
Abstract
The mammary gland is an important organ for lactation in dairy goats. Mammary gland development and lactation functions are primarily regulated by natural hormones and certain crucial regulatory factors. Nedd4 family-interacting protein 1 (Ndfip1) can specifically bind to neural precursor cell-expressed, developmentally downregulated protein 4 (Nedd4) family members to participate in ubiquitination, which in turn regulates a range of biological processes in the body. However, the effects of Ndfip1 expression regulation at the post-transcriptional level on the development of mammary gland cells have not been previously reported. To study the regulation of Ndfip1 at post-transcriptional level, the overexpression and interference vectors of Ndfip1 were constructed, and co-transfected into the primary mammary gland epithelial cells cultured in vitro with miR-143 mimics and inhibitor. Dual luciferase reporter gene system, real-time quantitative polymerase chain reaction, western blotting, cholecystokinin octapeptide assays, and flow cytometry were used to identify their regulation and function. As a result, Ndfip1 was targeted and regulated by miR-143, which influences the development of mammary gland epithelial cells in dairy goats cultured in vitro. This study will lay an experimental foundation for further understanding the functions of Ndfip1 and miR-143.
Introduction
The mammary gland is a unique organ of dairy animals that begins its development in the embryonic stage and has varying degrees of growth before and after puberty. During pregnancy, mammary gland cells rapidly develop, proliferate, and differentiate, the number and volume of acini continuously increase. At the end of lactation, the secretory cells of the mammary gland begin to undergo apoptosis, and thereafter, it gradually develops into resting mammary gland. When pregnancy occurs, the mammary gland redevelops (Macias and Hinck, 2012; Inman et al., 2015). The entire physiological process of mammary gland development is primarily regulated through endocrine, paracrine, and autocrine mechanisms. Interactions between specific hormones, growth factors, and proteins play an important role in regulating mammary gland development and the physiological process of lactation.
Nedd4 family-interacting protein 1 (Ndfip1) is a multifunctional protein involved in ubiquitination, and it can bind specifically to neural precursor cell-expressed, developmentally downregulated protein 4 (Nedd4). Ndfip1 is considered an adaptor and activator protein in the Nedd4 protein family (Harvey et al., 2002; Liu et al., 2015). Recent studies have shown that the Nedd4 gene participates in regulation of many cellular processes, including endocytosis, the transforming growth factor-b signaling pathway, gene transcription, protein transport, degradation of defective proteins, and viral budding (Huang et al., 1999). Through binding to Nedd4, Ndfip1 is involved in protection of neurons against traumatic brain injury and cerebral ischemic neuronal damage, and by stimulating the peripheral T cell tolerance of itself and exogenous antigens, Ndfip1 regulates autoimmune diseases and allergic responses, and the pro-apoptotic effects of certain tumor cells (Putz et al., 2008; Yip et al., 2016; Tian et al., 2018). Other studies have shown that Ndfip1 overexpression can decrease the protein level of dopamine transporter (DAT), thus reducing 1-methyl-4-phenylpyridinium (MPP+) uptake and inducing apoptosis resistance, whereas the proteasome inhibitor MG132 can reverse Ndfip1-mediated inhibition of DAT protein. Ndfip1 can enhance the BCL-2/BAX ratio, reduce the release of cytochrome c from mitochondria to the cytoplasm and thus the associated cell damage, and exert an anti-apoptotic effect by reducing MPP+-induced Caspase-3 activation (Liu et al., 2015; Xing et al., 2016). Lack of Ndfip1 leads to defects in hematopoietic lineage cells and activation of T cells, thereby causing an increase in interleukin-4 (IL-4) production in vitro. Therefore, mice with insufficient Ndfip1 can have multiple skin and lung diseases, resulting in premature death (Layman et al., 2017). Ndfip1 is also involved in regulation of the susceptibility to gastrointestinal inflammation and inflammatory bowel disease mediated by T cells. Moreover, Ndfip1 has synergistic effects on inhibiting proliferation and facilitating apoptosis in human hepatocellular carcinoma SMMC-7721 cells, suggesting that Ndfip1 may promote apoptosis of tumor cells (Lv et al., 2011).
MicroRNAs (miRNAs) are endogenous small RNA molecules ∼21–24 nucleotides (nt) in length that do not encode for proteins. miRNA is widely involved in many biological activities in animals, plants, and microorganisms, including reproduction, development, viral defense, cell proliferation, apoptosis, and fat metabolism (Bartel, 2004; Wienholds and Plasterk, 2005). In the mammary gland, miRNA can regulate post-transcriptional expression of target genes and thereby control the formation of mammary ducts and acini and the proliferation, differentiation, and apoptosis of mammary gland epithelial cells. These activities in turn influence mammary gland development and lactation (Ji et al., 2013, 2015; Wang et al., 2015; Chen et al., 2018). During differentiation and degeneration of the mammary gland in small mice, the expression level of miR-101a continuously varies, and miR-101a overexpression can downregulate the expression of β-casein, a marker of mammary gland epithelial cell differentiation, and concurrently inhibit cell growth. Additional studies have found that miR-101 regulates mammary gland development by downregulating the expression of the cyclooxygenase gene, which promotes cell proliferation (Tanaka et al., 2009). Furthermore, miR-212 and miR-132 are mammary duct-specific miRNAs. When the expression levels of miR-212 and miR-132 are inhibited, the matrix metalloproteinase content increases, and a large number of matrix metalloproteinases surround the mammary duct, thus preventing its formation (Ucar et al., 2010).
In our previous studies, we found that chi-miR-143-3p can inhibit proliferation and promote apoptosis of primary dairy goat mammary gland epithelial cells cultured in vitro (Ji et al., 2016). Other research has shown that post-transcriptional regulation of Ndfip1 only occurs in uveal melanoma cells. miR-155 can promote the growth and invasion of uveal melanoma cells by inhibiting the expression of Ndfip1, whereas the original effects of miR-155 on these cells can be restored by Ndfip1 overexpression (Peng et al., 2017). miRNA regulation of Ndfip1 in dairy goat mammary gland epithelial cells has not been reported. In this study, based on the bioinformatics analysis of the target gene, dual luciferase reporter gene system, real-time quantitative polymerase chain reaction (qRT-PCR) and western blotting, the targeted regulation of Ndfip1 by chi-miR-143-3p was addressed, and effects of Ndfip1 on cell development were also certified using CCK-8 and flow cytometry (FCM). These results illustrated that miR-143 can inhibit the proliferation and promote their apoptosis of mammary gland epithelial cells by targeting and regulating the expression of Ndfip1, which may provide a theoretical basis for the functional roles of chi-miR-143-3p and Ndfip1 in dairy goat mammary gland epithelial cells cultured in vitro.
Materials and Methods
Animals and ethical statement
Laoshan dairy goats from Laoshan dairy goat primary farm in Qingdao, Shandong Province, China were used, which were 4-year-old, third lactation period, feed and managed in the same environment. The mammary gland tissue samples were collected by surgery under general anesthesia.
The Committee of the Management and Use of Laboratory Animals of Shandong Agricultural University decided to approve the use of goat tissues for experiment studies. The laboratory is required to strictly abide by the Regulations on the Administration of Laboratory Animals of Shandong Agricultural University. All personnel (including teachers, experimenters, postdoctoral students, doctoral students, and master's students) who operate experiment animals must be trained. All animals must be normally raised in animal houses under the supervision of the Laboratory Animal Center of Shandong Agricultural University. The principles of animal welfare should be strictly observed in the operation and execution of animals (SDAUA-2018-40).
Culture and transfection of primary mammary gland epithelial cells from dairy goats
The mammary gland tissue of dairy goats was surgically resected, washed with phosphate-buffered saline (PBS; supplemented with 1% antibiotic/antimycotic solution), and then cut into milky granules using ophthalmic scissors. Primary dairy goat mammary gland epithelial cells were obtained by tissue block culture at 37°C in 5% CO2. The culture medium used was Dulbecco's modified Eagle's medium/Ham's F-12 nutrient mixture (DMEM/F-12) supplemented with 10% fetal bovine serum, 0.25 mM hydrocortisone, 5 mg/mL insulin, 50 U/mL penicillin/streptomycin, and 10 ng/mL epidermal growth factor 1.
The cultured primary mammary gland epithelial cells were inoculated in six-well cell culture plates. When cell confluence reached 65% or higher, the cells were transfected using Lipofectamine™ LTX and PLUS™ Reagent (Invitrogen). Three duplicate wells were prepared for each transfection group.
Total RNA extraction and qRT-PCR detection
In this experiment, total RNA was extracted from the dairy goat primary mammary gland epithelial cells using an OMEGA MicroElute Total RNA Kit (Omega). miRNAs were reverse-transcribed into cDNA according to the instructions provided with a SYBR® PrimeScript™ miRNA RT-PCR Kit (Takara, Japan). The reaction was carried out at 37°C for 1 h and 85°C for 5 min. Messenger RNAs (mRNAs) were reverse-transcribed into cDNA according to the instructions of a SYBR PrimeScript miRNA RT-PCR Kit (Takara).
Real-time quantification of miRNA and mRNA expression levels was performed using a Mx3000® SYBR™ Green qPCR analyzer. According to the instructions for SYBR® Premix EX Taq™ II Tli RNaseH Plus (Takara), the qPCR primer for miR-143-3p was 5′-TGAGATGAAGCACTGTAGCTCGi-3′, and the U6 primers were 5′-CTCGCTTCGGCAGCACA-3′ and 5′-AACGCTTCACGAATTTGCGT-3′. According to the instructions for SYBR Premix EX Taq Tli RNaseH Plus (Takara), the qPCR primers for Ndfip1 mRNA were 5′-TGCTCCTCCACCTTACAG-3′ and 5′-ATCCCATCGTTTCCTATC-3, and the qPCR primers for GAPDH were 5′-CACCCTCAAGATTGTCAGC-3′ and 5′-CAGTGGTCATAAGTCCCTCC-3′. All reagents were prepared on ice, and each reaction was repeated three times.
Construction of the dual luciferase reporter gene vector
Based on the Ndfip1 sequence in the National Center for Biotechnology Information (NCBI) database, PCR primers (5′-CCGCTCGAGATGCCTTATCTATTGTTT-3′ and 5′-ATAAGAATGCGGCCGCCTATGAGTTTGCTTCCAG-3′) for the 3′ untranslated region (UTR) of Ndfip1 were designed with the addition of NotI and XhoI double-digest restriction sites and protective bases at both ends. After PCR amplification and purification, these primers were subcloned into psiCHECK2, which was then subjected to double digestion and sequencing. A wild-type dual luciferase reporter gene vector based on Ndfip1 was constructed and named psiCHECK2-Ndfip1-Wt-3′UTR. Using the wild-type vector sequence as a template, the site-directed mutagenesis primers 5′-GGCAATCTTGCTTTTATTCATCCTCCGGCGCAAAATGAATTTGG-3′ and 5′-AAATTCATTTTGCGCCGGAGGATGAATAAAAGCAAGATTGCC-3′ were designed. A mutant dual luciferase reporter gene vector based on Ndfip1 was constructed and named psiCHECK2-NDFIP1-Mut-3′UTR.
Construction of Ndfip1 overexpression and interference vectors
Based on the single coding sequence (CDS) of Ndfip1 in the NCBI database, the overexpression primers 5′-AGCTTTGTTTAAACGCTCACTACCCGAGGAGA-3′ and 5′-ATAAGAATGCGGCCGCTGTGTTCCGCATCTTGAT-3′ were designed. The PCR product was double-digested with PmeI and NotI and then ligated into the recombinant plasmid pEZ-M02 (GeneCopoeia) to construct a Ndfip1 overexpression vector, which was named pEZ-M02-Ndfip1. The interference vector was synthesized by Guangzhou Ruibo Biological Technology Co., Ltd.
Activity detection of dual luciferase reporter gene assay system
After 48 h of transfection, cells were mixed with 20 μL of lysis buffer. According to the instructions of the Dual-Luciferase® Reporter Assay System (Promega), the supernatant was mixed with 20 μL of firefly luciferase assay reagent, after which firefly luciferase activity was measured. Then, 20 μL of Renilla luciferase assay working solution was added, and after thorough mixing, Renilla luciferase activity was measured. Using Renilla luciferase as the internal reference, the relative activity values of the two luciferases were used to calculate the luciferase expression activity value (Renilla luciferase activity value/firefly luciferase activity value) to determine the strength of luciferase expression activity. Each assay was repeated three times. The mimics and inhibitors of miR-143 were ordered from Guangzhou Ruibo Biological Technology Co., Ltd.
Western blotting to detect protein expression
When the confluency of the dairy goat mammary gland epithelial cells reached 70–80%, cultured cells in the transfection groups were transfected separately. After 24–48 h, an appropriate amount of lysis buffer was added to obtain cell lysates and extract proteins. Protein quantification was performed using the bicinchoninic acid (BCA) method. After preparation of sodium dodecyl sulfate–polyacrylamide gel electrophoresis (SDS-PAGE) stacking and separating gels, 25 μL of the denatured protein sample and 10 μL of the prestained protein marker were added to the corresponding wells. Electrophoresis was performed at 80 V, which was changed to a constant voltage of 100 V after 1 h. When bromophenol blue reached the bottom of the rubber plate, a constant current of 200 mA was maintained for protein transfer. After 2 h, the polyvinylidene fluoride (PVDF) membrane was blocked in blocking solution and incubated with primary antibody (Rabbit Anti-NFIP1 polyclonal antibody, bs-19058R; Bioss, Beijing) overnight at 4°C. After rinsing, the PVDF membrane was incubated with horseradish peroxidase-labeled secondary antibody at room temperature for 2 h. Protein expression was detected using enhanced chemiluminescence, and the results were analyzed.
CCK-8 assay to detect cell proliferation
Primary dairy goat mammary gland epithelial cells in logarithmic growth phase were obtained, digested with 0.25% trypsin, and inoculated into 96-well culture plates. Each group consisted of four wells, and cells were cultured for 48 h at 37°C until adhesion. The optical density (OD) value of each well at 490 nm was measured at 0, 12, 24, 36, and 48 h after transfection. Cell viability was measured as follows: [(OD value of cells in transfected group − OD value of blank cells)/(OD value of cells in control group − OD value of blank cells)] × 100%.
FCM to detect cell apoptosis
After 48 h of transfection, primary dairy goat mammary gland epithelial cells in each group were collected and rinsed with PBS. Then, 500 μL of binding buffer was added to prepare a single-cell suspension. Next, 5 μL annexin V-FITC and 5 μL propidium iodide (PI) were successively added to the suspension, which was mixed thoroughly. The reaction was conducted in the dark at room temperature for 10 min. FCM (BD, Franklin Lake, NJ) was used to detect apoptosis in each group. Statistical analysis of the raw data was performed using Flowjo 7.6.1 software (Ashland, OR).
Statistical analysis of data
Raw qPCR data were processed using the 2−ΔΔCt method. Experimental data were analyzed using SPSS 20.0 software. All the results are expressed as mean ± standard error of the mean. Data from multiple groups were compared by one-way analysis of variance. Pairwise comparisons in each group were performed using a least significant difference t-test. Statistical significance was assessed at p < 0.05.
Results
chi-miR-143-3p regulates Ndfip1 expression in dairy goat primary mammary gland epithelial cells cultured in vitro
To identify the potential target genes of chi-miR-143-3p, we used the TargetScan 7.0 database and Pictar to predict putative target genes. By examining the overlap in the prediction results from the two software programs, Ndfip1 was chosen as a potential gene target for this study (Fig. 1). There is a potential 7mer-A1 target site at position 329 in the 3′UTR of Ndfip1 with a potential energy of −23.5 kcal/mol that is highly conserved among species.
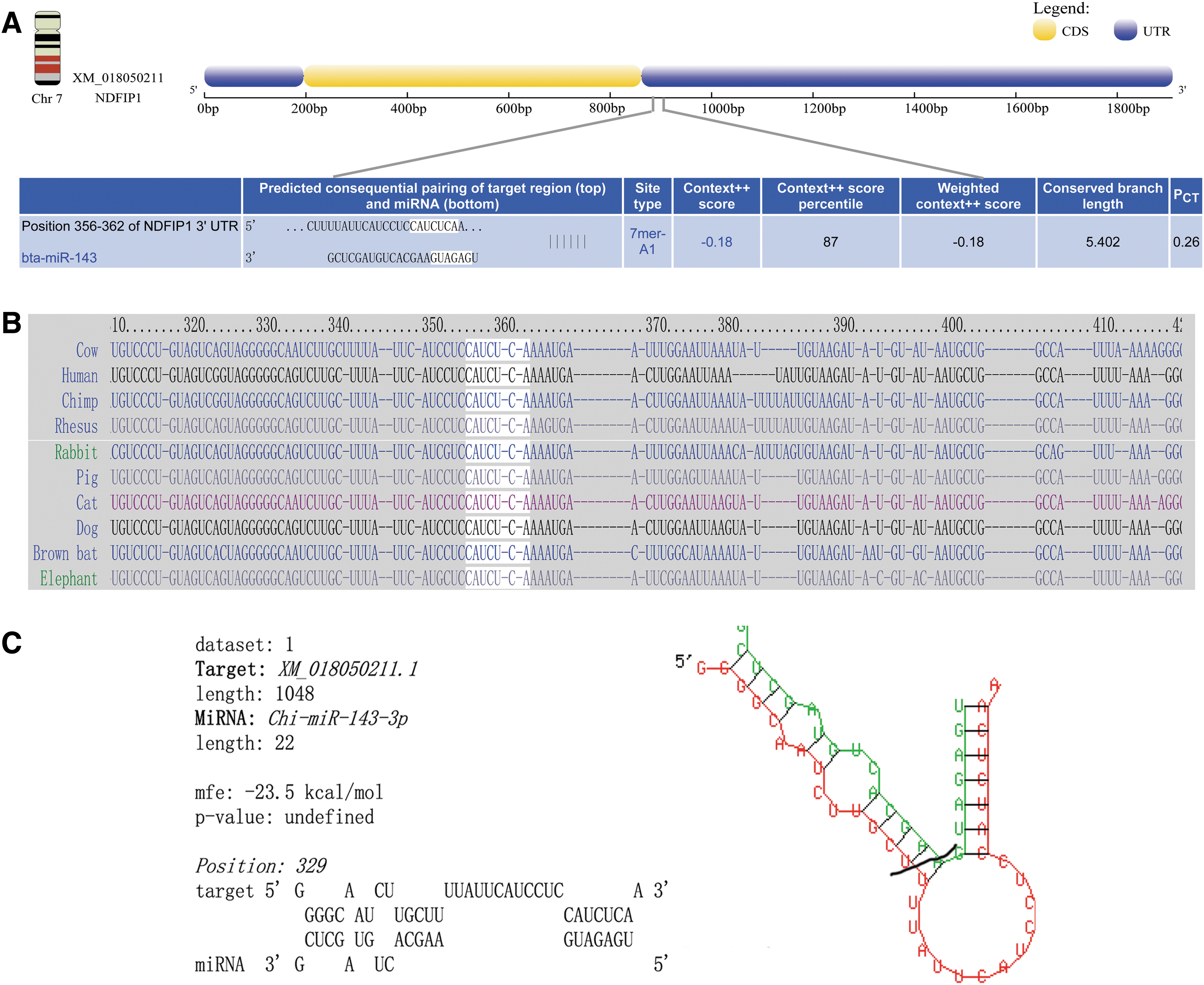
The 3′UTR of Ndfip1 existed a potential target site.
To further verify whether chi-miR-143-3p regulates Ndfip1 expression, chi-miR-143-3p was separately overexpressed and inhibited in mammary gland epithelial cells from dairy goats. qRT-PCR and western blotting were used to detect Ndfip1 expression. As given in Fig. 2, compared with the two control groups, NC and Mock, cells transfected with chi-miR-143-3p mimic and inhibitor showed successful overexpression and inhibition of chi-miR-143-3p, respectively (p < 0.01) (Fig. 2A). After chi-miR-143-3p overexpression, the mRNA and protein expression levels of Ndfip1 significantly decreased relative to those in the control group (p < 0.05) (Fig. 2B, C), and after chi-miR-143-3p inhibition, the mRNA and protein expression levels of Ndfip1 significantly increased relative to those in the control group (p < 0.05) (Fig. 2B, C). These results indicate that chi-miR-143-3p is a potential regulator of Ndfip1 expression at both the transcriptional and protein level.
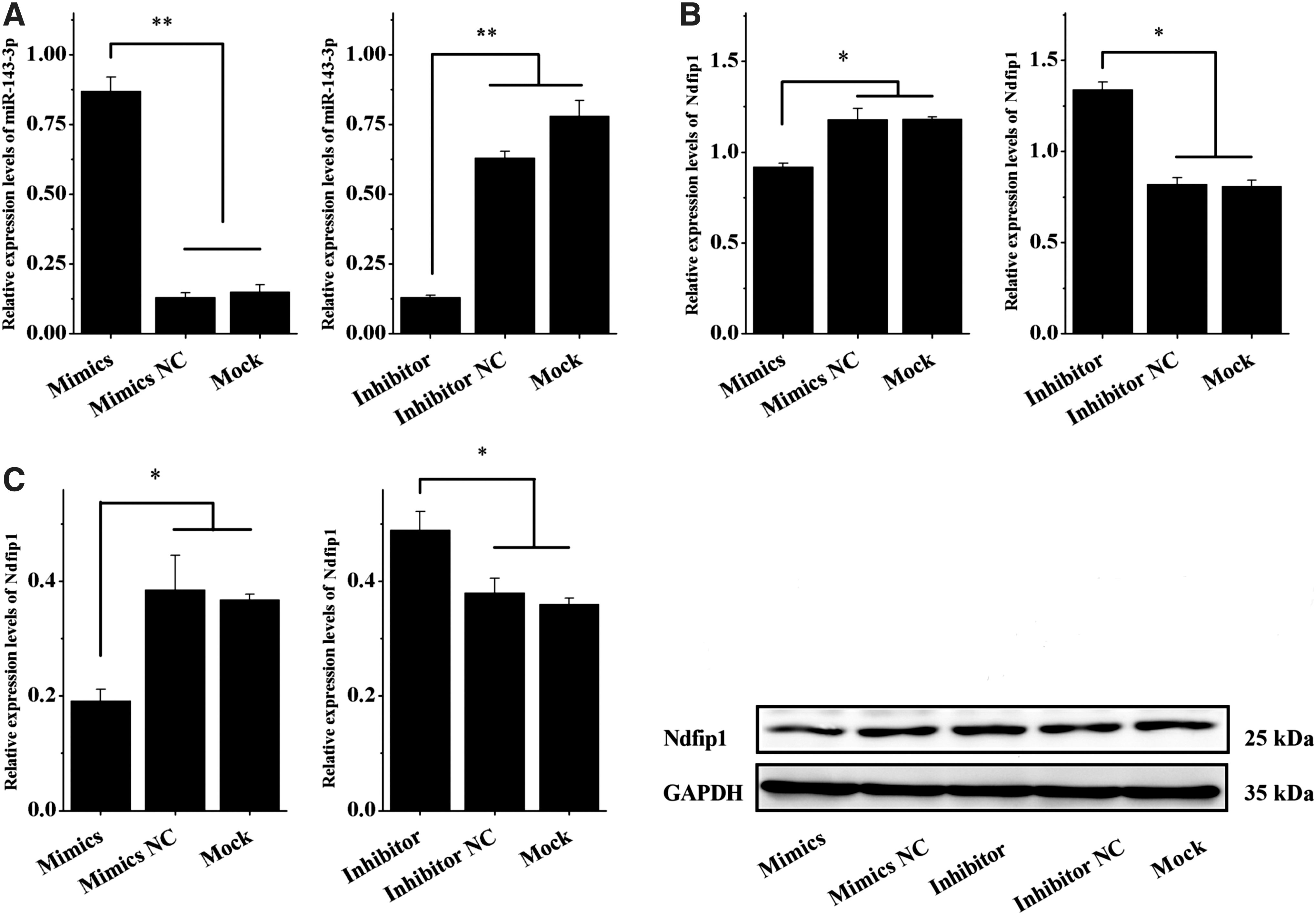
chi-miR-143-3p inhibited Ndfip1 expression at mRNA and protein levels.
A regulatory target of chi-miR-143-3p exists in the 3 ′UTR of Ndfip1
To further verify whether the target located in the 3′UTR of Ndfip1 is a true target of chi-miR-143-3p, we co-transfected primary dairy goat mammary gland epithelial cells with chi-miR-143-3p mimic and constructed the wild-type eukaryotic expression vector psiCHECK2-NDFIP1-Wt-3′UTR, along with chi-miR-143-3p mimic NC and mutant eukaryotic expression vector psiCHECK2-NDFIP1-Mut-3′UTR. A dual luciferase reporter assay showed that after co-transfection of the primary dairy goat mammary gland epithelial cells with chi-miR-143-3p mimic and psiCHECK2-NDFIP1-Wt-3′UTR, the luciferase activity in the cells significantly decreased relative to that in the other transfection groups (p < 0.05) (chi-miR-143-3p mimic + psiCHECK2-NDFIP1-Mut-3′UTR, chi-miR-143-3p mimic NC + psiCHECK2-NDFIP1-Wt-3′UTR, and chi-miR-143-3p mimic NC + psiCHECK2-NDFIP1-Mut-3′UTR) (Fig. 3). These results indicate that chi-miR-143-3p altered the activity of the luciferase reporter gene system by binding to the 3′UTR of Ndfip1. Bioinformatically predicted as a potential target, the 3′UTR of Ndfip1 is a true regulatory target of chi-miR-143-3p.
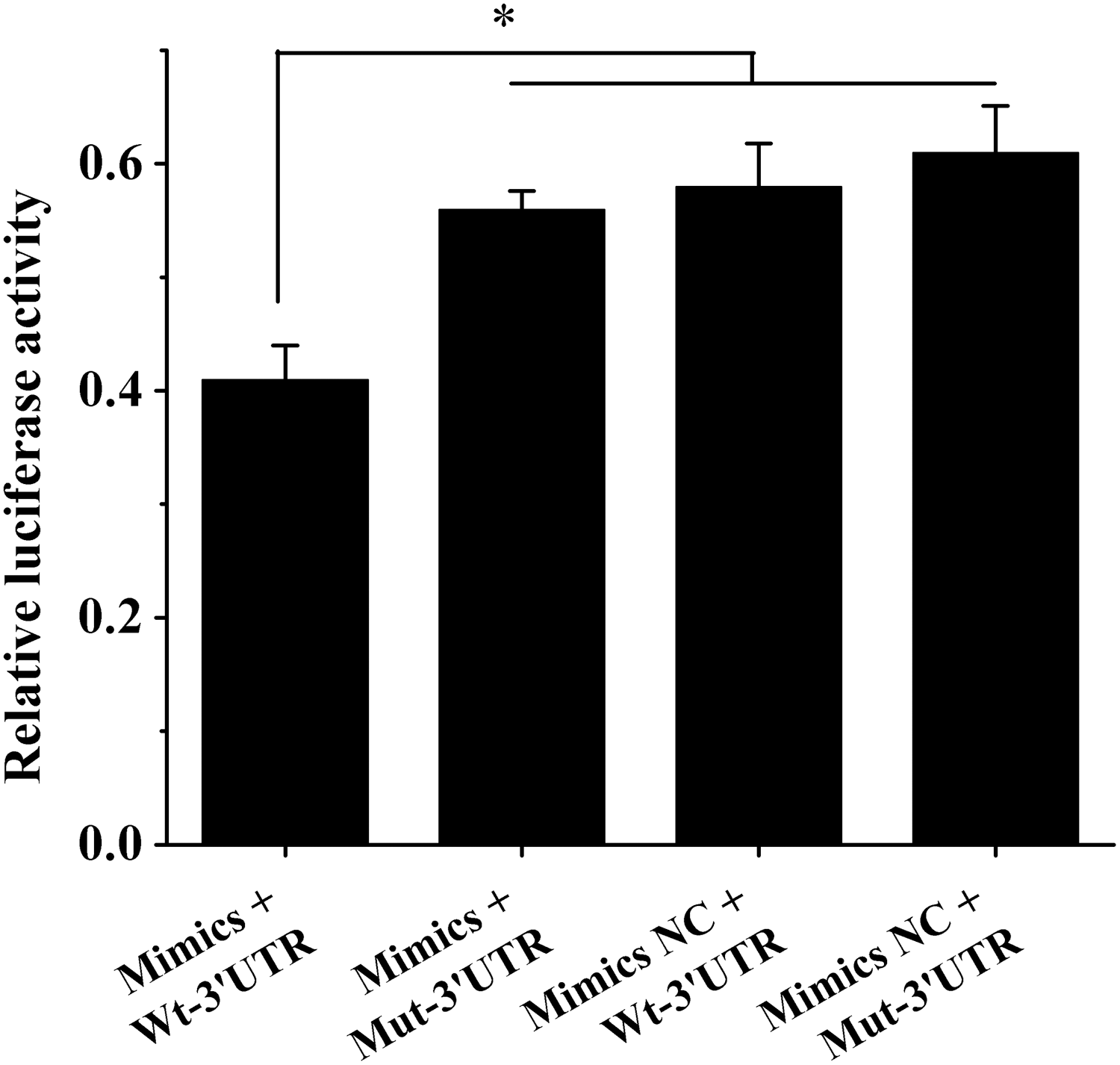
The results of double luciferase activity in different treatment groups. The luciferase activity was reduced after the co-transfection of chi-miR-143-3p mimics and psiCHECK2-NDFIP1-Wt-3′UTR. *A significant difference between different treatments (p < 0.05).
Ndfip1 promotes proliferation of primary dairy goat mammary gland epithelial cells cultured in vitro and inhibits their apoptosis
To study the effects of Ndfip1 on primary dairy goat mammary gland epithelial cells cultured in vitro, we separately transfected the cells with the constructed Ndfip1 overexpression and interference vectors. After 24–48 h, cholecystokinin octapeptide (CCK-8) and FCM assays were used to detect cell proliferation and apoptosis, respectively. The CCK-8 assay results showed that the number of viable cells differed between cells transfected with pEZ-M02-ndfip1 and control group cells after 36 h of transfection. At 24 h, the number of viable cells in the transfected group was significantly higher than that in the control group (p < 0.05) (Fig. 4A). The difference in cell viability was significant between the two groups at the 36 and 48 h time points, indicating that Ndfip1 promoted the proliferation of mammary gland epithelial cells. When cells were transfected with si-Ndfip1, the number of viable cells in the transfected group was significantly lower than that in the control group after 24 h of transfection, and the difference between the two groups at 36 and 48 h was significant (p < 0.05) (Fig. 4B). These findings show that interfering with Ndfip1 expression inhibits proliferation of mammary gland epithelial cells from dairy goats.
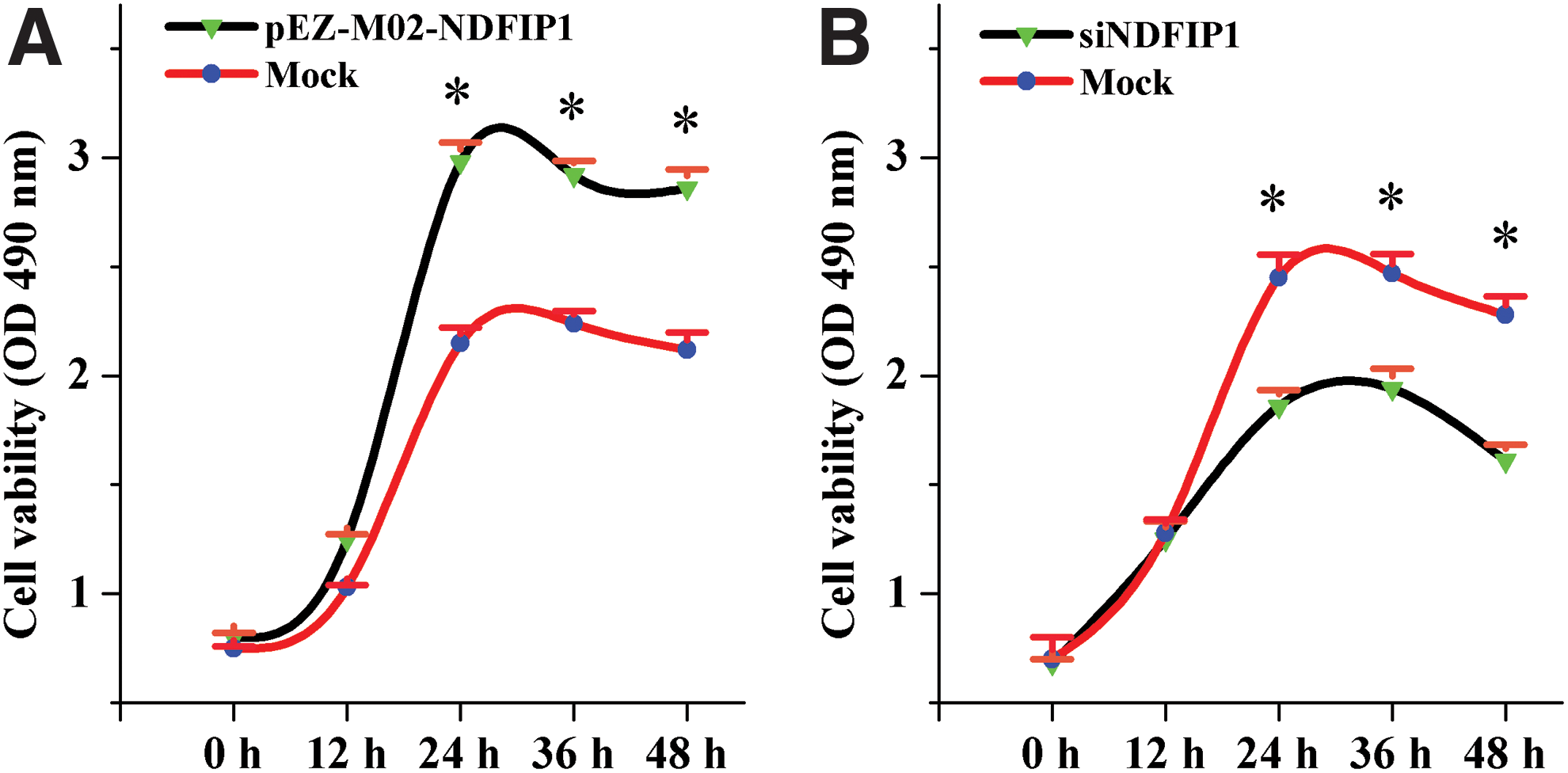
Cell viability was measured by CCK-8 assay after transfecting the pEZ-M02-NDFIP1 or si-NDFIP1.
Cells transfected with the Ndfip1 interference vector were collected after 24 h. Apoptosis was determined using FCM and double staining with PI/annexin V-FITC. The results showed that early-stage, late-stage, and total apoptosis rates in mammary gland epithelial cells transfected with the Ndfip1 interference vector were significantly higher than those in control cells (p < 0.05) (Fig. 5).
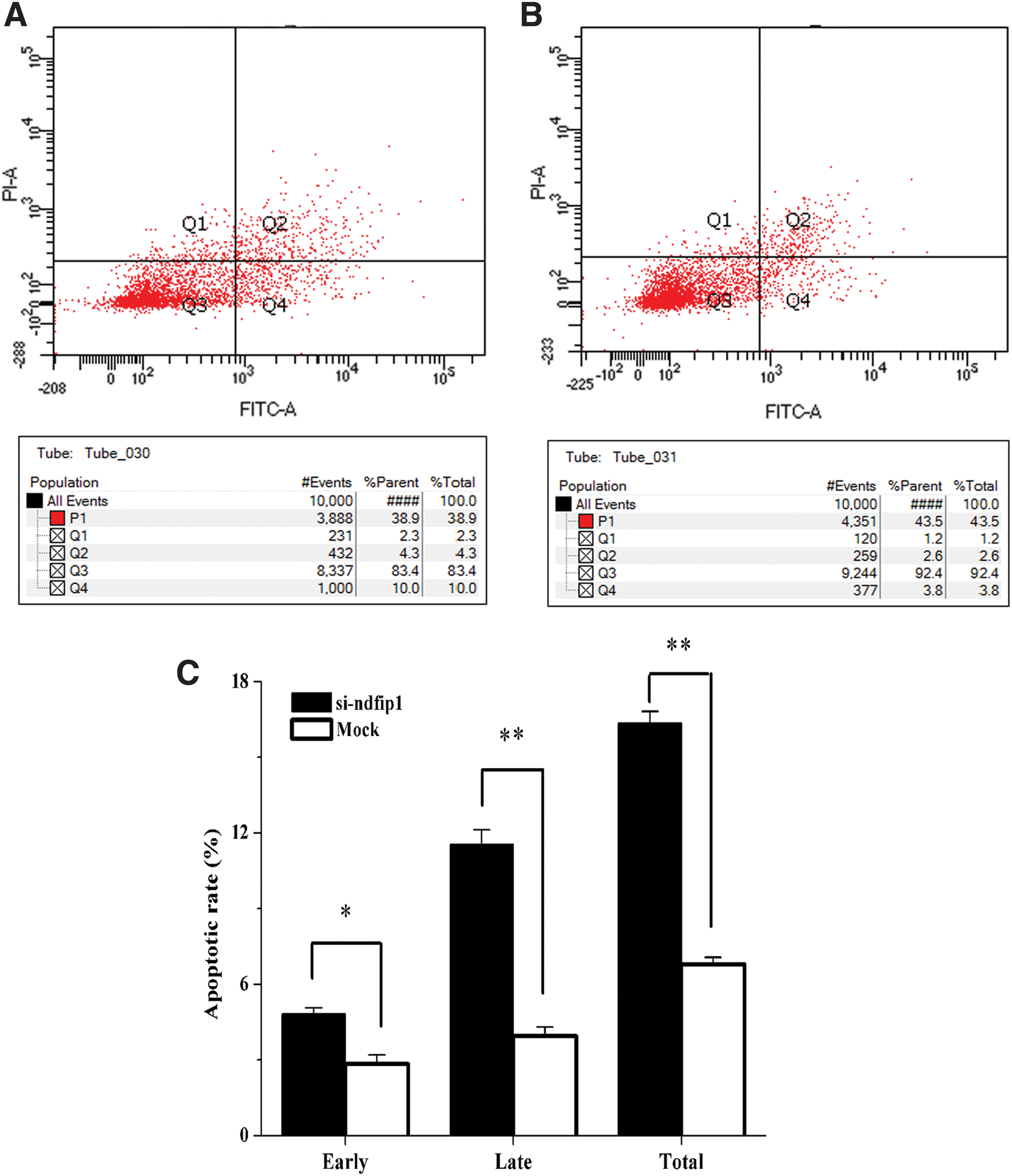
Inhibiting Ndfip1 expression promoted the apoptosis of dairy goat mammary gland epithelial cells cultured in vitro.
Discussion
Mammary gland development in dairy animals determines their lactation performance, which is an important factor affecting the quality of dairy products (Silanikove et al., 2014). Dairy goats are an important dairy animal species with a global distribution and have an increasingly important economic role because of the unique nutritional and health properties of their dairy products. Mammary gland development in dairy goats is the same as that in other dairy animals; development begins in the embryonic stage, and the gland grows allometrically before puberty and intensely develops during pregnancy (Rowson, et al., 2012; Davis, 2017). At different mammary gland development stages, mammary gland epithelial cells differentiate, divide, grow, and develop, and repeated cycles of cell proliferation, apoptosis, and programmed death occur. These processes determine the number, structure, activity, and other properties of the cells, which in turn influence the physiology and morphological characteristics of the mammary gland and its lactation performance (Macias and Hinck, 2012; Inman et al., 2015; McNally and Stein, 2017). Numerous studies have suggested that mammary gland development and lactation are primarily regulated by the endocrine system, which involves many types of hormones, including growth hormones, estrogen, progesterone, prolactin, and insulin-like growth factors. These hormones maintain a stable internal environment and collectively influence mammary gland growth and development and physiological processes, such as milk synthesis, secretion, and lactation (Schams et al., 1984; Berryhill et al., 2016).
Completion of physical maps of the human genome and determination of genomic sequences of other organisms has provided basic information providing insight into the nature of life at the genetic level. Studies of specific gene functions provide effective means for investigation of physiological changes in different types of economic animals (Li and Deng, 2018; Yang et al., 2019). Li et al. (2012) first used high-throughput transcriptome sequencing to analyze the miRNA expression profile of mammary gland tissue from Xinong Saanen dairy goats (Capra hircus) during the dry period and lactation peak. That study opened a new chapter in using transcriptome sequencing technology in goat research. Subsequently, the technology has been widely used in studies of goat skin, hair follicles, muscles, ovaries, lymph nodes, liver, and other organs, and in studies of blood and milk from goats and transgenic goats (Yuan et al., 2013; Su et al., 2015; Zhao et al., 2015; Golan-Gerstl et al., 2017; Manjunath et al., 2017; Chen et al., 2018). Gene expression can change among different cell types, differential physiological stages of the same cell, and different external conditions. Moreover, a small number of genes are selectively and specifically expressed, and this specific expression regulates the occurrence, growth, and developmental processes of individuals, which ultimately controls the morphology and physiological changes in organisms. Our research group previously analyzed transcriptome and miRNA expression profiles of mammary gland tissues from dairy goats in the early, peak, and late stages of lactation. We identified and screened genes and miRNAs differentially expressed at different stages, annotated and analyzed their functions, and successfully established a gene regulation network for development of the dairy goat mammary gland and dairy goat lactation. These studies provide theoretical and technical support for studying the regulation mechanisms underlying development of the mammary gland in dairy goats and lactation of dairy goats (Ji et al., 2012a, 2017, 2019).
Through cell counting, 3-[4,5-dimethylthiazol-2-yl]-2,5-diphenyltetrazolium bromide (MTT) assays, Hoechst 33342/PI double staining, FCM, and other techniques, previous studies from our research group found that miR-143-3p altered the cycles of dairy goat mammary gland epithelial cells cultured in vitro to inhibit cell proliferation and promote apoptosis. In addition, variations in miR-143-3p expression alter the mRNA expression of the cell proliferation/apoptosis marker BAX/BCL-2 (Ji et al., 2016). However, the mechanisms by which miR-143-3p regulates the growth of dairy goat mammary gland epithelial cells cultured in vitro remain unclear. Based on analyses of miRNA expression profiles and transcriptome data at different lactation stages, our previous study found that miR-143-3p was somewhat associated with Ndfip1 expression (Ji et al., 2012b, 2017, 2019).
To further investigate whether miR-143-3p and Ndfip1 have regulatory effects, a potential target of miR-143-3p was revealed to exist in the 3′UTR of Ndfip1 through bioinformatics predictions. Through analysis of Ndfip1 mRNA and protein expression levels through qRT-PCR and western blotting after miR-143-3p overexpression and inhibition, miR-143-3p was shown to regulate Ndfip1 expression in dairy goat mammary gland epithelial cells cultured in vitro (p < 0.05) (Fig. 2). Furthermore, miR-143-3p decreased Ndfip1 3′UTR dual luciferase activity (p < 0.05) (Fig. 3). These findings indicate that the potential target in the 3′UTR of Ndfip1 is a true regulatory target of miR-143-3p. Nevertheless, analysis of the functions of transfected dairy goat mammary gland epithelial cells cultured in vitro showed that Ndfip1 overexpression promoted cell proliferation, while reducing its expression inhibited cell proliferation (p < 0.05) (Fig. 4). FCM results also demonstrated that interfering with Ndfip1 expression enhanced the rate of early-stage, late-stage, and total apoptosis of mammary gland epithelial cells cultured in vitro (p < 0.05) (Fig. 5). Combined with the previous research findings concerning miR-143-3p (Ji et al., 2016), we hypothesize that miR-143-3p is involved in proliferation and apoptosis of mammary gland epithelial cells cultured in vitro through regulation of Ndfip1 expression. However, determining the mechanisms by which miR-143-3p regulates dairy goat mammary gland epithelial cells cultured in vitro requires further study.
Footnotes
Disclosure Statement
No competing financial interests exist.
Funding Information
This work was supported by the National Natural Science Foundation of China (31672401), Shandong Provincial Natural Science Foundation of China (ZR2014CM029), a project of the Shandong Province Higher Educational Science and Technology Program of China (J14LF07), the Shandong Provincial Modern Agriculture Industry Technology System (SDAIT-10), and funds of the Shandong “Double Tops” Program (SYL2017YSTD12).