
Abstract
Phospholipase A2-IVA (PLA2G4A) is the most abundant subtype of cytoplasmic phospholipase A2 (cPLA2) and is an important enzyme in tumor development. Our study aimed to explore the role of PLA2G4A in the regulation of lung cancer. The contents of cell-related cytokines (microsomal prostaglandin E synthase-1 [mPGES], PGE2, and prostacyclin [PGI2]) in A549 cells were analyzed by ELISA kits. Cell counting kit-8 (CCK8) was used to detect the effects of inhibitor of cPLA2 (arachidonyl trifluoromethyl ketone
Introduction
L
Human alveolar epithelial cell line A549 can fix or hang a solution in vitro. The characteristic of these cells is that they are able to synthesize lecithin, which is important for the maintenance of cell membrane phospholipids (Wang et al., 2016). A549 cells are widely used in in vitro model of drug metabolism and are also used as a transfection host (Wang et al., 2017).
Phospholipase A2 (PLA2), secreted and released by activated monocytes, macrophages, and neutrophils, is a large class of enzymes that can hydrolyze the structure of the Sn-2 position lipid bond (Liao et al., 2011). There are a variety of effects for PLA2, such as the production of inflammatory mediators and involvement of cell signaling (Foldbjerg et al., 2011). Especially, the activity of cytoplasmic PLA2 (cPLA2) was increased in the acute or chronic inflammation of the gastrointestinal system and is activated by many inflammatory mediators (Kasurinen et al., 2017). IVA PLA2 (PLA2G4A) is the most abundant subtype of cPLA2, and its elevated expression is associated with tumor invasion and metastasis (Xing et al., 2011). Studies have found that PLA2G4A expression is elevated in early lung cancer tissues compared with normal lung tissue.
Cyclooxygenase (COX) includes two isozyme isomers, namely COX-1 and COX-2. COX-1 is an inherent housekeeping enzyme, which catalyzes the production of physiological needs of prostaglandin E (PGE), regulates peripheral vascular resistance, protects the gastric mucosa, and regulates platelet aggregation (Slomiany and Slomiany, 2009). COX-2 is called inducible enzyme, which is rapidly expressed by mononuclear macrophages and fibroblasts after the organism is stimulated by inflammatory factors. It can promote inflammatory response and cause tissue damage (Prima et al., 2017). The levels of COX were increased in lung cancer, and the COX inhibitors could produce a strong antagonistic effect on the growth inhibition of lung cancer cells (Yan et al., 2013). Our study aimed to explore the role of PLA2G4A in the regulation of lung cancer cells, to search new target for the treatment of lung cancer.
Materials and Methods
Cell culture and grouping
Human lung carcinoma epithelial cells (A549) were purchased from American Type Culture Collection (Rockville, MD) and cultured in RPMI-1640 medium with 10% fetal bovine serum, 2 mM glutamine, and 1 mM pyruvate (Beijing Solarbio Science & Technology Co., Ltd., China) at 37°C and 5% CO2. The cells in logarithmic phase were collected and used for subsequent experiments. The cells were treated with 50 μM arachidonyl trifluoromethyl ketone (AACOCF3), the inhibitor of cPLA2 (Calbiochem, San Diego, CA), meanwhile the control group cells were treated with 30 μL phosphate-buffered saline (PBS).
Detection of cell proliferation by cell counting kit-8
A549 cells (2 × 103 cells/well) were inoculated into 96-well plate, then incubated at 37°C and 5% CO2 incubator overnight, and then incubated for 24 h with 50 μM AACOCF3. Then 10 μL cell counting kit-8 (CCK8) solution was added per well and incubated for 1 h at 37°C and 5% CO2 incubator, subsequently the absorbance was measured at the wavelength of 450 nm used an enzyme labeling instrument (SpectraMax 190; Eppendorf, Hamburg, Germany). Cell inhibition rate = (ODcontrol group − ODAACOCF3 group)/(ODcontrol group − ODblank group) × 100%.
Scratch wound healing assay
A549 cells were harvested with 0.25% trypsin–EDTA solution (Sigma). The cells were resuspended in culture medium, then 5 × 105 cells/well were seeded in six-well plates. When the cells were fully fused, the cells were scratched at the bottom of the culture plate with 10 μL pipette tip. PBS was used to wash the plates for two times to remove floating cells. Serum-free medium was then added. The plates were placed at 37°C and 5% CO2 environment. Photographs of the plates were taken at 0 and 24 h with inverted microscope (Olympus, Japan).
Transwell assay
The precooled RPMI 1640 culture medium and Matrigel (Beijing Solarbio Science & Technology Co., Ltd.) were mixed in the ratio 1:1. Then 100 μL matrigel solution was added per well and evenly spread in the bottom of the upper chamber (Thermo, Germany) then incubated at 37°C incubator. About 200 μL transfected cells suspension (5 × 104 cells) was added per well. In the lower chamber, 500 μL RPMI-l640 medium containing 10% fetal bovine serum was added and incubated at 37°C incubator for 24 h. Then, the chambers were washed with PBS for two times and fixed with 1 mL methanol for 30 min. The invasive cells were stained with 0.5% crystal violet (Beijing Solarbio Science & Technology Co., Ltd.) for 30 min, then washed with PBS for two times. Under the inverted microscope, five visual fields were randomly selected to calculate the number of invasion cells.
ELISA
The contents of cell factor microsomal prostaglandin E synthase-1 (mPGES) (ABIN491828; Cloud-Clone, Houston), PGE2 (ABIN5592019; Signalway, MD), and prostacyclin (PGI2) (ABIN1980586; Houston) were detected by the corresponding kits in strict accordance with the manufacturer's description.
Detection of expression level of related mRNA by real-time quantitative PCR
Total RNA was extracted by TRIzol Kit (Invitrogen) and the purity of RNA was qualified by OD260/OD280. SuperScript III Reverse Transcriptase (Thermo Fisher Scientific) was used to reverse cDNA. Master EP realplex2 (Eppendorf) was used to conduct real-time quantitative PCR (conditions: 94°C for 5 min, 94°C for 60 s, 53°C for 45 s, 35 cycles). The relative expression of mRNA was calculated by the 2−ΔΔCt method (Livak and Schmittgen, 2001), and the β-actin mRNA was used for a reference to calculate the relative expression levels. Primer sequences used in this study are as follows: PLA2G4A, forward: 5′-GCCAAGTGACTCTAGTCCTCC-3′ and reverse: 5′-AGACATAGCACTCCTTCAGCC-3′; COX-2, forward: 5′-CCCACTTCAAGGGATTTT-3′ and reverse: 5′-CCAGACCAAAGACCTCCT-3′; β-actin, forward: 5′-CGGAGTGAGCGATCTTACAGG-3′ and reverse: 5′-TCATCAGCGACTCTGACCACA-3′.
Western blot
After washing with PBS, cells were lysed by sonication, centrifuged at 18000 g for 30 min at 4°C. The protein concentration was measured by the BCA Kit (Beijing Solarbio Science & Technology Co., Ltd.). A total of 4 μL protein samples were added to the 5 × buffer solution, then added to 10% SDS-PAGE electrophoresis (Mini-Protean-3 type; Bio-Rad, Hercules, CA). Protein samples were transferred to polyvinylidene fluoride membrane (Merck, Darmstadt, Germany). The membrane was then blocked with Tris-Buffered Saline and Tween 20 solution containing 5% nonfat dry milk for 1 h. The rabbit anti-mPGES antibody (1:100, ABIN1860349; Cloud-Clone), anti-COX-1 antibody (1:1000, ABIN3180550, antibodies-online; Aachen, Germany), and anti-COX-2 antibody (1:15,000, ABIN272477; Oxford Biomedical Research, Oxford, United Kingdom) were used as primary antibodies. The anti β-actin antibody (1:1000, ab8226; Abcam, Cambridge, England) was internal reference. After incubating the membranes at 4°C overnight and after washing three times, the membranes were incubated with goat anti-rabbit IgG-HRP (1:1000, ab16284; Abcam) for 1 h. The results were observed and recorded by Roche Elecsys-2010 chemiluminescence (Switzerland). Protein expression levels were normalized according to the β-actin and quantified by Image J (NIH) software.
Statistical analyses
Data were analyzed by SPSS19.0 statistical software (SPSS, Inc.). Data are expressed as mean ± standard deviation. t-Test was used for comparison between groups with p < 0.05 as the significant difference.
Results
The contents of cytokines were reduced by AACOCF3
The contents of cytokines in cells were analyzed by ELISA kits. As shown in Figure 1, the levels of mPGES, PGE2, and PGI2 in control group were significantly higher than those in the AACOCF3 group (p < 0.05).

AACOCF3 reduced the concentration of mPGES
AACOCF3 inhibits proliferation of lung cancer cells
The effect of inhibitor AACOCF3 on the proliferation of lung cancer cells was detected by CCK8. The result showed that the cell inhibition rate was 14.72% after adding AACOCF3, which was significantly higher than that in the control group (0.30%, p < 0.05) (Fig. 2).
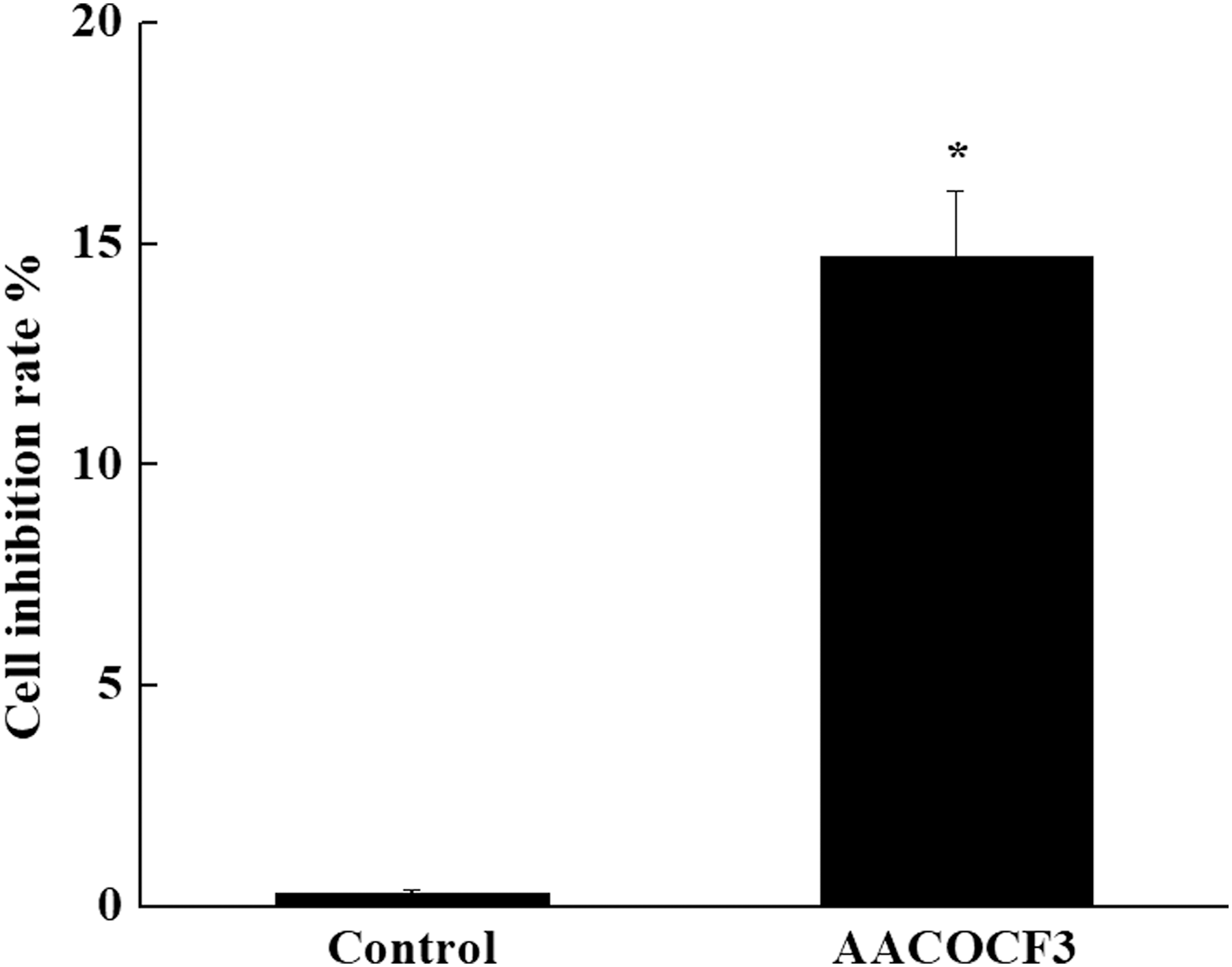
Effect of inhibitor AACOCF3 on the proliferation of lung cancer cells. Proliferation of lung cancer cells was detected by cell counting kit-8. Cell inhibition rate was 14.72% after adding AACOCF3, which was significantly higher than that in the control group. Data are expressed as the mean ± SD (n = 5). Compared with the control group, *p < 0.05.
AACOCF3 inhibits migration and invasion of lung cancer cells
Scratch wound healing test indicated that AACOCF3 significantly inhibited cancer cell migration (53.47%, p < 0.05) compared with the control group (Fig. 3A). Transwell assay also confirmed that AACOCF3 significantly inhibited metastasis and invasiveness of lung cancer cell (Fig. 3B). The relative number of cells in the control group (85.7 ± 5.0) was significantly higher than those in the AACOCF3 group (33.9 ± 2.7) (p < 0.05).

Effect of inhibitor AACOCF3 on migration and invasion of lung cancer A549 cells. Migration of cells at 0 h and 24 h was tested by scratch wound healing test
AACOCF3 inhibits the expression of PLA2G4A and COX-2 mRNA in lung cancer cells
We examined the effect of AACOCF3 on the expression of PLA2G4A and COX-2 mRNA in lung cancer cells. As shown in Figure 4, the related expression of PLA2G4A and COX-2 mRNA in the control group was significantly higher than that in the AACOCF3 group (p < 0.05).
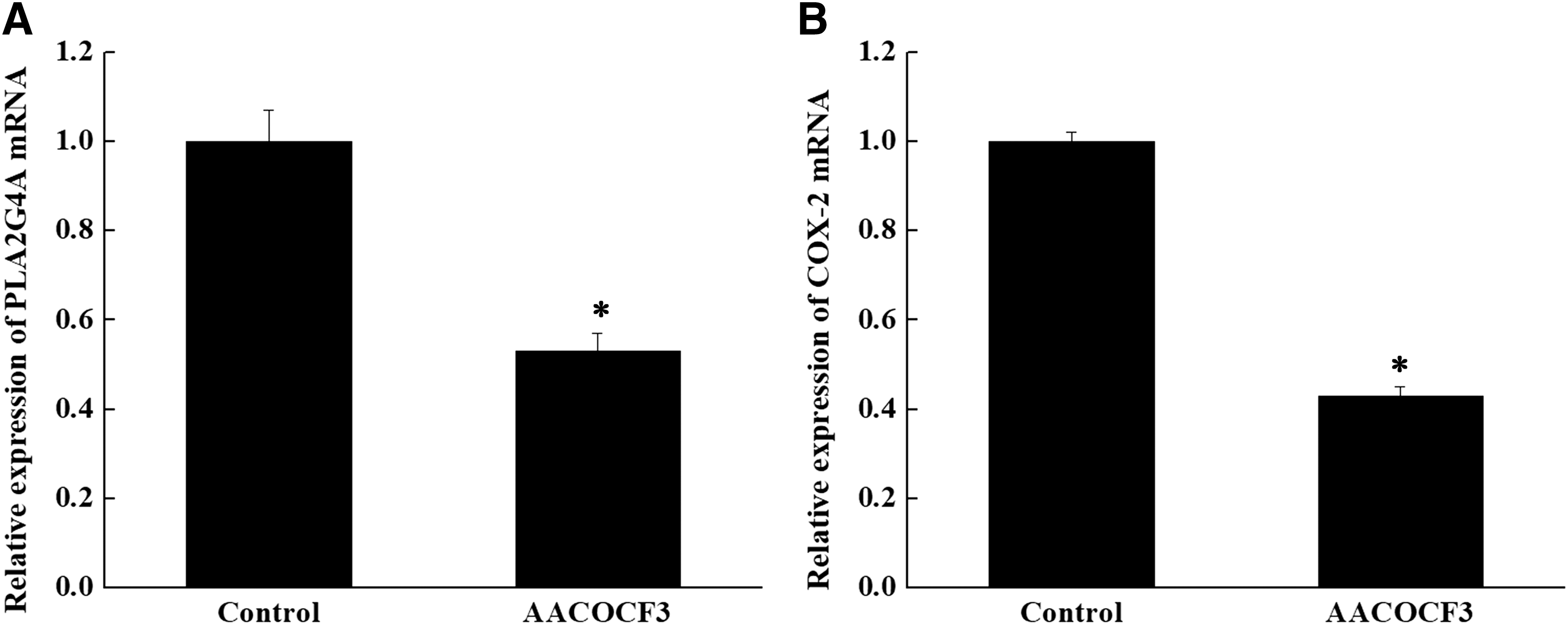
AACOCF3 inhibits the expression of PLA2G4A
AACOCF3 inhibits the expression of mPGES, COX-1, and COX-2 protein in lung cancer cells
To examine the mechanism of AACOCF3 in the progression of lung cancer, the levels of mPGES, COX-1, and COX-2 proteins were detected by Western blotting. Inhibition of cPLA2 in A549 cells significantly reduced the levels of mPGES, COX-1, and COX-2 protein compared with those in the control group (p < 0.05, Fig. 5).

AACOCF3 inhibits the expression of mPGES, COX-1, and COX-2 proteins.
Discussion
PLA2 could hydrolyze the structure of the Sn-2 lipid bond and play a key role in the metabolism of phospholipids (Maddirevula et al., 2016). PLA2 is the initiator of inflammation and is involved in many biological processes (Schaloske and Dennis, 2006). PLA2 decomposes phospholipids to produce arachidonic acid (AA) and a variety of inflammatory mediators, such as COX, PGE2 (Nakanishi and Rosenberg, 2006; Caiazza et al., 2011). In recent years, overexpression of COX-2 and PGE2 was found in many tumors (Patel et al., 2008). Membrane-associated PGE2 synthase (mPGES) is the end enzyme of PGE2 generation, which is mainly coupled with COX-2 in many physiological and pathophysiological processes, including inflammation and tumor (Kitsiouli et al., 2009; Wang et al., 2015).
COX contains two isozymes, COX-1 and COX-2 (Chen et al., 2002). Previous studies suggested that the prostaglandin produced by COX-1 was involved in the normal physiological processes and protective functions of the body, such as maintaining the integrity of the gastrointestinal mucosa (Hatzidaki et al., 2010; Prima et al., 2017), regulating the function of the platelets and the blood flow of the kidney (Letsiou et al., 2011). COX-2 is an induced enzyme, which is induced by various damaging chemical, physical, and biological factors, and then catalyzes PGs synthesis to participate in inflammatory reaction (Millanta et al., 2016).
Recent studies have shown that the activation of PLA2 is closely related to the occurrence and development of various tumors (Messaritakis et al., 2017). The high expression of cPLA2 mRNA in Luminal cast-in breast cancer patients has a poor prognosis, low overall survival, and poor efficacy of endocrine therapy (Vainio et al., 2011; Zhuang et al., 2017). The expression of PLA2 is closely related with the occurrence and development of lung cancer. Compared with normal lung tissue, PLA2 is highly expressed in lung cancer tissues (Samel et al., 2013). In our study, the expression levels of mPGES, PGE2, and PGI2 were very high in the lung cancer group, which were consistent with the previous studies.
AACOCF3, an analogue of AA, specifically inhibits the 85 kDa cPLA2. As an inhibitor of PLA2, AACOCF3 was used to treat inflammatory diseases (Touqui and Alaoui-El-Azher, 2001). In this study, the expression levels of PLA2G4A, mPGES, COX-1, and COX-2 in the control group were significantly higher than those in the AACOCF3 group. Our research further confirmed that AACOCF3 could inhibit lung cancer cells migration and invasion, which showed that PLA2G4A played a key role in lung cancer.
In summary, PLA2G4A was involved in migration and invasion of lung cancer cells, which could become a target to lung cancer. However, the correlation between PLA2G4A and clinical pathological stages of lung cancer needs to be analyzed.
Footnotes
Acknowledgments
This work was supported by Medical Health Technology Development Plan of Shandong Province (No. 2017WSA06063) and Science and Technology Planning Project of Yantai Urban (No. 2015WS004).
Disclosure Statement
No competing financial interests exist.