
Abstract
Polycystic ovary syndrome (PCOS) is the most typical metabolic syndrome in women of reproductive age, with a high prevalence and an increased risk of long-term complications. PCOS mainly manifests as hyperandrogenism (HA), ovulatory dysfunction, and polycystic ovaries, in addition to being relevant to infertility, insulin resistance (IR), obesity, lipid abnormalities, and chronic low-grade inflammation. The etiology of this syndrome remains largely unknown. microRNAs (miRNAs), small, noncoding RNAs (nearly 22 nucleotides long), regulate gene expression at the posttranscriptional level. Abnormal miRNA levels are closely associated with the occurrence of diseases, such as diabetes, cancers, and atherosclerosis, and miRNAs can be used as predictors and diagnostic biomarkers for cancer. Interestingly, the roles of miRNAs in PCOS pathology have attracted considerable attention in recent years. Research has established that alterations in miRNA expression in women with PCOS compared with healthy women may act as noninvasive biomarkers and new therapeutic targets in PCOS. This article aims to summarize the latest research on the relationship between miRNAs and the clinical manifestations of PCOS while also providing a few mechanisms based on previous studies. Understanding the relationship between miRNAs and PCOS will provide guidance for researchers to further explore the complexity and heterogeneity of PCOS.
Introduction
Polycystic ovary syndrome (PCOS), a complicated and heterogeneous endocrine condition, affects nearly 5–10% of all women of reproductive age (Yildiz et al., 2012). The definition of PCOS is based on three different criteria, namely, the National Institutes of Health (NIH) criteria, Rotterdam criteria, and Androgen Excess Society (AES) criteria, all of which, to some extent, include polycystic ovarian morphology, clinical and/or biochemical hyperandrogenism (HA), and chronic anovulation (Fux et al., 2013; Christakou and Diamanti-Kandarakis, 2014; Jiang and Ma, 2018) (Fig. 1). With the establishment of the Rotterdam criteria, the prevalence of PCOS and variability of the phenotype in the patient population noticeably increased. Therefore, the Rotterdam criteria are generally recommended for the diagnosis of PCOS: any woman who presents with at least two of the three described characteristics can be diagnosed with PCOS. Notably, the diagnosis must exclude typical signs and symptoms that overlap with PCOS, such as congenital adrenal cortical hyperplasia, Cushing syndrome, hyperprolactinemia, thyroid dysfunction, and androgen-secreting tumors (Rotterdam ESHRE/ASRM-Sponsored PCOS Consensus Workshop Group, 2004, Azziz et al., 2009). Although the biochemical manifestations and clinical characteristics of PCOS are typical, PCOS is still a complicated pathophysiological condition with no gold standard for diagnosis.
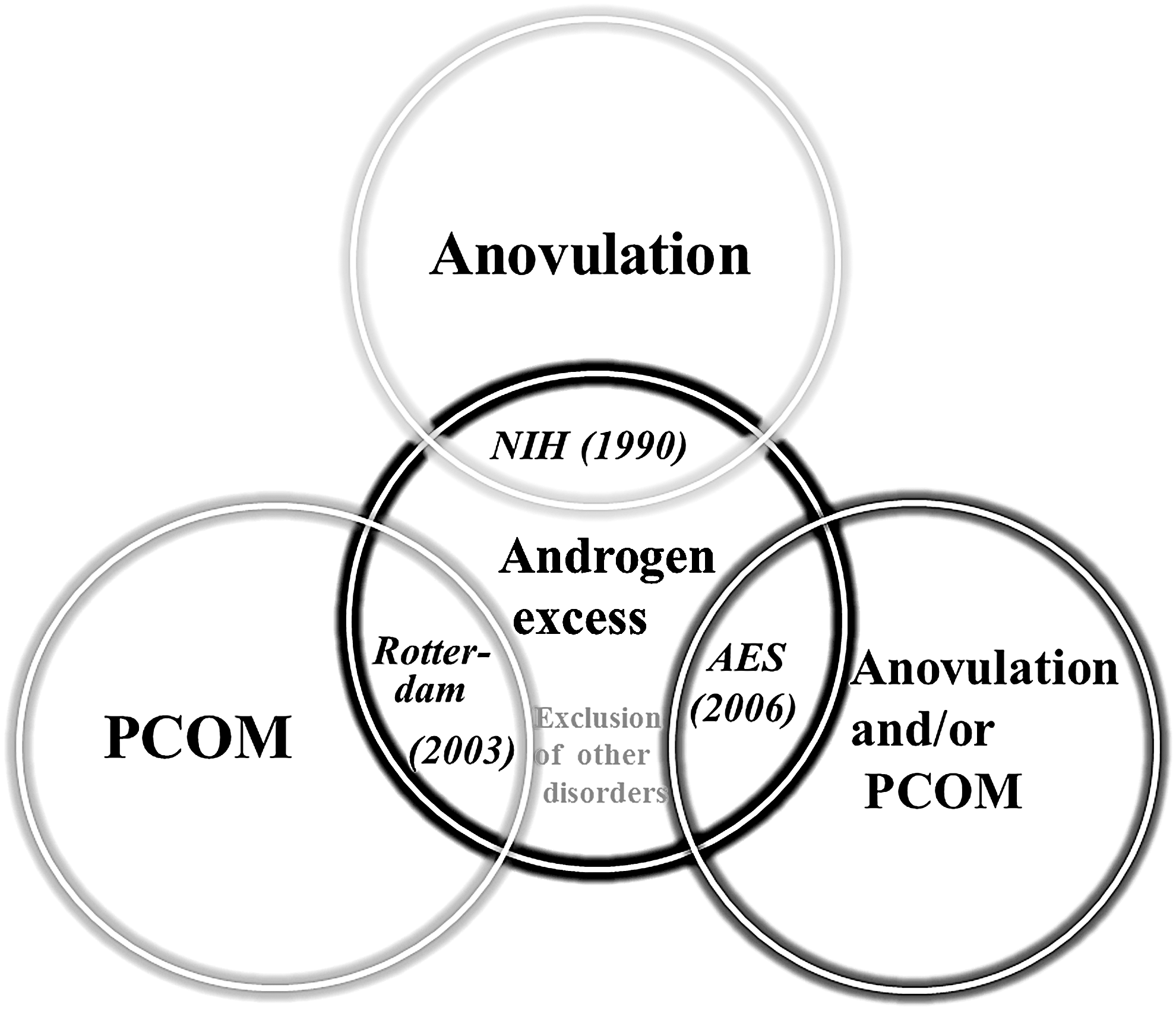
Diagnostic criteria and heterogeneous of PCOS. PCOS is a heterogeneous disease of women that is represented by clinical and/or biochemical signs of androgen excess, ovulatory dysfunction, and polycystic ovarian morphology. The definition of PCOS is based on three different criteria, namely NIH criteria, Rotterdam criteria, and AES criteria. Hyperandrogenism as defined either based on biochemical or clinical characteristics. All of the diagnostic criteria must exclude other disorders presenting with androgen excess.
PCOS is associated not only with infertility due to follicle growth arrest at the small antral stage, decreased granulosa cell (GC) proliferation, and HA but also with an increased prevalence of endometrial hyperplasia, ovarian cancer (Azziz et al., 2009), and metabolic diseases, such as cardiovascular diseases, obesity, and diabetes (Christakou and Diamanti-Kandarakis, 2014; Romakina et al., 2018). In addition, PCOS is also related to the occurrence of pregnancy complications, such as gestational diabetes mellitus (GDM), pregnancy-induced hypertension, placental abruption, premature rupture of membranes, and preterm birth. However, there is a lack of consistent and effective methods to assess the risk of these complications. Moreover, treatment by means of various drugs can alleviate the symptoms and signs of PCOS, but these treatments do not significantly improve the prognosis of PCOS.
microRNAs (miRNAs), which were first discovered in 1993, are widely present in the body and can be detected in serum, plasma, urine, semen, and follicular fluid (FF) (Sorensen et al., 2014). miRNAs negatively regulate the expression of their target genes in a posttranscriptional manner by cleaving the target mRNAs or inhibiting translation (Yates et al., 2013). Although ∼2800 miRNAs have been identified in Homo sapiens, the functions of most of them are not entirely clear (Kozomara and Griffiths-Jones, 2014). Accumulative studies have shown that miRNAs regulate ∼60% of human protein-coding genes and are involved in various physiological processes, including development, metabolism, apoptosis, differentiation, and cell cycle (Rossi, 2009; Xue et al., 2018). In addition, the existence of feedback regulatory mechanisms among miRNAs, their regulatory targets, and their products allow for the suppression or amplification of a specific signal. Hence, alterations in the expression levels of a small portion of miRNAs may contribute to many diseases, including cervical cancer, endometriosis, poor ovarian response, and cardiovascular diseases (Cortez et al., 2011; Bu et al., 2018; Mari-Alexandre et al., 2018; Romakina et al., 2018) (Fig. 2). Specifically, new evidence suggests that miRNAs are differentially expressed between women with PCOS and women without the disease (Long et al., 2014; Ding et al., 2015). Moreover, miRNAs are stable in serum and easy to detect due to their resistance of nuclease activity. Therefore, miRNAs may be candidate diagnostic markers for patients with PCOS (Ilie and Georgescu, 2015). However, our understanding of the relationship between miRNAs and PCOS is at a preliminary stage, and the possible roles of miRNAs in the pathogenesis of PCOS have not yet been clarified.

Biogenesis and function of miRNA. The miRNA gene is transcribed by RNA polymerase II to form the primary miRNA (pri-miRNA), then cleaved to pre-miRNA by the microprocessor complex (Drosha–DGCR8). Subsequently, to form mature miRNA in the cytoplasm, miRNA binding to its mRNA target regulates development, metabolism, apoptosis, and differentiation process by direct degradation or translational repression. Alteration of the expression of miRNAs may contribute to many diseases occurrence and development, including PCOS, cervical cancer, and cardiovascular diseases.
In light of these considerations, the aim of this review is to summarize the relationship among miRNAs and insulin resistance (IR), HA, abnormal lipid metabolism, and chronic anovulation in PCOS on the basis of previous studies. A more comprehensive understanding of the underlying connections behind this complicated and heterogenic syndrome will improve the diagnoses and outcomes of patients with PCOS.
miRNAs May Be New Biomarkers for PCOS
Differential expression of miRNAs in plasma
In past research, increased plasma levels of luteinizing hormone (LH) and androgen, low or normal levels of follicle-stimulating hormone (FSH), and abnormalities in estrogen secretion were suggested as diagnostic methods for women with PCOS (Franks, 1995). However, evidence suggests the existence of a minority of patients with PCOS without these hormonal level changes.
Recently, mounting research has shown that miRNAs are closely related to the occurrence of PCOS and may be used as new diagnostic markers for PCOS. Murri et al. (2013) found that the levels of miR-21, miR-155, miR-103, and miR-27b are significantly increased in PCOS patients compared with healthy women and men. Furthermore, the abnormal expression of miRNAs is associated with metabolic and inflammatory processes by target gene analysis and bioinformatics analysis. Long et al. (2014) found that serum miR-16, miR-222, miR-106b, miR-19a, miR-146a, miR-30c, miR-186, and miR-24 levels were increased in women with PCOS, while miR-320 levels are decreased in women with PCOS. Further Q-PCR analysis of the expression of nine miRNAs in a study population that included 68 PCOS patients and 68 normal controls showed that miR-30c, miR-146a, and miR-222 were significantly upregulated in women with PCOS. In addition, miR-222 combined with miR-146a and miR-30c can be used to distinguish women with PCOS from controls. Interestingly, evidence demonstrates that increased miR-222 expression is associated with type 2 diabetes and GDM (Ortega et al., 2014; Shi et al., 2014).
Ding et al. (2015) found that five miRNAs (let-7i-3pm, miR-5706, miR-4463, miR-3665, and miR-638) were significantly increased and that 4 miRNAs (miR-124-3p, miR-128, miR-29a-3p, and let-7c) were decreased in PCOS patients. More importantly, Gene Ontology (GO) and Kyoto Encyclopedia of Genes and Genomes (KEGG) analyses showed that these abnormally expressed miRNAs in PCOS participate in apoptosis, angiogenesis, the immune system, ATP binding, MAPK signaling, p53 signaling pathways, and the response to reactive oxygen species. Therefore, all these findings suggest that altered miRNA levels might contribute to the pathogenesis of PCOS.
Differential expression of miRNA in FF
Oocytes are considered good-quality oocytes when they undergo meiosis and achieve fertilization, followed by embryogenesis and development. FF provides an appropriate intrafollicular environment for oocytes maturation and development. Various pathological conditions that disrupt the intrafollicular environment may affect the quality of oocytes, the development of the early embryo, and the outcome of the pregnancy (Da et al., 2018). FF comprises the secretory and metabolic activity product of the oocyte, GCs, and theca cells (TCs) and the transfer of blood plasma components. FF is easily obtained from in vitro fertilization and oocyte harvesting procedures, which may make investigations of oocyte quality, maturation, and fertility outcomes convenient. In addition, analysis of FF ingredients may also reflect metabolic and functional changes in the oocyte, GCs, and TCs. Furthermore, alterations in blood circulation biochemical components may suggest changes in the composition of FF (Da et al., 2018).
The identification of miRNAs in body fluids has motivated researchers to explore their specific functions in various physiological processes. Research has confirmed that FF provides an abundant source of miRNAs and that these miRNAs can be isolated from human FF. Therefore, FF is valuable in determining the relationship between abnormal miRNA expression and PCOS development (Sang et al., 2013; Roth et al., 2014). For example, Sang et al. (2013) detected more than 100 differentially expressed miRNAs in the FF of PCOS patients. Xue et al. (2018) found that 177 miRNAs increased and 86 miRNAs decreased in the FF of PCOS patients. Roth et al. (2014) found 29 differentially expressed miRNAs between PCOS patients and controls. Moreover, of the 29 miRNAs, hsa-miR-135, hsa-miR-32, hsa-miR-18b, hsa-miR-9, and hsa-miR-34c were negatively correlated with the mRNA expression of synaptotagmin 1, insulin receptor substrate 2, and interleukin 8 in the FF of the PCOS samples.
In conclusion, these results indicate that there are differentially expressed miRNAs in the serum and FF of PCOS patients. Therefore, it is reasonable to suggest that an assessment of distinct miRNAs that are present in serum and FF may provide new biomarker-based approach for a PCOS diagnosis. Moreover, a characterization of differential miRNA expression in FF can improve the sensitivity and specificity of the PCOS diagnosis or point toward different etiologies of the syndrome. However, the exact mechanism of the changes in plasma and FF miRNAs and whether these miRNAs are biologically functional or simply biomarkers in PCOS patients is still a large concern, and more studies are needed to confirm these observations in a larger cohort. Furthermore, FF contains various hormones and proteins as well as components that are altered in women in different age groups. Thus, the miRNA profiles of FF collected from younger and older women are different (Diez-Fraile et al., 2014). Further studies of miRNAs in FF might provide a new method for improving the intrafollicular microenvironment and/or for recovering anovulation in PCOS.
miRNA Expression and Abnormal Follicular Development in PCOS
PCOS is a systemic disease that is involved in endocrine, metabolic, and other processes. Genetic and environmental features are key factors in the pathogenesis of PCOS, whereas the etiology of PCOS is still ambiguous. Many different hypotheses have been proposed to explain the mechanisms responsible for ovulatory dysfunction, follicle growth arrest, and excessive abnormal follicle formation in PCOS samples, but there is no breakthrough finding that which can confirm the mechanistic basis of the disease.
The normal reproductive function of women is a complex process that is affected by multiple factors. The accurate expression of genes, the moderate release of sex hormones, and the adequate production of cytokines are necessary for the development of follicles. Disorders in these factors can lead to reproductive dysfunction in women of childbearing age. Data suggest that ∼80% of patients with PCOS suffer from anovulatory infertility (Balen et al., 2016).
Recently, mounting evidence has suggested that miRNAs are involved in primordial follicle activation, follicle development, oocyte maturation, and ovulation (Hasuwa et al., 2013; Yang et al., 2013; Xue et al., 2018). Abnormal expression and regulation of miRNAs are often associated with follicular development in PCOS (Table 1). Hossain et al. (2013) found 72 upregulated miRNAs and 17 downregulated miRNAs in dihydrotestosterone (DHT)-treated ovaries compared with untreated rat ovaries. Most of these differentially expressed miRNAs were predominantly localized to GCs, TCs, and oocytes at early stages of follicular development. miR-222 was expressed in the follicular membrane and GCs of early follicles, and its expression disappeared in GCs when the follicle developed into the mature stage (Hossain et al., 2013). miR-21 is overexpressed in PCOS patients and was found to reduce large tumor suppressor kinase (LATS1) expression and prompt secondary follicle growth (Jiang et al., 2015). Yang et al. (2013) found that miR-145 regulates the initiation of primordial follicle development and maintains primordial follicle dormancy by targeting transforming growth factor β2 (TGF-β2).
The Relationship Between miRNA and Nonovulation, Androgen Overdose, Insulin Resistance, and Obesity in PCOS
GCs, granulosa cells; HA, hyperandrogenism; IR, insulin resistance; IGM, impaired glucose metabolism; LDL-C, low-density lipoprotein cholesterol; LDL, low-density lipoprotein.
→, promotion; ⊣, inhibition.
Moreover, miRNAs exhibited differential expression at different stages of oocyte development. Thirteen differentially expressed miRNAs were identified in human FF from MII oocytes compared with germinal vesicle oocytes, while seven differentially expressed miRNAs were identified in human FF from MII oocytes compared with MI oocytes. In addition, these differentially expressed miRNAs might regulate the timing of oocyte meiosis and maturation by targeting the gonadotropin-releasing hormone signaling pathway (Moreno et al., 2015). Therefore, investigations of the composition and expression of miRNAs in FF may reflect the state of the oocyte.
The interaction between oocytes and surrounding GCs is critical to the development of follicles. Hence, the proliferation and apoptosis rate and differentiation state of GCs control the quality of ovarian folliculogenesis (Table 1). More extensive research has been performed to investigate the roles of miRNAs in regulating GC apoptosis and proliferation balance. Sirotkin et al. (2010) found that the apoptosis of human GCs was increased by transfection with 80 different miRNAs mimicking endogenous precursor. Yang et al. (2012) demonstrated that apoptosis was increased by overexpression of pre-miR-23a in cultured human luteinized GCs, and the potential mechanism is linked to a decrease in antiapoptotic X-linked inhibitor of apoptosis protein (XIAP) expression. Song et al. (2016) demonstrated that serum miR-324-3p levels are decreased in PCOS patients. Jiang and Ma (2018) further confirmed that the miR-324-3p level was obviously decreased in the ovarian tissues of PCOS model rats, and transfection with a miR-324-3p mimic inhibited the proliferation and promoted the apoptosis of GCs by directly targeting WNT2B, a member of the WNT family that regulates drosophila embryonic development.
miR-15a and miR-182 are essential factors in ovarian GC physiology that function by regulating proliferation, apoptosis, and steroidogenesis, while the level of miR-182 is significantly decreased in the ovarian tissue of a DHT-treated rat PCOS model (Hossain et al., 2013; Sirotkin et al., 2014). Moreover, miR-15a also regulates steroidogenesis in GCs under the control of gonadotropins (Sirotkin et al., 2009, 2014). miR-143 is differentially expressed in the FF of PCOS subjects (Roth et al., 2014). Another study also indicated that upregulated expression of miR-143 suppresses the formation of primordial follicles by inhibiting GC proliferation, suggesting that miR-143 is involved in primordial follicle formation (Zhang et al., 2013a). miR-224 has been identified in FF (Sang et al., 2013; Roth et al., 2014). TGF-β is crucial to regulate ovarian follicular growth by regulating GC function and oocyte maturation. The proliferation of GCs isolated from mouse preantral follicles that were treated with TGF-β1 was increased by upregulating miR-224, while blocking the TGF-β receptor attenuated GC proliferation (Yao et al., 2010). These results highlighted that miRNAs are necessary for controlling the apoptosis and proliferation of GCs and folliculogenesis.
In addition, a decreased rate of follicular atresia is also a feature of PCOS (Webber et al., 2007). Lin et al. (2012) found that miR-92b, let-7a, and let-7i are downregulated in atretic follicles compared with normal follicles. However, miR-26b is upregulated during follicular atresia and suppressed ataxia telangiectasia mutation expression, a protein known to modulate DNA repair, resulting in GC apoptosis. miR-125b is also upregulated in both mouse GCs and KGN cells and human GCs during follicular atresia under the regulation of testosterone and DHT (Sen et al., 2014). These results suggest that the expression levels of miRNAs are closely related to the process of follicular atresia in PCOS.
Generally, abnormal expression of miRNAs might play an important role in folliculogenesis by regulating GC proliferation and apoptosis, which might lead to chronic ovulation and infertility in PCOS patients. Nevertheless, the potential mechanisms of anovulation in PCOS patients remain largely uncertain due to the complexity of follicular development. Therefore, more studies are needed to confirm whether abnormal expression of miRNAs affects human folliculogenesis by regulating GC proliferation and apoptosis via the same pathway or molecule.
miRNAs and Ovarian Androgen Production
The ovarian cycle is regulated by hormones released from the hypothalamic–pituitary–ovarian axis. Abnormal hormone production is an important characteristic of PCOS. Androgen excess is the typical manifestation of PCOS in terms of the endocrine disorder. The excessive androgen levels may be attributed to enhanced steroidogenic enzyme activity, increased androgen production and/or defects in androgen metabolism (Kempna et al., 2015). With regard to androgen excess, accumulative research suggests that androgen excess is the main cause of oligoovulation, facilitates IR, and causes metabolic dysfunction by promoting abdominal fat deposition and visceral adiposity in PCOS patients, which in turn favors ovarian and adrenal androgen production (Dumesic et al., 2016; Escobar-Morreale, 2018). Moreover, androgen excess is accompanied by various symptoms and signs, including hirsutism, acne, seborrhea, and alopecia (Escobar-Morreale, 2018).
Androgens exert their function mainly via the androgen receptor (AR). AR is generally expressed in the GCs of growing follicles. Research over recent decades has shown that several androgens, including DHT, testosterone, and androstenedione (A4), participate in female fertility by regulating follicle development, oocyte maturation, and ovulation via the AR (Walters, 2015). Studies have found that the expression of AR is increased in PCOS patients (Apparao et al., 2002). Testosterone is bound to albumin or sex hormone binding globulin (SHBG), thus, testosterone levels are increased in PCOS due to decreased SBEG. Although the etiology by which PCOS/HA occurs remains largely uncertain, the underlying mechanism and interventions for HA in PCOS are currently hot topics of research.
The oocyte in the follicle is surrounded by inner GCs and outer TCs, all of which are steroidogenically active. Importantly, several previous studies have demonstrated that miRNAs participate in the regulation of steroid hormone production (Table 1). Cell-based studies by Sang et al. (2013) have shown that transfection with miR-193b, miR-483-5p, and miR-24 mimics is associated with decreased progesterone production. In addition, transfection with miR-320, miR-132, miR-222, and miR-520c-3p mimics promotes estradiol production in KGN cells, while transfection with a miR-24 mimic decreases estradiol production. Long et al. (2014) found that miR-146a is negatively correlated with serum testosterone. Sirotkin et al. (2009) found that miR-107 increases the secretion of testosterone in GCs.
The relationship between miRNA and abnormal androgen production in PCOS has also been extensively studied. miR-21, miR-103, and miR-155 were positively associated with free testosterone levels in PCOS patients, suggesting that the disordered PCOS hormone profiles could partly be explained by abnormal levels of miRNAs (Murri et al., 2013). Sorensen et al. (2014) have summarized the presence of several miRNAs related to androgen metabolism in the FF of PCOS patients. miR-518, miR-29a, and miR-320 are positively correlated with serum testosterone, while miR-151 is negatively correlated with serum testosterone. miR-135, miR-146a, miR-9, miR-132, and miR-18b suppress testosterone secretion. miR-151, miR-29a, miR-155, and miR-132 participate in androgen metabolism in PCOS samples. Arancio et al. (2018) also proved that miR-155 and miR-29a are negatively correlated with androstenedione concentrations in women with PCOS. These results suggest a network of miRNAs that target ovarian androgen production in PCOS subjects.
Androgen excess is the main hormone metabolic disorder of PCOS. From a physiological perspective, androgen synthesis by TCs is regulated by LH; GCs convert testosterone and A4 to estrogen via FSH-mediated regulation. Genes and many key steroidogenic enzymes, such as CYP19A1, 3b-HSD, and CYP17, are involved in steroid hormone synthesis, lipid transport, and cellular signaling and are necessary for regulating ovarian androgen and estrogen metabolism. Aromatase is necessary for the synthesis of estrogens, which are produced in GCs under CYP19A1 gene regulation. Overexpression of miR-378 reduces the expression of aromatase, leading to a decrease in estradiol production in GCs isolated from porcine ovaries (Xu et al., 2011). Moreover, miR-181a also regulates the CYP19A1 gene, which downregulates estrogen biosynthesis in mouse GCs (Zhang et al., 2013b). miR-193a-5p and miR-199a-3p are positively associated with estradiol and SHBG levels and negatively associated with free and total testosterone levels in women with PCOS. Predictions of target genes indicate that these miRNAs target several steroidogenic enzymes. Thus, miRNAs should be investigated further in detail, as they appear to participate in the development of PCOS hyperandrogenemia (Murri et al., 2018).
Even so, there are no in-depth studies on how miRNAs cause androgen metabolic disorder in patients with PCOS. Therefore, understanding the mechanism, by which miRNAs contribute to the androgen production, will greatly help with improving the clinical symptoms and prognosis of PCOS.
miRNAs and IR in PCOS
IR is a common abnormality in PCOS. Approximately 50–70% of patients with PCOS suffer from IR, resulting in an increased risk of type 2 diabetes and metabolic syndrome (Diamanti-Kandarakis and Papavassiliou, 2006). Insulin, which acts as a cogonadotropin on the ovary, stimulates adrenal androgen secretion and regulates the pulsatility of LH secretion. Importantly, IR is often accompanied by compensatory hyperinsulinemia and HA. This may be attributed to insulin-mediated stimulation of androgen production that occurs via CYP17 enzyme activation and increased cAMP concentrations, which are worked by synergistic actions with LH in TCs and decreased production of SHBG, thereby increasing free testosterone (Burghen et al., 1980; Zhang et al., 2000). Accordingly, hyperinsulinism increases the incidence of PCOS (Escobar-Morreale, 2018).
In the context of IR, miRNA expression profiles have been investigated in PCOS (Table 1). Long et al. (2014) found that miR-222 is positively associated with serum insulin levels. In the study by Ling et al. (2009), 3T3-L1 cells showed IR, and the level of miR-320 was significantly increased by the administration of high levels of glucose and insulin. However, anti-miR-320 oligos ameliorated IR by increasing the expression of glucose transporter 4 (GLUT4) and promoting the insulin-mediated uptake glucose. miR-320 has been found to exist in FF from PCOS women (Sang et al., 2013), suggesting that miR-320 may act as a potential target to improve insulin sensitivity. Jiang et al. (2016) found that miR-193b, miR-194, and miR-122 were upregulated in PCOS patients with impaired glucose tolerance (IGT) compared with PCOS patients with normal glucose tolerance. Moreover, KEGG analysis of potential signaling pathway targets revealed that these miRNAs are involved in glycometabolism and ovarian follicle growth. Therefore, these studies expand our knowledge that miRNAs participate in the development of IR in patients with PCOS.
GLUT4, an insulin-stimulated glucose transporter, is positively correlated with insulin sensitivity in primary adipocytes isolated from PCOS patients (Chen et al., 2013). miR-223 and miR-93 are significantly increased in adipose tissue from IR patients, regardless of whether they had PCOS. Its mechanism is related to a decrease in GLUT4 expression and interference with insulin-stimulated glucose metabolism (Chen et al., 2013; Chuang et al., 2015). Yang et al. (2018) found that miR-33b-5p expression was increased in the ovarian tissues of insulin-resistant PCOS rats, and miR-33b-5p levels were negatively correlated with GLUT4, high-mobility group A2 (HMGA2) and sterol regulatory element-binding protein 1 (SREBF1) expression. In addition, the expression of miR-33b-5p was also increased in excess-glucose or insulin-treated adipocytes, whereas the levels of GLUT4, SREBF1, and HMGA2 expression were reduced. Moreover, the expression of GLUT4, SREBF1, and HMGA2 was increased by inhibiting miR-33b-5p. Thus, these findings revealed that miR-33b-5p is an important molecule for regulating IR development in PCOS patients, and the possible mechanism is inhibition GLUT4 of production via targeting of HMGA2 and SREBF1. In addition, Shi et al. (2015) suggest that miR-483-5p reduces IR and facilitates cumulus cell proliferation by activating PI3K/Akt.
In summary, miRNA affects glucose metabolism and IR in PCOS patients mainly by regulating the expression of GLUT4. However, it is not clear whether miRNA regulates IR in PCOS by regulating the expression of other glycometabolism-related proteins or enzymes. Therefore, it is important to explore the roles of hyperinsulinemia-related miRNAs in the pathogenesis of PCOS.
miRNA, Obesity, and Dyslipidemia in PCOS
PCOS is associated with obesity and dyslipidemia. Obesity plays a definitive role in the metabolic abnormalities of PCOS; ∼38–88% of women with PCOS are obese or overweight. Dyslipidemia is also the typical metabolic disorder of PCOS, even though most women with PCOS show no changes in their blood lipid profiles, and qualitative alterations in lipoproteins have also been described in PCOS (Legro et al., 2001; Berneis et al., 2007). Abnormal lipid metabolism in PCOS patients is closely related to obesity, IR and HA. To date, the etiology of obesity and the lipid metabolism imbalance in PCOS has not been studied in detail, and the existing data remain unclear.
miRNAs play fundamental roles in the maintenance of metabolic homeostasis. The relationship between miRNAs and abnormal lipid metabolism in PCOS patients is a hot topic in endocrine and reproductive fields (Table 1). Murri et al. (2013) found that the expression levels miR-27b, miR-103, and miR-21 are significantly reduced in obesity but tend to increase in women with PCOS. Xiong et al. (2017) found that miR-23a/b levels are decreased in PCOS subjects, while body mass index (BMI) has positive effects on the levels of serum miR-23b, indicating that miR-23a is affected by BMI. Moreover, increasing miR-23a expression may reduce the occurrence of PCOS. Murri et al. (2018) also demonstrated that miR-199a-3p and miR-199a-5p are negatively associated with BMI and the waist-to-hip ratio. Data by Arancio et al. (2018) show that serum miR-320 and miR-30a-3p levels are inversely correlated with low-density-lipoprotein cholesterol concentrations in women with hyperandrogenic PCOS. Therefore, these results indicate that miRNA may be a promising target for regulating lipid metabolism in PCOS. Nevertheless, the potential mechanism underlying lipid metabolism disorders has not been fully elucidated, thus, the therapeutic approach for PCOS has not generally taken dyslipidemia into consideration.
miRNA-Based Therapeutics
Currently, the clinical studies and guidelines regarding the currently available therapies are designed to overcome infertility and improve the prognosis of PCOS. Most of the drugs used for PCOS are intended to improve the symptoms of PCOS, including drugs that increase insulin sensitivity, antiandrogens, and induce ovulation, and have been approved for routine use for the treatment of PCOS (Radosh, 2009). Currently, miRNAs have attracted strong interest for improving the treatment approach and for evaluating the prognosis of PCOS, and investigations into the differential expression of miRNAs in PCOS may provide new tools for the development of miRNA-based therapeutics.
Improved insulin sensitivity may lead to enhanced glucose metabolism and decreased androgen levels, thereby improving fertility. Metformin, an insulin-sensitizing drug, has been used to improve IR and IGT in PCOS women over the last few decades. In addition, metformin can also reduce androgen secretion by controlling the ovary with respect to steroidogenic enzymatic activity (Lord et al., 2003). Although the effect of metformin on inducing ovulation and fertility in PCOS patients is still controversial, accumulative evidence suggests that metformin has the potential effect of improving the ovulation rate (Cho et al., 2009; Hadziomerovic-Pekic et al., 2010; Oppelt et al., 2010). In addition, administration of metformin downregulates miR-221 and miR-222 levels in type 2 diabetes patients (Coleman et al., 2013). Although it is not clear whether metformin improves IR and decreases androgen levels in PCOS patients by reducing the level of miR-222, miR-222 may provide new insight for investigations of the mechanism of metformin in the treatment of PCOS.
PCOS and its metabolic comorbidities are mainly due to a vicious cycle of androgen excess (Escobar-Morreale, 2018). In this context, strategies targeting HA will improve the signs and symptoms of PCOS. Antiandrogens and/or AR blockers are used to treat the symptoms of androgen excess in women with PCOS. miR-155 is negatively associated with androstenedione concentrations (Arancio et al., 2018). miR-27b plays a crucial role in controlling fatty acid metabolism and cholesterol homeostasis (Fernandez-Hernando et al., 2011). Sequential reverse antiandrogenic therapy (ethinyl-estradiol and cyproterone-acetate) induces increased levels of miR-27 b and miR-155 (Arancio et al., 2018). These suggest that miRNA may improve PCOS from the perspective of antiandrogen.
Perspectives
In summary, PCOS is a syndrome that affects a large number of women and has multiple types of clinical manifestations, which seriously affect the quality of life of many premenopausal women. Until now, no single criterion has been established for the diagnosis of PCOS, and the pathology of PCOS has not been easily generalized because it involves endocrinological, gynecological, and other factors. As a consequence, it is difficult to provide guidance to women and health professionals regarding PCOS. As such, the mechanisms of the anovulation, HA, IR, and lipid metabolism disorder in PCOS patients have not been fully elucidated. Therefore, the specific mechanism causing PCOS has become a research hotspot. Insight into the interplay between genetic regulation and the environment leading to differential miRNA expression may help elucidate the pathogenesis of PCOS.
Interestingly, based on the present study, it appears that miRNAs may serve as biomarkers that distinguish women with PCOS from those without PCOS. Moreover, differential miRNA expression, increased androgen levels, and reduced insulin sensitivity in PCOS patients are inextricably linked. The current understanding of the interrelation between miRNAs and PCOS development is limited, and the specific effects of miRNAs in PCOS development remain unclear because one miRNA may have numerous mRNA targets and because one mRNA may be controlled by several miRNAs. Our current knowledge of how miRNAs lead to increased GC proliferation and/or apoptosis, excessive folliculogenesis, abnormal androgen production, and a lipid metabolism imbalance is not yet complete. To the best of our knowledge, it is not possible to distinguish whether an altered miRNA expression profile is the cause or consequence of PCOS. Herein, further functional miRNA-PCOS studies are needed.
Although the exact underlying mechanism, by which insulin sensitizers and antiandrogenic therapy improve symptoms in women with PCOS, is largely uncertain, miRNA profiling can provide new insights into these molecular changes, thereby providing therapeutic information and improving prognosis. Nevertheless, there is no consensus regarding which strategy is the most effective for relieving PCOS symptoms, improving infertility and reducing long-term complications at present. Understanding these connections and the mechanisms underlying them can be fundamental factors for improving PCOS prognosis.
Footnotes
Acknowledgments
We appreciate the Horizontal cooperation project (2018KHX41), the Natural Sciences Foundation of Hunan Province (2019JJ40248, 2018JJ2344), the Scientific Research Innovation Program of postgraduate in Hu-nan Province (No. CX2018B623), the Key Lab for Clinical Anatomy & Reproductive Medicine of Hengyang City (2017KJ182), and the Chuanshan Talents Project in University of South China.
Disclosure Statement
No competing financial interests exist.