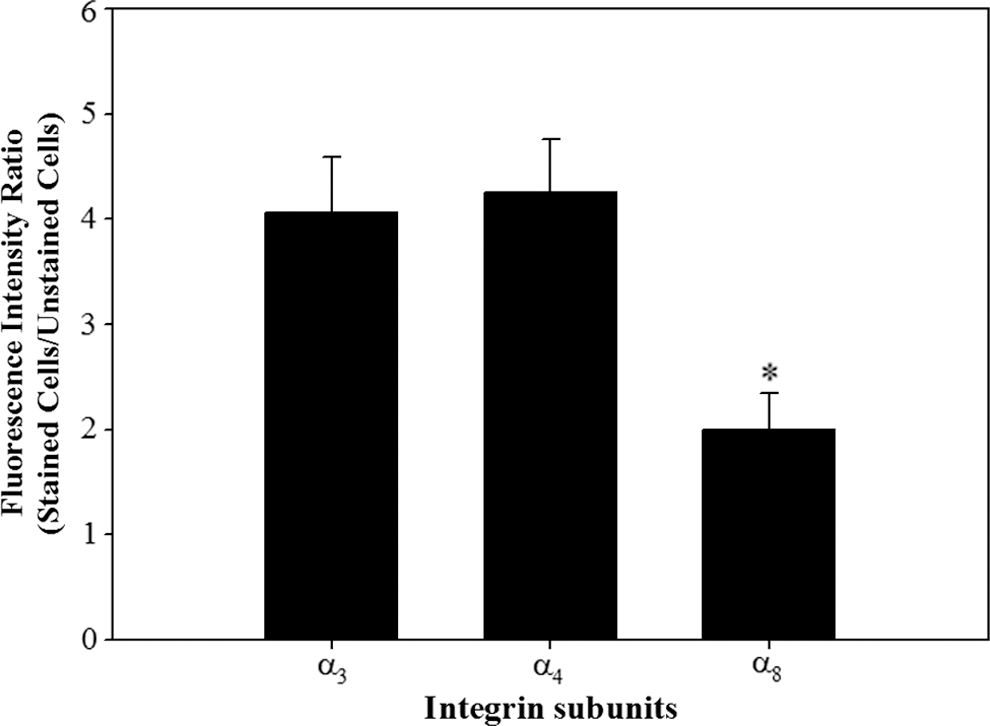
Quantitative analysis of the integrin subunits α3, α4, and α8 in undifferentiated porcine SSCs. The isolation of SSCs from the testicular cells enzymatically isolated from porcine testis was conducted by Petri dish plating post-DP method. Then, the expression of integrin α3, α4, and α8 subunit proteins associated with binding domains within fibronectin in the sorted porcine SSCs was measured using fluorescence immunoassay. Subsequently, the expression level of each integrin subunit protein was quantified as the ratio of fluorescence intensity of stained cells to fluorescence intensity of unstained cells. Integrin subunits α3, α4, and α8 were expressed on the surfaces of undifferentiated porcine SSCs. The expression of α8 was significantly lower than those of α3 and α4. All data shown are mean ± SEM from three independent experiments. *p < 0.05.
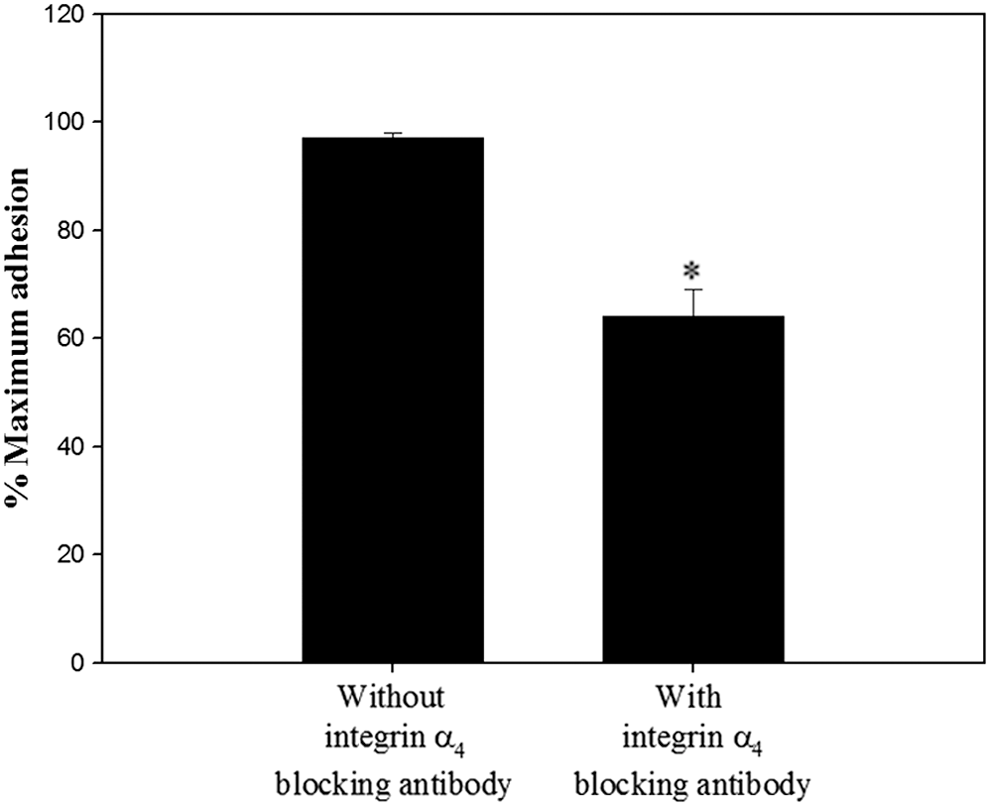
Functional analysis of integrin α4β1 on the surface of undifferentiated porcine SSCs. Porcine SSCs incubated in the absence or presence of anti-integrin α4 [9C10 (MFR4.B)] antibody was seeded on 40 μg/mL fibronectin-coated wells and incubated for 8 h in 37°C. After staining adherent cells with Crystal Violet, quantification of adherent level was conducted using a microplate reader. As the parameter of functional blocking by antibody, the percentage of maximum adhesion, which represents as the optical density of cells plated on 1 mg/mL poly-L-lysine-coated wells in the absence of any antibody, was demonstrated. Porcine SSCs treated with integrin α4β1 blocking antibody showed a significant decrease in attachment compared with SSCs not treated with blocking antibodies. All data are mean ± SEM from three independent experiments. *p < 0.05.
Discussion
To generate niches that will allow for self-renewal or differentiation of porcine SSCs, it is essential to understand the integrin family members that mediate intracytoplasmic transduction of signals derived from ECM proteins of the STBM in the seminiferous tubule. Therefore, we investigated the types of integrin heterodimers expressed on the membranes of undifferentiated SSCs in pigs. Through translational analysis of eight integrin subunits, followed by confirmation of their attachment to ECM proteins and inhibition with blocking antibodies, we found that the integrin heterodimers α4β1, α6β1, and αVβ1, and the integrin subunits α5 and α9 were expressed on the membranes. We also indirectly identified the localization of α3 and α8 on the membranes. These results suggest that the fibronectin-interacting integrin α4β1, the laminin-interacting integrin α6β1, and the vitronectin-interacting integrin αVβ1 may play an important role in the maintenance of porcine SSC self-renewal, and that fibronectin, laminin, and vitronectin analogs will likely be important for the development of niches customized to the maintenance of SSC self-renewal. In addition, we can speculate that the integrin subunits α3, α5, α8, or α9 may play a pivotal role in inducing early spermatogenesis of undifferentiated porcine SSCs.
The localizations of α3, α4, and α8 were determined on the surfaces of undifferentiated porcine SSCs (Figs. 5 and 6). The lack of any antibodies that can block α3 and α8 made it impossible to define the conformation of these subunits. However, weakened attachment of integrin α4β1 function-blocked porcine SSCs was observed on fibronectin (Fig. 7), indicating that integrin α4β1 functions as a heterodimer on the cell surface. This result also demonstrates that α3 and α8 may exist as inactive subunits on the surfaces of undifferentiated porcine SSCs.
In the seminiferous tubule microenvironment that generates spermatozoa from spermatogonium through spermatogenesis (Siu and Cheng, 2004a, 2004b, 2008; Cheng et al., 2010; Oatley and Brinster, 2012), inactive integrin subunits can induce cytological, chromosomal, and morphological changes in the spermatogonium following detachment of SSCs from the STBM. In this study, the subunits, α3, α5, and α8, were observed in inactive forms on cell membranes, indicating that signals derived from the interaction between fibronectin and integrin heterodimers with subunits α3, α5, and α8 may be important in the differentiation of SSCs into sperm; this was supported by reports that fibronectin is expressed in spermatogenic cells in the seminiferous tubule (Fusi and Bronson, 1992; Schaller et al., 1993). Furthermore, the presence of the inactive integrin subunit α9 on cell membrane demonstrates that signals derived from the interaction between tenascin C and α9-containing heterodimers may facilitate the movement of SSCs into the adluminal compartment of the seminiferous tubule during spermatogenesis, which is supported by the fact that tenascin C regulates cell migration (Deryugina and Bourdon, 1996; Zagzag et al., 2002). Therefore, knowledge of the inactive integrin subunits expressed on the membranes of specific cells is important in the construction of microenvironments customized for a particular cell fate.
The integrins α4β1 and α6β1 were observed on the surfaces of undifferentiated SSCs (Figs. 3 –7), suggesting that the presence of fibronectin and laminin in the STBM may be important for maintaining self-renewal in porcine SSCs. Despite the absence of vitronectin in the STBM, the integrin αVβ1 was expressed on the surfaces of undifferentiated SSCs (Figs. 3 and 4). Therefore, we can speculate that αVβ1 may help maintain self-renewal through an interaction with vitronectin in the ECM of neighboring undifferentiated SSCs or spermatogenesis through an interaction with vitronectin in the ECM of neighboring supporting cells in the seminiferous tubules. There have been no reports of the expression of vitronectin in undifferentiated SSCs, although its expression has been reported in spermatogenic cells of adult testes during normal spermatogenesis (Sawada et al., 1996). The integrin αVβ1 may contribute to spermatogenesis of porcine SSCs, although further studies are required to validate this.
Translation and functional screening demonstrated that the integrins α4β1, α6β1, and αVβ1 are expressed on the surfaces of undifferentiated porcine SSCs. Expression of the inactive integrin subunits α3, α5, α8, or α9 was also demonstrated. Identification of these integrins will be useful for determining the identity of undifferentiated SSCs in pigs, and for generating synthetic niches appropriate for the maintenance of SSCs in the undifferentiated state or to induce their differentiation into sperm. Moreover, these achievements will greatly improve the applicability of synthetic niches with respect to the regulation of SSC destiny in pigs.
Footnotes
Acknowledgments
This work was supported by the Korea Institute of Planning and Evaluation for Technology in Food, Agriculture, Forestry, and Fisheries (IPET) through Agri-Bioindustry Technology Development Program, funded by the Ministry of Agriculture, Food, and Rural Affairs (MAFRA) under Grant IPET117042-3 and IPET112015-4.
Disclosure Statement
No competing financial interests exist.