
Abstract
Growth differentiation factor (GDF) 9 gene is involved in regulating reproductive traits in animals, but little is known about the promoter, single-nucleotide polymorphisms (SNPs), transcription factor binding sites, and regulating mechanism of GDF9 gene. In this study, the SNPs in the GDF9 promoter region were explored and their transcription mechanisms in regulating GDF9 expression were analyzed. Ear tissues of 267 Hu ewes were collected, and genomic DNA was extracted. GDF9 promoter region was amplified by PCRs, and identified SNPs genotyped by sequencing. SPSS16.0 software was used to analyze the association between genotypes and litter sizes. Flow cytometry assay was used to detect cell apoptosis, and dual-luciferase reporter assay was used to discover the promoter activity. A length of 1789 bp promoter region of GDF9 in Hu sheep was obtained by PCR amplification, and luciferase activity assay showed that there was a negative regulatory element in the region within −725 to −309 bp and a positive regulatory element in the region within −309 to +43 bp. Three complete linkage SNPs at −534A/G, −407T/G, and −332C/T were detected, resulting in three genotypes (namely, AA, AB, and BB). The association analysis indicated that the AA genotype ewes had larger litter size at average parity than those with the BB genotype. The −534A/G mutation created a novel binding site for the octamer transcription factor 1 (OCT1), and the Annexin V FITC/PI flow cytometry assay showed OCT1 promoted cell apoptosis in sheep ovarian granulosa cells. Overexpression of OCT1 considerably inhibited the luciferase activity of both genotypes and the inhibition effect of pGL3-BB was higher than that of pGL3-AA. Three complete linkage SNPs of the GDF9 gene regulate the litter size in Hu sheep probably via inhibition of the promoter activity by binding with OCT1 at −534 GG genotype and forming a complex between OCT1 and CCAAT/enhancer-binding protein (C/EBP).
Introduction
Growth differentiation factor (GDF) 9, first identified in the human ovary, is a member of the transforming growth factor β superfamily and an important growth and differentiation factor during early folliculogenesis (Juengel et al., 2004). It has been widely studied owing to its high and specific expression in ovarian tissue (McPherron and Lee, 1993; Hsueh et al., 2000). GDF9 mRNA is synthesized only in the oocyte from the primary 1-layer follicle stage until ovulation (Incerti et al., 1994). Loss of function of GDF9 during early follicular development resulted in infertility in female mice (McGrath et al., 1995); higher mature GDF9 levels in the follicular fluid were significantly correlated with oocyte nuclear maturation and embryo quality (Gode et al., 2011).
Dong et al. (1996) found that GDF9−/− female mice completely lost reproductive ability. GDF9 can regulate follicular growth and differentiation, facilitating granulosa cell (GC) proliferation and maintaining follicular microenvironment stability and cooperative action along with bone morphogenetic protein 15 (BMP15), which are essential for ovulation, fertilization, and female reproduction (Vitt et al., 2000). GDF9 promotes follicle-stimulating hormone-induced progesterone production, regulates the transcription of related genes, and inhibits the apoptosis of chicken follicular GCs (Li et al., 2019).
The ovine GDF9 gene is located on chromosome 5 and comprises two exons and one intron. In sheep, GDF9, along with bone morphogenetic protein receptor type 1B (BMPRIB) and BMP15, has been shown to have a considerable effect on litter size and ovulation rate (Davis, 2005). Studies have revealed polymorphism of GDF9 and its association with litter size in sheep, but most studies are related to the coding region (Paz et al., 2015; EI-Seedy et al., 2017; Abdoli et al., 2018). Bravo et al. (2016) found eight single-nucleotide polymorphisms (SNPs) in Araucana creole sheep (ACS), and reported c.978A>G and c.994G>A SNPs as genetic markers of fecundity in ACS. A novel synonymous A/G mutation at 41, 769, 471 of GDF9 was found to be closely associated with the litter size in Hu sheep (Wang et al., 2018). So far, no promoter polymorphisms of the GDF9 gene have been detected in prolific sheep such as Hu sheep, and the molecular mechanisms regulating the GDF9 gene expression in sheep remain poorly understood.
In the present study, with the native Hu sheep as the subject, the SNPs in the GDF9 promoter region were explored and their mechanisms in regulating the expression of GDF9 were analyzed. The promoter sequence was amplified, and its characteristics in the GDF9 gene were explored, its SNPs were screened, its correlation with litter size was analyzed, and its molecular mechanism was clarified.
Materials and Methods
Sampling and DNA extraction
The ear tissue of 267 Hu ewes (independent individuals and there is no relationship among them) in a sheep farm of Xilaiyuan sheep Industry Co., Ltd., (Taizhou, China) was collected to extract DNA using the standard phenol–chloroform extraction protocol (Psifidi et al., 2010). All experiments were performed in accordance with the protocol approved by the Committee on Ethics of Animal Experimentation from the Jiangsu Academy of Agricultural Sciences (No. 63 of Jiangsu Academy of Agricultural Sciences, approved on July 8, 2014).
Cloning, sequencing, and genotyping
PCRs were performed using a mixture of total volume 50 μL containing 1 μL reverse transcription product or 60 ng DNA, 0.25 μL 5 U/μL Taq (Takara, Japan), 5 μL 10 × buffer (Mg2+ plus), 4 μL dNTP mixture, and 1 μL 10 nM upstream and downstream primers (Table 1). The PCR conditions used for the amplification of the GDF9 and octamer transcription factor 1 (OCT1) gene were as follows: 10 s at 98°C for denaturation, 30 cycles of amplification (10 s at 98°C, 30 s at 55°C, 1 min at 72°C), and 5 min at 72°C for extension. The PCR production was separated using 1% agarose gel electrophoresis, and the expected size was purified and sequenced in Qingke Biological Technology Co., Ltd. (Nanjing, China).
Primer Sequences and the Parameters of PCR in this Study
Underline indicates the restriction enzyme sites.
GDF9, growth differentiation factor 9; OCT1, octamer transcription factor 1.
The GDF9 promoter sequences were amplified to screen SNPs using the DNA pooling sequencing assay with P1 and P2 (Table 1). Five microliters of 100 ng/μL DNA collected from five ewes were pooled. The PCR products were sequenced in both the directions to identify SNPs using Chromas v2.32 (Technelysium, Australia). The SNPs were genotyped by sequencing.
Bioinformatic analysis
The GDF9 promoter sequences were analyzed using DNASTAR program (DNASTAR, Inc., Madison, WI). The core promoter was predicted using Promoter 2.0 software (Steen Knudsen, 1999), CpG islands were predicted using the MethPrimer software, and the transcription binding sites in the promoter were predicted using JASPAR online software.
Plasmid construction
The Hu sheep GDF9 promoter vectors were constructed by PCR cloning. Three specific primers (P3, P4, and P5, Table 1) containing NheI and HindIII restriction enzyme sites were designed to amplify GDF9 promoter deletion fragments (1: −309 to +43 bp; 2: −725 to +43 bp; 3: −1239 to +43 bp), and the PCR products were first cloned into pMD19-T vector (Takara) and then digested with corresponding plasmids and pGL3-basic with NheI and HindIII enzymes (Takara) to create pGL3-1 (−309∼+43), pGL3-2 (−725∼+43), and pGL3-3 (−1239∼+43) products. To identify the promoter activity, the sequences were amplified using the −534 GG and −534 AA genotype ewes' DNA as the template, amplified with P6 (Table 1) and obtained −534 GG and −534 AA GDF9 promoter fragments, and then connected them to pMD19-T vector (Takara) and constructed pMD19T-534GG and pMD19T-534AA, digested pMD19T-534GG, pMD19T-534AA, and pGL3-basic with NheI and HindIII (Takara), to create pGL3-534-AA and pGL3-534-GG. The OCT1 CDS sequence of Hu sheep was amplified used P4 (Table 1) and first obtained pMD19T-OCT1, and then cloned into the pCDNA3.1 (+) vector (Invitrogen) with NheI and HindIII (Takara) to obtain an overexpression vector pCDNA3.1-OCT1. All the constructs were confirmed by dual-enzyme digestion and sequencing. The plasmids were extracted using the Endo-Free Plasmid Mini Kit (Tiangen, China).
Cell culture
The COS-7 cell line (Tongpai Biology Company, Shanghai, China) was cultured in RPMI1640 (Gibco) supplemented with 10% fetal bovine serum (FBS) (Gibco-Invitrogen) in a humidified air atmosphere with 5% CO2 at 37°C, and subcultured every 2–3 days in fresh media. The cells were seeded into 24-well plates for transfection.
GCs were separated from the sheep fresh ovaries obtained from the Dongling slaughterhouse (Taicang, China) following the methods described by Du et al. (2016), and cultured at 37°C and 5% CO2 in Dulbecco's modified Eagle's medium 12 (DMEM/F-12) (Gibco) containing 15% FBS (Gibco), 100 U/mL penicillin, and 100 mg/mL streptomycin (Gibco).
Transfection
Luciferase reporter plasmids and overexpressing OCT1 plasmids were transfected into COS-7 cell lines and sheep GCs using Lipofectamine 3000 (Invitrogen) according to the manufacturer's instructions. Briefly, COS-7 cells were seeded to be about 80% confluent at 24-well plates for transfection luciferase reporter plasmids and overexpression OCT1 plasmids to measure the luciferase activity, and GCs were seeded at 6-well plates for transfection overexpression OCT1 plasmids to detect cell apoptosis. Then the Lipofectamine 3000 and different plasmids were diluted by Opti-MEM medium, respectively. Five minutes later, diluted plasmids were added to each tube with diluted Lipofectamine 3000 reagent (1:1) and incubated for 25 min. Last, plasmids and Lipofectamine mixture were added to the cells and cultured for 48 h, and all the remaining steps were performed following the methods described by Du et al. (2016).
Luciferase assay
The cells were harvested and the lysates were collected 24 h post-transfection, and then, firefly and Renilla luciferase activities were measured using the Dual-Luciferase Reporter Assay System (Promega, China) according to the manufacturer's instruction. The relative luciferase activity indicates the ratio of firefly luciferase activity to Renilla luciferase activity as a control.
Apoptosis analysis
The apoptosis of sheep GCs was determined using the Annexin V-FITC/Propidium Iodide Apoptosis Kit (KeyGen, Nanjing, China), according to the manufacturer's protocol. The apoptosis rate of GCs was calculated and analyzed using FlowJo software (V7.6; Stanford University, Stanford, CA).
Statistical analyses
PopGene v1.31 software was used to calculate gene frequencies, genotype frequencies, heterozygosity, and polymorphism information content. Haploview 4.2 software (Barrett et al., 2005) was used to analyze the linkage relationship among three SNPs. The association analysis between different genotypes and the litter size of Hu sheep was referenced in Li et al. (2015). All data are expressed as mean ± standard error of the mean of at least three independent experiments. Statistics analyses were performed using SPSS16.0 software (SPSS, Inc., Chicago, IL). The results with p < 0.05 were considered statistically significant.
Results
Identification and characterization of the GDF9 promoter
A 1789 bp sequence of the GDF9 promoter from −1746 to +43 nt in Hu sheep was amplified and sequenced. Promoter prediction software showed that GDF9 gene of Hu sheep contained two possible promoter regions (score ˃0.90), located at −633 to −583 nt, and −268 to −220 nt, respectively (Fig. 1). In addition, using JASPAR online software, several binding sites for the transcription factors near the two predicted promoter regions, including CCAAT/enhancer-binding protein (C/EBP), Fos proto-oncogene (FOS), SRY-box transcription factor 3 (SOX3), signal transducer and activator of transcription 5 (STAT5), E74-like ETS transcription factor 5 (ELF5), pancreatic and duodenal homeobox 1 (PDX1), and nuclear receptor subfamily 4 group A member 2 (NR4A2), were predicted to be present in this region. MethPrimer software showed that there was a CpG island, located from −1699 to −1472 nt, including 16 CpG sites (Fig. 1).
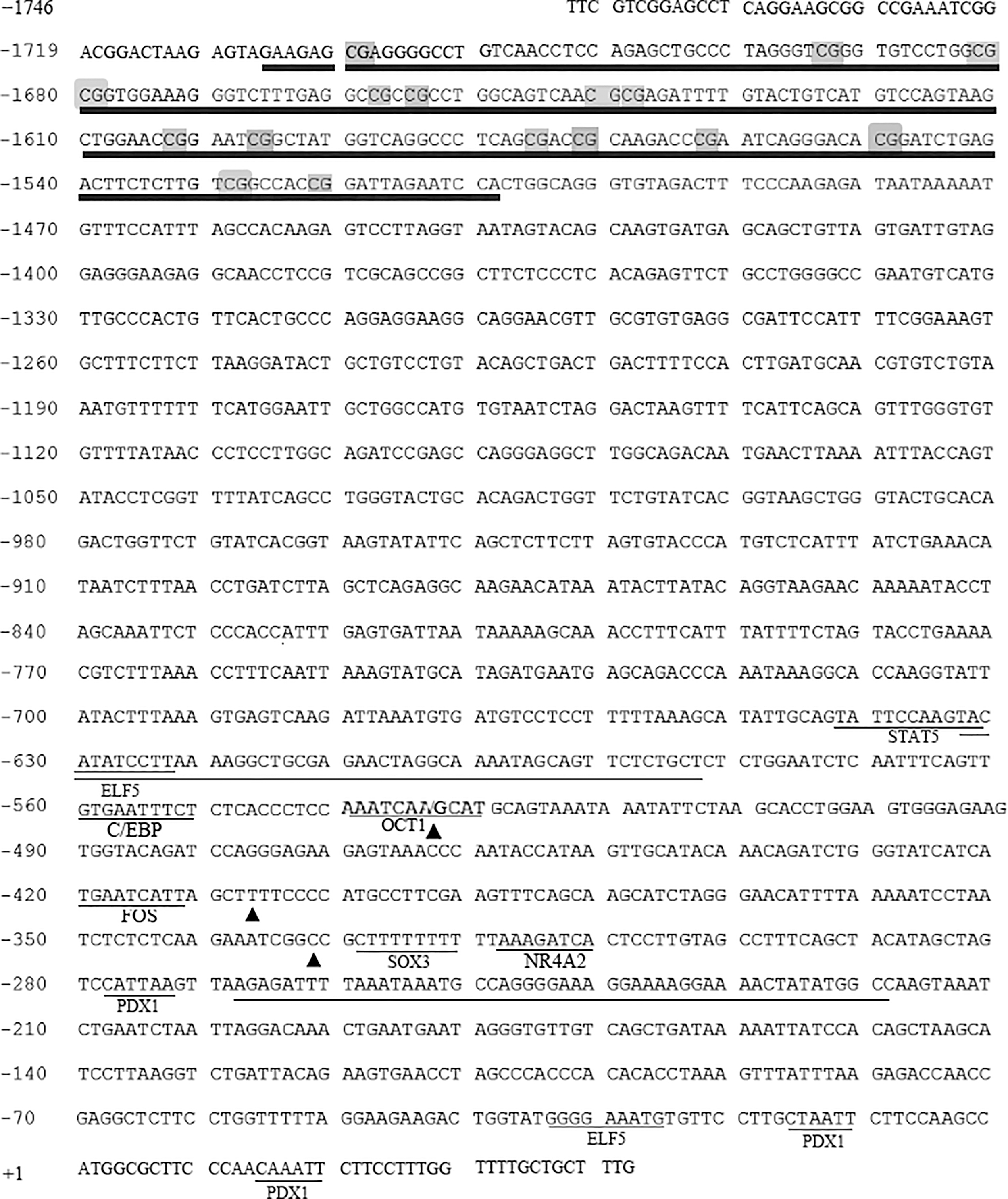
Characterization of the GDF9 promoter in Hu sheep. Long thin underline: predicated promoter region; Short thin underline: different transcription factor binding site; long thick underline predicated CpG island, the gray overlap represents the CpG site in the CpG island; the black triangles were SNP sites. GDF9, growth differentiation factor 9; SNP, single-nucleotide polymorphism.
The promoter activities of Hu sheep GDF9 gene were analyzed and are shown in Figure 2. The result showed that the activity of plasmid pGL3-1 (−309∼+43) was significantly higher than that of pGL3-basic (p < 0.05), while the activity of pGL3-2 (−725∼+43) significantly decreased compared with pGL3-1 (−309∼+43) (p < 0.05). The activity of plasmid pGL3-3 (−1239∼+43) was higher than pGL3-basic (p < 0.05) but no significant different with that of plasmid pGL3-1 (−309∼+43) and pGL3-2 (−725∼+43) (p > 0.05). All these data suggested that there was a negative regulatory element in the region within −725 to −309 bp and a positive regulatory element in the region within −309 to +43 bp.

Promoter activity identification of ovine GDF9 gene. *p < 0.05.
SNPs are associated with the litter size in Hu sheep
By DNA pool sequencing, three mutations were identified within −725 to −309 bp promoter regions of the GDF9 gene; they were −332C/T, −407 T/G, and −534 A/G, respectively (Fig. 3). Furthermore, the linkage analysis revealed that these three mutation loci were completely linked, both D’ and r2 had a value of 1.0 among these three loci (Fig. 4A). The genotyping results indicated three genotypes in the population of 267 ewes, that is, A-T-C/A-T-C, A-T-C/G-G-T, and G-G-T/G-G-T. We named A-T-C/A-T-C, G-G-T/G-G-T, and A-T-C/G-G-T as AA, BB, and AB genotypes, respectively. The frequency of AA, BB, and AB genotypes was 62.9%, 32.6%, and 4.5%, respectively (Table 2). The association between polymorphisms and litter size was investigated in Hu sheep populations, and the average litter size of the AA genotype (1.69 ± 0.06) was more than that of BB genotype (1.33 ± 0.14) (p < 0.05) (Fig. 4B). This indicated that these three linkage mutations in the promoter region of the GDF9 gene were associated with the litter size in Hu sheep.

SNP screening of ovine GDF9 promoter.

Linkage analysis of three SNPs in the GDF9 promoter region and the association analysis between the genotype and litter size in Hu sheep.
Genetic Diversity Index of Single-Nucleotide Polymorphisms in Sheep Growth Differentiation Factor 9 Promoter
He, heterozygosity; PIC, polymorphism information content.
To investigate whether the three mutations in the promoter region affected the promoter activity of the GDF9 gene, the AA- and BB-type promoter regions were cloned into the pGL3-basic vector, resulting in pGL3-AA and pGL3-BB, which were transfected into COS-7 cells. The results indicated that the luciferase activity of pGL3-AA was higher than that of pGL3-BB, but there was no significant difference (p > 0.05) (Fig. 5A).
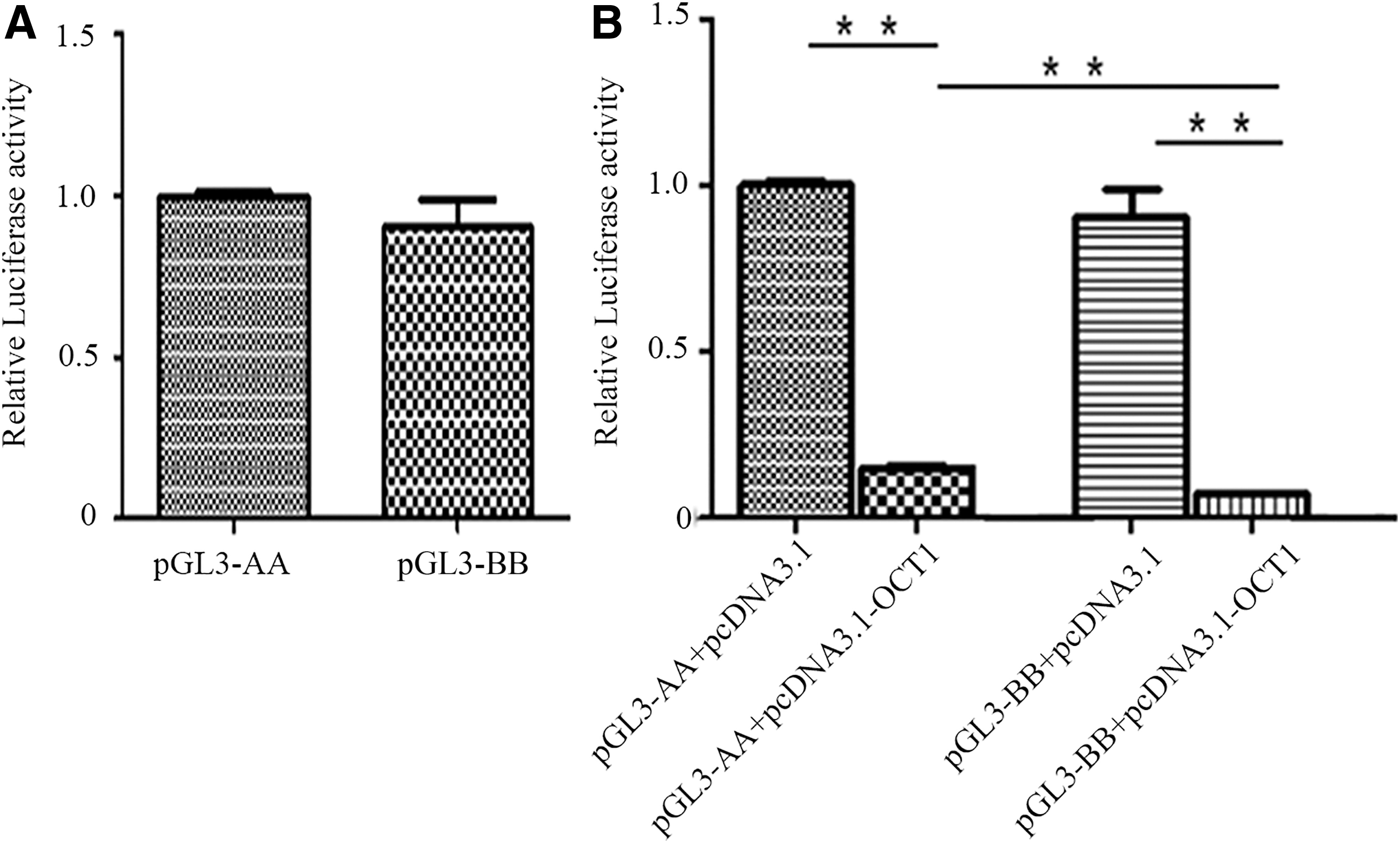
Effect of the transcription factor OCT1 on the promoter activity of different genotypes of the GDF9 gene in sheep **p < 0.01.
OCT1 transcription factor regulates the activity of the GDF9 promoter
The −534A/G mutation in the promoter region resulted in a novel transcription factor binding site OCT1. To determine whether the −534A/G mutation affected OCT1 regulation of GDF9 transcription, an OCT1 overexpression vector pCDNA3.1-OCT1 was constructed. pcDNA3.1-OCT1, pGL3-AA, and pGL3-BB were subsequently transfected into COS-7 cells. The promoter activity assay indicated that the overexpression of OCT1 significantly inhibited the luciferase activity of both genotypes (−534AA and −534 GG genotypes). Furthermore, the inhibition effect of pGL3-BB was greater than that of pGL3-AA (p < 0.05) (Fig. 5B).
OCT1 induces apoptosis in ovarian GCs
To further analyze the function of OCT1 in GCs, pcDNA3.1-OCT1 was transfected into sheep GCs, and the Annexin V FITC/PI flow cytometry assay showed that the apoptosis rate of GC overexpression of OCT1 was extremely higher than that of the control (Fig. 6), which indicated that the OCT1 gene could induce apoptosis in ovarian GCs.
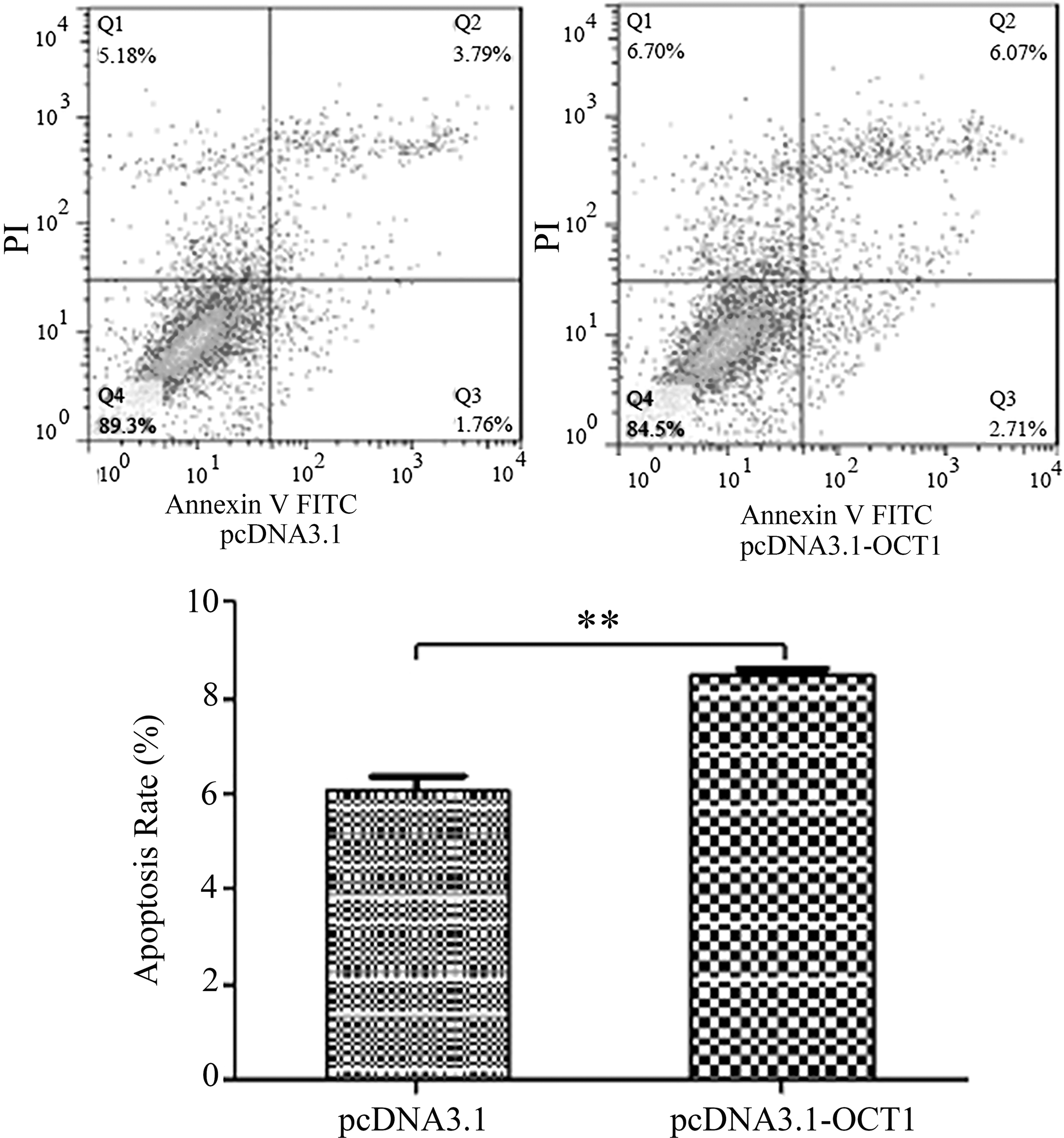
Effect of the transcription factor OCT1 on the apoptosis of GC **p < 0.01. GC, granulosa cell.
Discussion
It is well know that the promoter activity could affect the corresponding gene mRNA expression and further regulate biological functions such as growth (Li et al., 2017; An et al., 2018), reproduction (Li et al., 2018, 2019), and heat stress (Salces-Ortiz et al., 2015; Badri et al., 2018). The GDF9 gene is an important member of the TGF-β superfamily, and is involved in regulating various biological functions especially ovarian function and reproduction in mammals (Belli and Shimasaki, 2018; Wang et al., 2019). Studies have shown that the GDF9 gene can be used as a candidate molecular marker for breeding high-prolific sheep (Bravo et al., 2016; Abdoli et al., 2018; Wang et al., 2018). Little related research on the promoter and SNPs of the GDF9 gene, and its correlation with the litter size in sheep has been reported. In the present study, a 1789 bp promoter sequence of the GDF9 gene in Hu sheep was sequenced, and two possible promoter regions and one CpG island were predicted. We found several transcription factor binding sites near the predicted promoter regions such as C/EBP, FOS, SOX3, STAT5, ELF5, PDX1, and NR4A2, which are important factors regulating reproduction. For example, SOX3 has been reported to bind to the 11β-hsd gene promoter and transactivate to regulate male reproduction in teleost (Rajakumar and Senthilkumaran, 2016). FOS could block transcriptional activity of the nuclear receptor steroidogenic factor 1 and further regulate steroid production (Sirianni et al., 2010). NR4A2 has been reported to be involved in the regulation of the luteal function in rats (Qi et al., 2018). Therefore, it is reasonable to believe that these transcription factors might also play an important role in regulating the function of the GDF9 promoter.
In the GDF9 gene promoter, three completely linked polymorphisms (−332C/T, −407 T/G, and −534 A/G) were identified and were associated with the average litter size in Hu sheep, and the individuals with the AA (A-T-C) genotype had a larger litter size than those with the BB (G-G-T) genotype (p < 0.05). Further analysis showed that the presence of the −534GG allele resulted in an OCT1 binding site. OCT1, also named as Pou2f1, is a ubiquitously expressed member of the family of octamer-binding proteins containing the POU-homeodomain (Sturm et al., 1988; Veenstra et al., 1997). As a transcription factor, OCT1 could bind to the asymmetrical octamer canonical sequence ATGCAAAT and variants of this motif. This has been shown to drive the expression of both ubiquitous and tissue-specific genes (Scholer, 1991), and further regulate physiological and pathological processes such as development, growth, reproduction, and stress response (Sebastiano et al., 2010; Puscheck et al., 2015). It has been demonstrated using the ChIP and sequential chromatin immunoprecipitation assays that OCT1 is recruited to the FASL (Fas ligand) promoter region due to the rs763110 C/T mutation downregulating the FASL gene transcription (Wu et al., 2017). OCT-1 could bind within DHS3 to silence follicle stimulating hormone receptor (FSHR) transcription in myoid cells (Hermann and Heckert, 2005). Overexpression of either Oct-1 or Oct-2 in a fibroblast cell line markedly inhibited the HIV-1 long terminal repeat promoter transcriptional activity (Liu and Latchman, 1997; Cron, 2001). In this study, we found that the −534A/G mutation in the GDF9 promoter resulted in an OCT1 binding site. Furthermore, the luciferase activity assay showed that the overexpression of OCT1 decreased the activity of −534 GG genotype of GDF9, suggesting that OCT1 is involved in downregulating the promoter activity by binding with the −534GG genotype promoter of the GDF9 gene and further affecting the litter size in Hu sheep. We noticed that the overexpression of OCT1 not only significantly inhibited the −534GG genotype promoter activity but also significantly decreased the −534AA genotype promoter activity significantly; the activity of the −534GG genotype promoter decreased more than that of the −534AA genotype promoter, which is probably the reason of decrease of luciferase activity within −725 to −309 of GDF9 promoter. In human granulosa-luteal cells, OCT1 consensus sequence inhibited protein complex formation with the C/EBP motif in the gonadotropin releasing hormone (GnRH) receptor promoter, suggesting a possible cross talk between the OCT-1 transcription factor and this C/EBP motif (Cheng et al., 2002), and OCT1 could inhibit the FASL promoter activity by interacting with C/EBP (Wu et al., 2017). In the GDF9 promoter region, there was a C/EBP transcription factor binding site near the −534A/G locus. Therefore, we speculated that OCT1 probably first formed a complex with C/EBP and then downregulated the promoter activity of GDF9; new OCT1 binding site formed by the −534A/G mutation further enhanced downregulation of the GDF9 promoter activity.
Furthermore, researchers have found that the transcription factor OCT-1 might participate in cellular response to DNA damage (Zhao et al., 2000; Jin et al., 2001), and that OCT1 could promote liver cancer cell proliferation via calpain 6 (CAPN6) (Liu et al., 2016). In our study, we also found that the overexpression of OCT1 in GCs induced apoptosis. It is well known that there are complex and necessary bidirectional communications between GCs and oocytes during ovarian development and fertility in animals. GCs contribute to oocyte antioxidant defense against reactive oxygen species, and provide nutrients and molecular signals that regulate oocyte development, while oocytes promote the organization of follicles, and proliferation, differentiation, and function of GCs (Eppig, 1991; Soyal et al., 2000; Matzuk et al., 2002; Gilchrist et al., 2004; Devine et al., 2012; Belli and Shimasaki, 2018). GDF9 is an important oocyte-derived factor, and it has been implicated to be essential for the development of follicles and proliferation/apoptosis of GCs (Tuck et al., 2015; Li et al., 2019). In the present study, on one hand, OCT1 downregulated the promoter activity of the GDF9 gene, which might further affect oocyte function; on the other hand, OCT1 promoted GC apoptosis, and then, the two aspects might together negatively regulate ovarian function, even the litter size in Hu sheep (Fig. 6).
In summary, there are three complete linkage SNPs in the GDF9 promoter region and they affect the litter size in Hu sheep. Furthermore, GDF9 mutations predicated to result in a novel OCT1 transcription factor binding site, which can downregulate the GDF9 promoter activity by directly binding with the −534 GG allele and probably forming a complex with C/EBP (Fig. 7). Our findings highlight the bidirectional mutual regulation between oocytes and GCs and contribute to the understanding of the molecular mechanism of GDF9-regulated ovarian function and reproduction in female mammals.

Schematic representation of the proposed regulating pathway of OCT1–GDF9 between oocyte and GCs in Hu sheep.
Footnotes
Disclosure Statement
No competing financial interests exist.
Funding Information
This study was supported by the Natural Science Foundation of China (grant No. 31501934) and the Jiangsu Province Independent Innovation Fund (grant No. CX(18)3004).