
Abstract
Accumulating evidence has suggested that microRNAs play important roles in the development of hepatocellular carcinoma (HCC) and are involved in drug resistance. miR-21-5p was overexpressed in a variety of cancers and promoted the tumorigenesis; however, the function of miR-21-5p in HCC still remains unknown. In this study, our results showed that miR-21-5p was highly expressed in HCC tissues and cell lines. Notably, the level of miR-21-5p was relatively higher in cisplatin (DDP)-resistant HCC patients. Overexpression of miR-21-5p attenuated the inhibitory effect of DDP on the proliferation and apoptosis of HCC cells. Mechanistically, the luciferase report assay-identified FAS ligand (FASLG) was a direct target of miR-21-5p. Overexpression of miR-21-5p decreased both the mRNA and protein levels of FASLG in HCC cells. FASLG was downregulated in HCC tissues and was significantly negatively correlated with the expression of miR-21-5p. Restoring the expression of FASLG upregulated the chemosensitivity of HCC cells expressing miR-21-5p. In conclusion, our results demonstrated that miR-21-5p targeted FASLG and suppressed the sensitivity of HCC cells to DDP treatment.
Introduction
Hepatocellular carcinoma (HCC) is one of the most prevalent cancers worldwide (Clark et al., 2015). The high morbidity of HCC makes it the leading cause of cancer-associated mortality in developing countries (El-Serag and Kanwal, 2014; Clark et al., 2015; McGlynn et al., 2015). It has been widely accepted that chronic inflammation, infections of hepatitis B and C viruses, and alcoholism contribute greatly to the initiation and development of HCC (Kudo, 2018).
Currently, surgical resection combined with the application of chemotherapy and radiochemotherapy has improved the prognosis of HCC patients. However, the 5-year overall survival (OS) of the patients still remains poor, owing to the chemoresistance of HCC (Cabibbo et al., 2010; Yu et al., 2017). Therefore, it is quite urgent to investigate the molecular basis of HCC chemosensitivity and identify novel targets that are responsible for the resistance of HCC cells to the chemotherapy drugs.
Genetic alternation has been considered the main cause of cancer initiation. microRNAs (miRNAs) are characterized as a class of single-stranded, small noncoding RNAs with the length of approximately 22 nucleotides (Cortez and Calin, 2009). As important post-transcriptional regulators of gene expression, miRNAs bind directly to the 3′-untranslated region (3′-UTR) of the targeted mRNAs, which leads to the degradation of mRNA or inhibition of translation (Fabian et al., 2010; Mohr and Mott, 2015). The function of miRNAs is tightly involved in modulating the proliferation, differentiation, and apoptosis of cells (Ambros, 2004). Accumulated evidence suggested that miRNAs act as tumor suppressors or oncogenes in the establishment and progression of human cancers (Kwak et al., 2010; Farazi et al., 2011; Qu et al., 2011). Notably, the involvement of miRNAs in the chemotherapy resistance of HCC has attracted wide attention. For example, miR-9 was recently reported to enhance the sensitivity of HCC to cetuximib by downregulating the expression of eukaryotic translation initiation factor 5A-2 (Xue et al., 2018). In addition, upregulation of miR-137 induced the degradation of ANT2 and reversed the sorafenib resistance of HCC cells (Lu et al., 2017). miR-21-5p was identified as an oncogenic miRNA that was highly expressed in a variety of human cancers (Park et al., 2016; Kao et al., 2017; Cheng et al., 2018; Ghorbanmehr et al., 2018; Li et al., 2018; Xia et al., 2018; Zhu et al., 2018). miR-21-5p promoted the metastasis of cervical cancer cells by targeting the tumor suppressor von Hippel-Lindau (Cai et al., 2018). Recent study demonstrated that inhibition of miR-21-5p enhanced the radiation sensitivity of non-small cell lung cancer by modulating the expression of human MutS homolog 2 (Song et al., 2017). These evidence suggested miR-21-5p as a key player in the progression of cancers. However, the potential involvement of miR-21-5p in the development of HCC has not been elucidated.
It has been well documented that aberrant regulation of cell apoptosis results in the initiation and progression of cancers. The FAS/FAS ligand (FASLG) signaling is a key part of the apoptotic pathway (Oh et al., 2004). FASLG was identified as a type II transmembrane protein belonging to the tumor necrosis factor superfamily (Waring and Mullbacher, 1999). Mechanistically, the interaction of membrane-bound FASLG to FAS-expressing cells leads to a proapoptotic signaling cascade that consequently induces cell apoptosis (Waring and Mullbacher, 1999). Targeting the FAS/FASLG signaling by miRNAs has been a novel strategy to modulate the growth of cancer cells.
In this study, our results demonstrated that miR-21-5p was overexpressed in HCC tissues. Notably, the level of miR-21-5p was significantly higher in HCC patients with resistance to the chemotherapy treatment. The molecular experiment uncovered that miR-21-5p targeted FASLG and downregulated the expression of FASLG in HCC cells.
Materials and Methods
Clinical tissues and cell lines
Fifty paired HCC tissues and corresponding normal tissues were obtained from HCC patients who underwent surgical resection at Linyi People's Hospital between April 2012 and June 2014. The clinical data of the patients are summarized as Supplementary Table S1. These patients never received chemotherapy or radiotherapy before the surgery. In addition, other 50 HCC samples were provided by HCC patients who underwent standard cisplatin (DDP)-based chemotherapy following the surgical resection at Linyi People's Hospital. These 50 samples were divided into DDP-sensitive or DDP-resistant groups according to the computed tomography (CT) scanning and response evaluation criteria in solid tumors (RECIST) criteria after two cycles of DDP chemotherapy. The collection and usage of these tissue samples were approved by the Ethics Committee of Linyi People's Hospital in accordance with the Declaration of Helsinki protocols (Approval No. LPH2016052213). Written informed consents were obtained from all participants.
Human HCC cell lines were purchased from the Cell Bank of the Chinese Academy of Sciences (Shanghai, China). Cells were maintained in the Dulbecco's modified Eagle's medium (DMEM; GE Health care Life Sciences, Beijing, China) supplemented with 10% fetal bovine serum and 1% penicillin/streptomycin at 37°C with 5% CO2.
Reverse transcription-quantitative polymerase chain reaction
Total RNA was extracted from tissues or cells using TRIzol reagent (Tiangen Biotech Co., Ltd., Beijing, China) according to the manufacturer's instructions. cDNA was generated with 1 μg of RNA by reverse transcription using the TaqMan MicroRNA Reverse Transcription Kit (Applied Biosystems, Foster City, CA). The expression level of miR-21-5p was detected using the SYBR Green PCR Master Mix (Applied Biosystems) on the ABI7500 Real-time PCR system (Applied Biosystem). The level of U6 RNA was detected as the internal control. The polymerase chain reaction (PCR) cycling conditions were as follows: 95°C for 5 min and 40 cycles at 95°C for 10 s and 60°C for 1 min. The relative expression ratio was calculated with the 2−ΔΔCq method.
Western blot
HepG2 and Huh7 cells were transfected with miR-21-5p mimics or miRNA negative control (miR-NC) with Lipofectamine 2000 (Invitrogen, Thermo Fisher Scientific, Inc.) according to the manufacturer's instructions. For the western blot, cells were harvested and lysed using the RIPA buffer (Beyotime Institute of Biotechnology, Shanghai, China) containing protease inhibitor. After centrifugation at 12,000 rpm for 15 min at 4°C, the protein concentration was determined with the BCA Protein Assay kit (Beyotime Institute of Biotechnology). Equal amount of protein was separated by 12% sodium dodecyl sulfate–polyacrylamide gel electrophoresis and then transferred onto the polyvinylidene fluoride membrane (Bio-Rad, CA). After blocking with 5% non-fat milk for 1 h at room temperature, the membrane was incubated with the primary antibody against FASLG (#68405; Cell Signaling Technology, MA), GAPDH (#5174; Cell Signaling Technology), cleaved caspase-3 (#9661; Cell Signaling Technology), and caspase-8 (active/cleaved, NB100-56116SS; Novus Biologicals) overnight at 4°C. Subsequently, the membrane was washed twice with phosphate-buffered saline (PBS) and then incubated with horseradish peroxidase-conjugated secondary antibody for 1 h at room temperature. The protein bands were visualized with the enhanced chemiluminescence (ECL) kit (Millipore, MA) according to the manufacturer's instructions. GAPDH was detected as the loading control for normalization.
Chemotherapy sensitivity assay
The chemotherapy sensitivity of HCC cells was evaluated with the Cell Counting Kit-8 (CCK-8; Dojindao, Japan) assay. Cells transfected with the corresponding vector were cultured in the 96-well plate with the density of 1000 cells per well. After incubating with different concentrations of DDP for the indicated time, 10 μL of CCK-8 solution were added into the medium and incubated for another 3 h at 37°C. The absorbance at the wavelength of 450 nm was measured with the microplate reader (Bio-Rad).
Cell apoptosis assay
Both HepG2 and Huh7 cells were transfected with miR-21-5p mimics or miR-NC with Lipofectamine 2000. After treatment with DDP for 24 h, cells were harvested and washed twice with precold PBS. The cell apoptosis percentage was determined by dual staining with the FITC Annexin V Apoptosis Detection Kit (BD Biosciences, CA) according to the manufacturer's instructions. The rate of apoptosis was detected by flow cytometry (FACSCalibur; BD Biosciences).
In vivo xenograft mouse model
Female BALB/c nude mice (5 weeks old; weighting 15–18 g) were purchased from Chinese Academy of Sciences (Shanghai, China) and housed in special pathogen-free condition with free access to food and drink under a 12-h dark/12-h light cycle. Two hundred microliters of cell suspension (∼1 × 105 cells) were subcutaneously injected into the left inguinal of the mice. When the tumor volume reached 60 mm3, the mice were divided into four groups (n = 8 for each group): control miRNA group; miR-21-5p group; control miRNA+DDP group; and miR-21-5p+DDP group. Accordingly, mice were intratumorally injected with the indicated miRNA at the dose of 1 nmol per day or combined with DDP (3.0 mg/kg body weight) thrice per week. After 30 days, mice were sacrificed by cervical dislocation. The tumor volume and weight were measured. The animal experiments were approved by the Ethics Committee of Linyi People's Hospital.
Dual-luciferase reporter assay
The wild-type or mutant FASLG 3′-UTR luciferase reporter plasmids were co-transfected with miR-21-5p mimics or miR-NC into HCC cells using the Lipofectamine 2000 according to the manufacturer's instructions. After transfection for 48 h, cells were collected and the luciferase activity was measured with the Dual-Luciferase reporter assay (Promega Corporation, Madison, WI). The Renilla luciferase activity was detected as the normalization. The cell number was counted for the normalization.
Statistical analysis
Data were presented as mean ± standard deviation. Statistical analysis was performed using the GraphPad Prism 5.0 (GraphPad Software, CA). Significant difference was analyzed with the Student's t-test or one-way analysis of variance (ANOVA) followed by least-significant difference post-doc test. p < 0.05 was considered statistical significant.
Results
miR-21-5p was significantly overexpressed in HCC tissues
To explore the involvement of miR-21-5p in the progression of HCC, the expression of miR-21-5p in paired HCC tissues and adjacent normal tissues was detected by the reverse transcription-quantitative PCR (RT-qPCR). As shown in Figure 1A, the expression of miR-21-5p was significantly higher in HCC tissues in comparison with that of normal tissues. An increased expression of miR-21-5p was also found in HCC patients with tumor metastasis (Fig. 1B). In addition, the level of miR-21-5p in HCC cell lines, including Huh7, Bel07404, SMMC-7721, and HepG2, was significantly enhanced compared with that in normal LO2 cells (Fig. 1C). These data suggested the overexpression of miR-21-5p in HCC. The relationship between the expression of miR-21-5p and the clinical parameters of HCC patients was analyzed. The data showed that highly expressed miR-21-5p significantly correlated with the tumor size, TNM stage, and lymph node metastasis of HCC patients (Table 1). To further characterize the potential clinical significance of miR-21-5p in HCC, the correlation between the expression of miR-21-5p with the OS and disease-free survival (DFS) was analyzed. As presented in Figure 1D and E, overexpressed miR-21-5p was significantly correlated with the reduced OS and DFS of the HCC patients, respectively. These results demonstrated the potential application of miR-21-5p in the prognosis of HCC. To detect the association between the expression of miR-21-5p and the chemotherapy resistance of these HCC patients, the level of miR-21-5p was investigated with the tissues from HCC patients with or without DDP resistance. As presented in Figure 1F, the expression of miR-21-5p was significantly increased in HCC patients with DDP resistance.

miR-21-5p was overexpressed in HCC tissues.
Correlation Between miR-21-5p Expression and Clinical Factors of Hepatocellular Carcinoma Patients
miR-21-5p decreased the sensitivity of HCC cells to DDP
As miR-21-5p was highly expressed in DDP-resistant HCC tissues, to detect the effect of miR-21-5p on the chemoresistance of HCC cells, HepG2 and Huh7 cells were transfected with miR-21-5p mimics and the expression of miR-21-5p was validated by RT-qPCR assay (Fig. 2A). The CCK-8 assay showed that the overexpression of miR-21-5p along with DDP treatment resulted in significantly enhanced viability compared with the control miRNA plus DDP-treated group (Fig. 2B, C). A reduced rate of apoptosis of both HepG2 and Huh7 cells was observed following treatment with miR-21-5p mimics, with DDP in comparison with that of the cells explored by control miRNA and DDP (Fig. 2D). Furthermore, the colony formation assay showed that overexpression of miR-21-5p attenuated the inhibitory effect of DDP on the growth of HCC cells (Fig. 2E). To further validate the influence of miR-21-5p on the DDP sensitivity of HCC cells, the nude mice xenograft model was constructed. Compared with the control miRNA group, overexpression of miR-21-5p significantly promoted the tumor growth with bigger tumor weight and volume (Fig. 2F, G). Moreover, compared with tumors treated with DDP and control miRNA, combination of miR-21-5p and DDP resulted in increased tumor growth, suggesting that overexpression of miR-21-5p attenuated DDP-induced cytotoxicity in HCC cells in vivo (Fig. 2F, G). These results indicated that highly expressed miR-21-5p reduced the sensitivity of HCC cells to DDP.
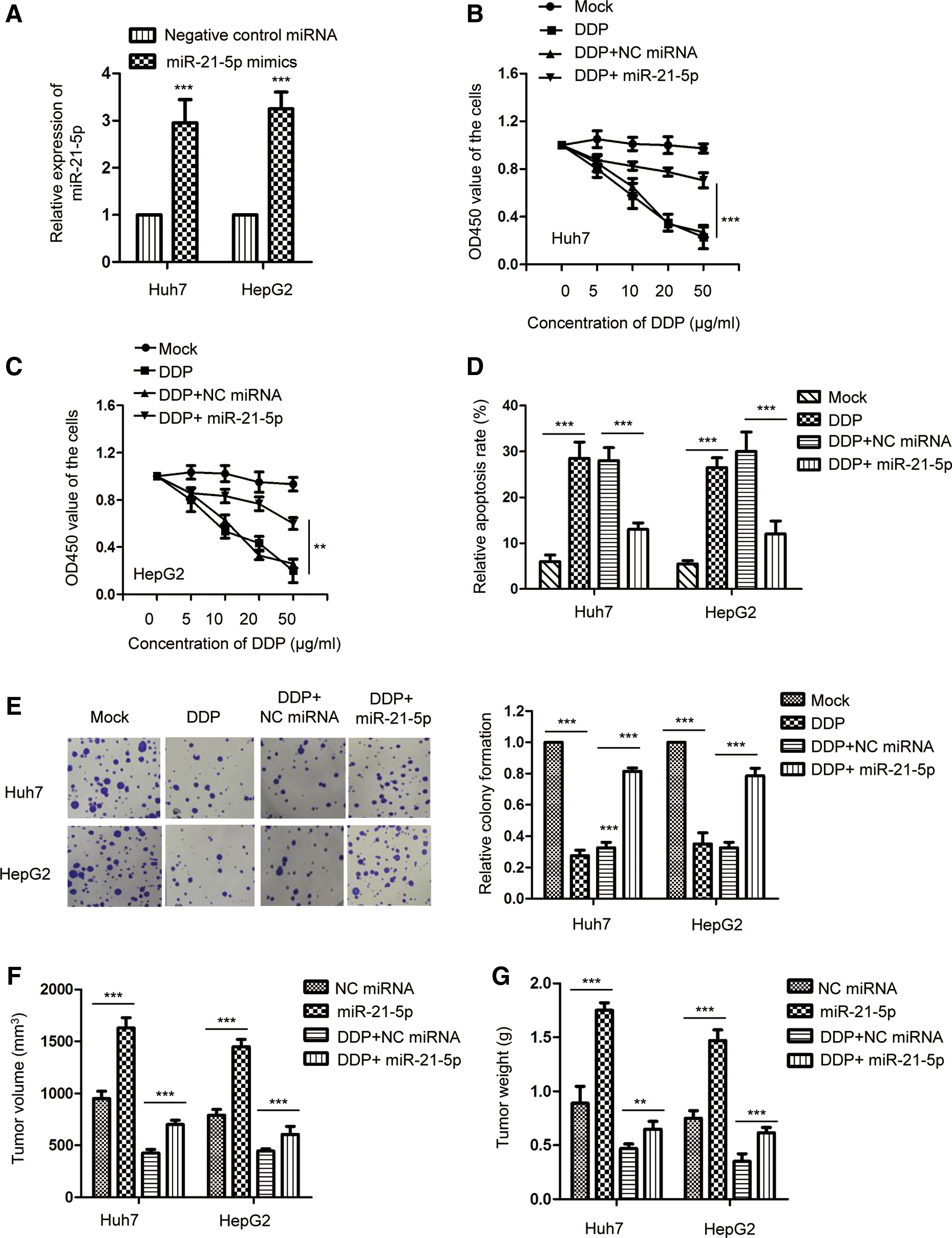
Overexpression of miR-21-5p decreased the chemoresistance of HCC cells.
miR-21-5p targeted FASLG in HCC cells
To investigate the possible mechanism by which miR-21-5p suppressed the sensitivity of HCC cells to DDP, the potential targets of miR-21-5p were predicted using the TargetScan database. The data indicated that miR-21-5p might target the 3′-UTR of FASLG (Fig. 3A). To verify this prediction, luciferase assay was performed by transfecting the luciferase reporter vector harboring wild-type or mutant 3′-UTR of FASLG into HepG2 and Huh7 cells. As shown in Figure 3B and C, the luciferase activity of wild-type FASLG 3′-UTR construct was significantly decreased with the expression of miR-21-5p, whereas no remarkable reduction was observed in cells transfecting mutant 3′-UTR of FASLG and miR-21-5p. To detect the regulation of miR-21-5p on the expression of FASLG, HepG2 and Huh7 cells were transfected with miR-21-5p or negative control miRNA; both the mRNA and protein levels of FASLG were determined by the RT-qPCR and western blot analysis, respectively. The data showed that ectopically expressed miR-21-5p significantly decreased both the mRNA and protein abundance of FASLG in HCC cells (Fig. 3D, E). To further confirm the regulation of miR-21-5p on FASLG, FASLG-related signals, such as the cleavage of caspase-8 and caspase-3, were detected with the transfection of miR-21-5p. The results showed that overexpression of miR-21-5p decreased the cleavage of both caspase-8 and caspase-3 in HCC cells (Fig. 3F). These findings demonstrated that miR-21-5p targeted FASLG and suppressed the expression of FASLG in HCC cells.
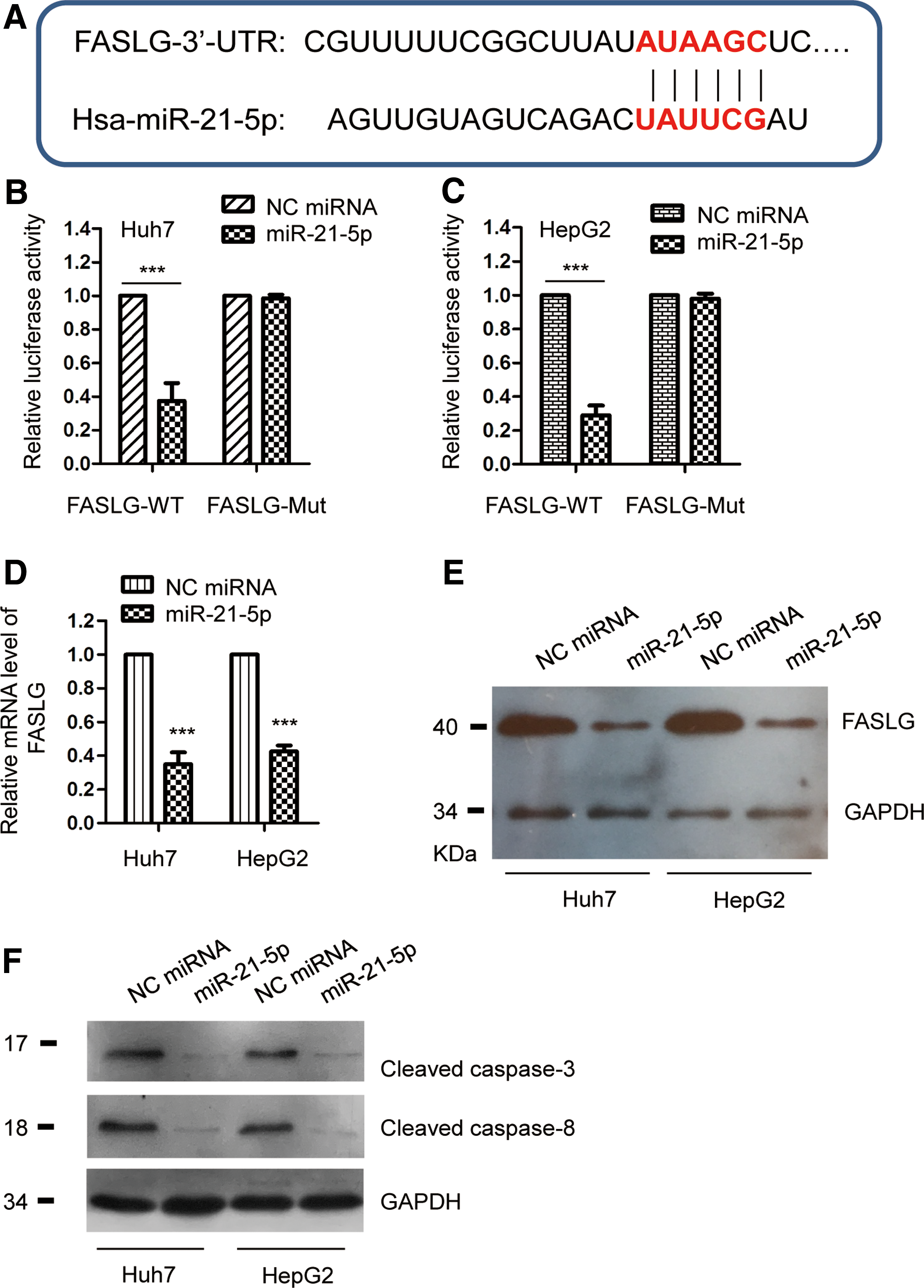
FASLG was a target of miR-21-5p in HCC.
FASLG was inversely correlated with the expression of miR-21-5p in HCC
As miR-21-5p was identified as an upstream negative regulator of FASLG, the expression of FASLG was detected in paired HCC tissues and normal control group. The result showed that FASLG was significantly downregulated in HCC tissues compared with the adjacent normal tissues (Fig. 4A). Consistently, the protein expression of FASLG was also decreased in HCC cell lines compared with the control normal cells (Fig. 4B). The Spearman correlation test revealed that the level of FASLG in HCC tissues was significantly inversely correlated with that of miR-21-5p (Fig. 4C). In addition, the expression of FASLG in DPP-resistant or DPP-sensitive HCC tissues was also evaluated by immunohistochemistry staining. As shown in Figure 4D, the expression of FASLG was decreased in HCC tissues with DDP resistance compared with that of HCC tissues that are DDP sensitive (Fig. 4D). To confirm the involvement of FASLG in miR-21-5p-mediated chemosensitivity of HCC cells, FASLG was transfected into HCC cells that were treated with DDP along with the overexpression of miR-21-5p. As shown in Figure 4E, restoration of FASLG significantly attenuated the DDP resistance of Huh7 cells caused by the high expression of miR-21-5p. Similar results were also obtained in HepG2 cells (Fig. 4F). Collectively, these results revealed that miR-21-5p downregulated FASLG and decreased the sensitivity to DDP in HCC cells.
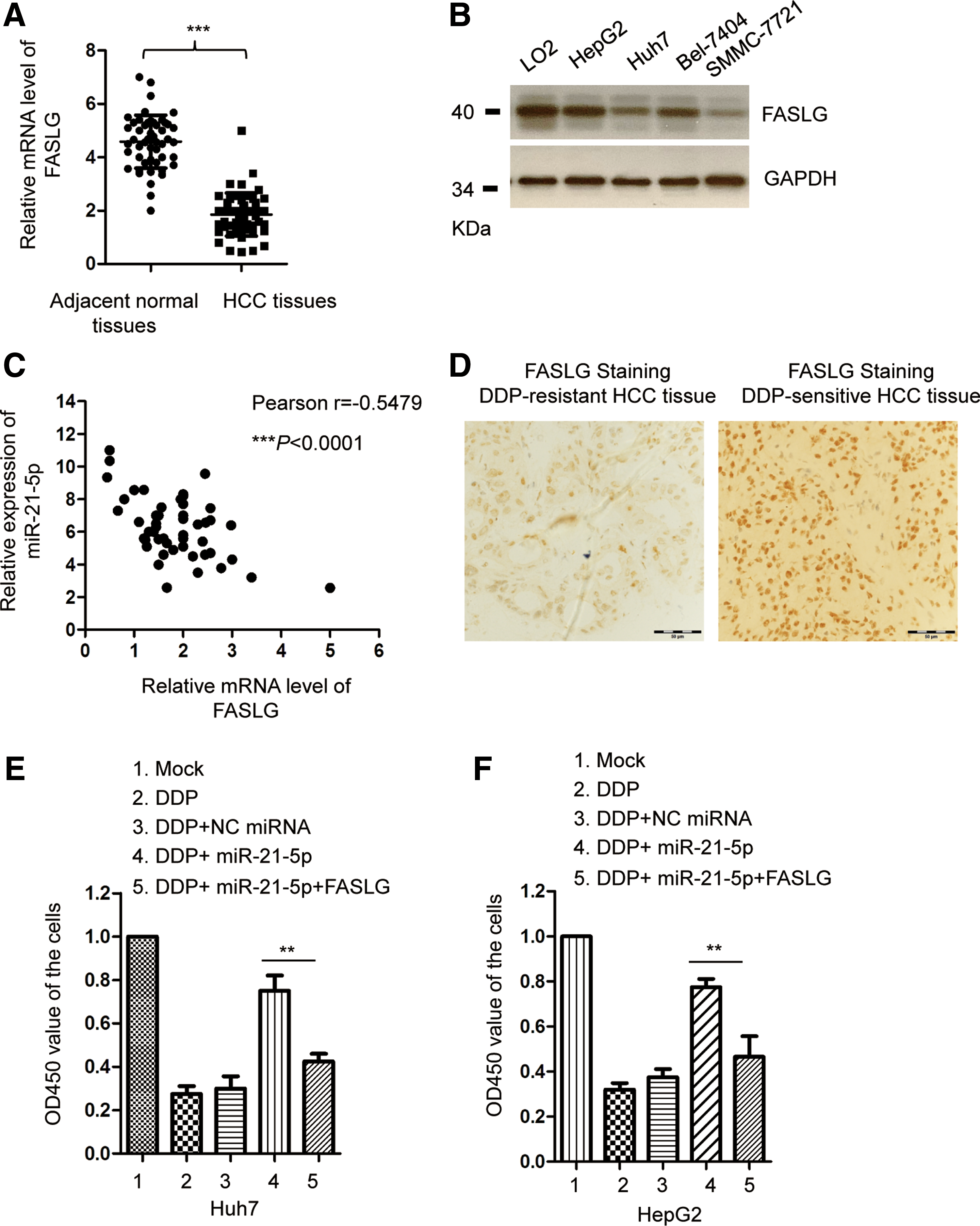
FASLG was inversely correlated with the expression of miR-21-5p in HCC.
Discussion
Remarkable progress has been made in the treatment of HCC patients; however, multiple drug resistance remains the major challenge for chemotherapy (Ding et al., 2018). Increasing evidence has suggested that dysregulation of miRNAs play critical roles in the pathogenesis of chemoresistance (Corra et al., 2018). In this study, our finding provided novel insight into the function of miR-21-5p in the DDP resistance of HCC cells. Overexpression of miR-21-5p enhanced the cell viability and decreased the apoptosis of HCC cells in the presence of DDP. Our results supported the finding that miRNAs are important regulators of chemoresistance in HCC.
miR-21-5p was initially identified as an oncogene in a variety of human cancers (Park et al., 2016; Kao et al., 2017; Ghorbanmehr et al., 2018; Li et al., 2018; Lopes-Ramos et al., 2014). Higher level of miR-21-5p was considered a urine-promising biomarker for the identification of prostate and bladder cancer (Ghorbanmehr et al., 2018). The oncogenic function of miR-21-5p was also found in Hodgkin lymphoma (Yuan et al., 2018). Reduced miR-21-5p inhibited the proliferation, migration, and invasion of esophageal cancer cells (Xia et al., 2018). Consistently, inhibition of miR-21-5p suppressed high glucose-induced proliferation and angiogenesis of retinal microvascular endothelial cells (Qiu et al., 2018). In this study, our data showed that the abundance of miR-21-5p was increased in HCC patients with DDP resistance. The functional analysis indicated that ectopically expressed miR-21-5p promoted the viability and decreased the apoptosis of HCC cells treated with DDP. These results suggested that highly expressed miR-21-5p might confer chemoresistance in HCC cells. Further study is necessary to evaluate the involvement of miR-21-5p in drug resistance by in vivo experiments.
The molecular mechanism by which miR-21-5p regulated the chemoresistance of HCC cells was investigated by identifying the targets of miR-21-5p. Our data showed that miR-21-5p bound the 3′-UTR of FASLG and negatively modulated the expression of FASLG in HCC cells. Recent study uncovered that FASLG was a target of miR-149-5p and mediated the function of miR-149-5p in regulating the apoptosis of acute myeloid leukemia cells (Tian and Yan, 2016). Notably, decreased level of FASLG has been demonstrated in cancers, which decreased the apoptosis of cancer cells (Liu et al., 2009). In addition to HCC, FASLG was also identified as the downstream target of miR-21-5p in glioblastoma cancer and gastric cancer (Yang et al., 2014; Shang et al., 2015). In this study, restoring the expression of FASLG deduced the chemoresistance of HCC cells to DDP. It would be interesting to investigate whether miR-21-5p also modulates the drug resistance in other types of cancers.
In conclusion, our results showed that overexpressed miR-21-5p decreased the chemosensitivity and promoted the growth of HCC cells by targeting the expression of FASLG. These findings provided novel insights into the potential application of miR-21-5p/FASLG signaling to overcome DDP resistance in HCC treatment.
Conclusions
Our results demonstrated that miR-21-5p was significantly upregulated in HCC. Overexpression of miR-21-5p decreased the chemosensitivity of HCC cells by targeting FASLG. These findings suggested that the miR-21-5p/FASLG axis is a potential target to overcome the chemoresistance of HCC.
Footnotes
Disclosure Statement
No competing financial interests exist.
Supplementary Material
Supplementary Table S1
References
Supplementary Material
Please find the following supplemental material available below.
For Open Access articles published under a Creative Commons License, all supplemental material carries the same license as the article it is associated with.
For non-Open Access articles published, all supplemental material carries a non-exclusive license, and permission requests for re-use of supplemental material or any part of supplemental material shall be sent directly to the copyright owner as specified in the copyright notice associated with the article.