
Abstract
microRNAs (miRNAs) have a crucial role in erythropoiesis. However, the understanding of the apoptosis of erythroid lineage remains poorly understood. Hence, an additional examination is required. K562 cell lines can be differentiated into early erythrocytes by hemin and the model of early erythrocytes can be established, consequently. miR-196a has been proven to take part in antiapoptosis in many cell lines. However, the role of miR-196a associated with the apoptosis in hemin-induced K562 cells remains unclear. To study the potential function of miR-196a involved in the common progenitor of erythroblasts, miR-196a mimics and microRNA-small hairpin negative control (miRNA-ShNC) were transfected into hemin-induced K562 cells with lentiviruses. After that, the viability of the transfected hemin-induced K562 cells was tested by CCK-8 assay, and the alteration of cell cycle and apoptosis rate were detected by flow cytometry. Furthermore, bioinformatics and dual-luciferase report system verified that p27kip1 is a target gene of miR-196a. Additionally, the expression of some proteins associated with cell cycle and apoptosis was tested by Western blotting assays. It was found that after overexpressing miR-196a, the proliferation of hemin-induced K562 cells was promoted while the apoptosis inhibited. Furthermore, miR-196a combines with the 3′UTR of p27kip1 directly. Additionally, the relationship between miR-196a and the protein level of p27kip1 is negative. After restoring the expression of p27kip1, the growth rate of hemin-induced K562 cells was not as high as before and the inhibition of apoptosis was alleviated. The present study validates that miR-196a overexpression inhibits apoptosis in hemin-induced K562 cells through downregulating p27kip1.
Introduction
K562 cells are derived from human erythroid leukemic cell line, which can differentiate into erythroid lineage under the induction of hemin, while into the megakaryocytic lineage under 12-O-tetradecanoyl phorbol acetate (TPA) (Chou and Hsu, 2009). After hemin treatment, benzidine staining, the expression of erythroid-specific gene γ-globin, and CD235a would be used to distinguish the differentiated K562 cells (Ma et al., 2013; Wang et al., 2018; Li et al., 2018b). Hemin is a molecule with many properties; in addition to its cellular toxic effects, it may also have cellular protective effects. For example, the cell growth rate of K562 cells was declined to some extent after the treatment of hemin (Huo et al., 2006).
microRNAs (miRNAs) are a kind of endogenous, small (about 16–24 nucleotides long), and noncoding RNA molecules. miRNAs negatively modulate their target genes either through targeting messenger RNAs (mRNAs) for degradation, or through inhibiting translation. Actually, miRNAs participate in various cell biological regulatory processes, such as proliferation, cell cycle, and apoptosis (Vienberg et al., 2017). To date, some miRNAs, such as miR-223, miR-210, miR-451, and miR-144, are considered to play a significant role in regulating the erythropoiesis (Sarakul et al., 2013; Gentner et al., 2015; Kim et al., 2015; Liang et al., 2018). However, studies on the apoptosis of early erythroid cells remain limited. miR-196a is derived from the miR-196 family (Slater et al., 2014). There are some researches about the involvement of miR-196a in other cell lines. Yang et al. (2017) reported that the downregulation of miR-196a can inhibit the proliferation and migration of human hepatoma cells through targeting Foxo1. Furthermore, Zhang et al. (2019) showed that miR-196a induces and enhances the proliferation of immature porcine sertoli cells while reduces the number of the apoptotic cells. These findings indicate miR-196a might also influence the proliferation and apoptosis of hemin-induced K562 cells, which is the model of early erythrocytes, and that the mechanism is well worth exploring.
Cyclin-dependent kinase inhibitor (CDKI) p27kip1 is a factor which can modulate cell cycle through some molecular mechanisms (Gonçalves, 2018). High levels of p27kip1 suppress the process of the cell cycle by inhibiting cyclin-dependent kinases (CDKs). Furthermore, p27kip1 also participates in progression of cellular growth, proliferation, differentiation, and apoptosis (Abbastabar et al., 2018). In addition, p27kip1 is a target of miR-196a according to bioinformatic prediction, and includes a complementary binding region with miR-196a.
This study investigates the function of miR-196a about the proliferation, cell cycle, and apoptosis of hemin-induced K562 cells, which is a model of early erythroid cells. Furthermore, the investigators aimed to confirm if p27kip1 is a target of miR-196a directly. Consequently, it was discussed whether miR-196a might be a model target for further studies on the therapy of anemia.
Materials and Methods
Cell culture
The K562 leukemia cells were acquired from the American Type Culture Collection (ATCC, CCL-243). They were cultured in RPMI 1640 media (Gibco), added with 10% FBS (Gibco), 2 mM of glutamine, 50 U/mL of penicillin/streptomycin, in an incubator with 5% CO2 concentration at 37°C. K562 cells were differentiated into erythroid lineage under the induction of hemin (25 μM; Sigma). HEK 293 cells (ATCC, CRL-1573) were cultured in DMEM media (Gibco) with 10% FBS (Gibco) in an incubator with 5% CO2 concentration at 37°C.
Cells transfection
The lentiviral vector with the miR-196a mimics, miRNA-small hairpin negative control (miRNA-ShNC) and psiCHECK™2-p27kip1 were purchased from Hanbio Biotechnology (Shanghai, China), and the sequences are showed in Table 1. Viruses were used to infect the hemin-induced K562 cells at a multiplicity of infection of 30 with 5 μg/mL of polybrene (Hanbio, Shanghai, China). Further studies were performed after infection for 48 h.
Sequences of the miR-196a Mimics
Cell proliferation assay
The proliferation was tested by using CCK-8 assay (Dojindo, Japan). First, cells were plated in 96-well plates. Then, after infection with miR-196a mimics or miRNA-ShNC, cells were added with 10 μL of CCK-8 solution to each well at 0, 24, 48, and 72 h. Subsequently, the plates were incubated for 2 h. Cell proliferation was measured by scanning at 450 nm with a microplate reader (Powerwave XS2).
Cell cycle and apoptosis assay
Cells were collected after miR-196a mimics or miRNA-ShNC infection for 48 h. For the cell cycle assay, the results were detected with Propidium Iodide (PI; Beyotime, Shanghai, China) staining and a flow cytometer (Beckman Coulter). For the analysis of cell apoptosis, cells were stained with Annexin V/fluorescein isothiocyanate/PI (Bestbio, Shanghai, China) and a flow cytometer.
Real-time PCR
TRIzol reagent (Invitrogen) was used to extract total RNA from cells. Next, the PrimeScript™ RT Reagent Kit (TaKaRa, Dalian, China) was accordingly used to synthesize the cDNA by reverse transcription. Real-time PCR was performed on an ABI 7500 (Applied Biosystems, Foster Coey, CA) thermal cycle with the SYBR Green qPCR Kit (TaKaRa, Dalian, China). U6 was used as the reference gene for miRNAs and GAPDH for mRNAs (the primers showed in Table 2). The ΔΔCt method for the relative quantification of gene expression was used to determine the levels of miRNA and mRNA.
Sequences of Primers for the RT-PCR
Benzidine staining
Benzidine staining is a qualitative approach, which can measure the hemoglobin accumulation of K562 cells. After hemin treatment, cells were washed with PBS three times. Then, 0.5 mL of suspended cells supplemented with 14 μL of 0.4% benzidine solution (dissolved with 1 mL of 12% acetic acid) was incubated for 3 min. Afterward, 1 μL of 30% hydrogen peroxide was added. After 5 min, 1 μL of 5% sodium nitroferricyanide dihydrate was added into the cell suspension for 3 min at room temperature. Benzidine-positive cells were stained blue, whereas negative cells were stained pale yellow. Around 300 cells were counted for each sample, and the proportion of benzidine-positive cells was calculated.
Western blot analysis
Total proteins from cells were extracted with a lysis buffer, which was made up with RIPA buffer (Thermo Fisher) and PhosStop cocktail (Roche, Mannheim, Germany). The concentration of protein was determined with a BCA Protein Assay Kit (Thermo Fisher), and the protein concentration of each sample was adjusted to be consistent. After that, the protein extracts were separated by SDS–PAGE with 40 μL for each sample, and transferred to nitrocellulose membranes in transfer buffer. Five percent nonfat dry milk was then used to block NC membranes for 2 h. After incubation with primary antibodies (p27kip1, CDK4, cyclin D2 [CCND2], Bax, Bcl-2, and actin, Cell Signaling Technology), as well as HRP-conjugated secondary antibodies (Cell Signaling Technology), the protein bonds were visualized by enhanced chemiluminescence (ECL; Thermo Fisher).
Bioinformatics analysis
The possible target genes of miR-196a were projected through miRBase (
Dual-luciferase reporter assay
The dual-luciferase reporter vectors psiCHECK™2-p27kip1 mutant type (mut)/wild-type (wt)/NC (400 ng/μL) were synthesized by Shanghai Hanbio Biotechnology (Shanghai, China). HEK293 cells were cotransfected with plasmids, and miR-196a mimics or miRNA-ShNC. The luciferase was measured at 48 h after transfection with the Dual-Luciferase Reporter Kit (Promega), in accordance with the manufacturer's instructions.
Statistical analysis
Whole experiments were conducted for at least three times, and whole data were expressed as mean ± standard deviation. The results were processed with the SPSS 16.0. The significant differences between groups were confirmed by ANOVA. The differences among groups were considered to be statistically significant at p < 0.05, p < 0.01, and p < 0.001.
Results
Characterization of γ-globin and CD235a expression and benzidine-positive cell rate over erythroid differentiation
Hemin can induce K562 cells to differentiate into the erythroid lineage. γ-globin and CD235a can be used as erythrocytic lineage markers to evaluate the differentiation. Twenty-four hours after treatment with 25 μM of hemin, the cells revealed features of erythrocyte differentiation, as measured by increasing the levels of γ-globin, CD235a and the rate of benzidine-positive cells. Two groups were performed. Cells in the first group were cultured for 24 h with 25 μM of hemin. After 24 h, these cells were resuspended with fresh medium, and collected at the following time points: 0, 24, 48, and 72 h. Cells in the second group were cultured with 25 μM of hemin for 24–72 h, and collected at the appropriate time. In addition, the 0-h cell groups serve as blank controls. Next, the mRNA levels of γ-globin and CD235a in the two groups were determined with qPCR. After 0 h, both levels of γ-globin and CD235a in hemin-induced K562 cells increased gradually, and the level declined after 24 h of treatment (Fig. 1A, B). For the rate of benzidine staining, after cells were collected at the above time points, PBS was used to wash them for three times. Then 0.4% benzidine solution, 30% hydrogen peroxide, and 5% sodium nitroferricyanide dihydrate were added for the reasonable time. In the end, a microscope was used to observe and count cells. In addition, similar results were also reflected in this experiment (Fig. 1C). These results show that K562 cells can efficiently differentiate into the erythrocytic lineage under the induction of 25 μM of hemin for 24 h. In addition, the inductive effect did not decrease when hemin was removed after the induction treatment for 24 h.

The 25 μM of hemin for 24 h can efficiently induce K562 cells to differentiate into the erythroid lineage. In the first group, cells were cultured for 24 h with 25 μM of hemin. After 24 h, cells were resuspended with fresh medium, and collected at appropriate time points. In the second group, cells were cultured with the same amount of hemin without resuspended with fresh medium, and collected at the appropriate time as well. Zero-hour cell groups serve as blank controls. Furthermore, the inductive effect will not decrease if hemin is removed.
The overexpression of miR-196a promotes hemin-induced K562 cell proliferation
To discuss the effect of miR-196a in the proliferation of hemin-induced K562 cells, miR-196a mimics or miRNA-ShNC were tranfected (Fig. 2A). The results revealed that after transfecting miR-196a mimics, the relative expression of miR-196a became significantly and obviously upregulated, when compared with the control. The results of the CCK-8 analysis indicated that the miR-196a mimics prominently enhanced the viability of hemin-induced K562 cells, when compared with control cells (the miRNA-ShNC group and the blank control group) (Fig. 2B). The cell cycle analysis revealed that in comparison with control groups, the percentage of miR-196a mimics-transfected hemin-induced K562 cells during the G0/G1 phase was less, while more cells were detected in the S and G2/M phase significantly, which means in the group of miR-196a mimics, more cells transited from the G0/G1 phase into S phase than control groups (Fig. 2C). The results show that miR-196a promotes the proliferation and accelerates the transition from the G0/G1 phase into the S phase in hemin-induced K562 cells.
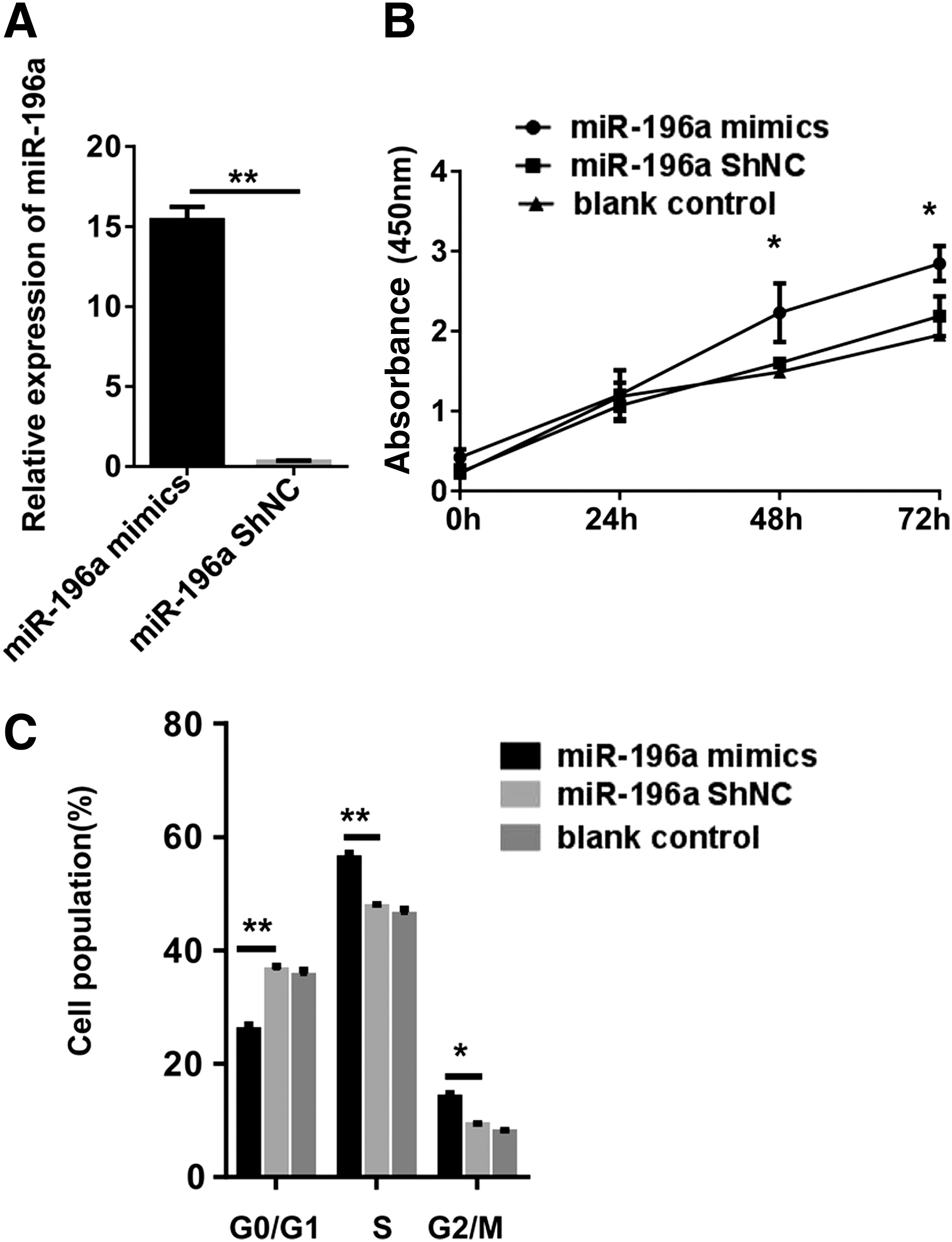
miR-196a upregulation promotes cell proliferation and cell cycle transition in hemin-induced K562 cells.
The overexpression of miR-196a inhibits cell apoptosis in hemin-induced K562 cells
To determine whether antiapoptosis took part in the miR-196a mimic-transfected hemin-induced K562 cell proliferation, cells were labeled with Annexin-V/PI double staining and analyzed with a flow cytometry to detect the apoptosis rates. Hemin-induced K562 cells were transfected with the miR-196a mimics or miRNA-ShNC. The cell apoptosis assay results revealed that the number of apoptotic hemin-induced K562 cells was prominently lower in the miR-196a mimics group, compared with that in control groups (Fig. 3A, B). To study the molecular mechanism of antiapoptosis, some apoptotic proteins, such as Bax and Bcl-2, were also detected (He et al., 2017). The Western blot results revealed that the overexpression of miR-196a downregulated the expression of Bax, but upregulated the expression of Bcl-2 (Fig. 3C). To sum up, these results suggest that miR-196a might inhibit apoptosis in hemin-induced K562 cells.
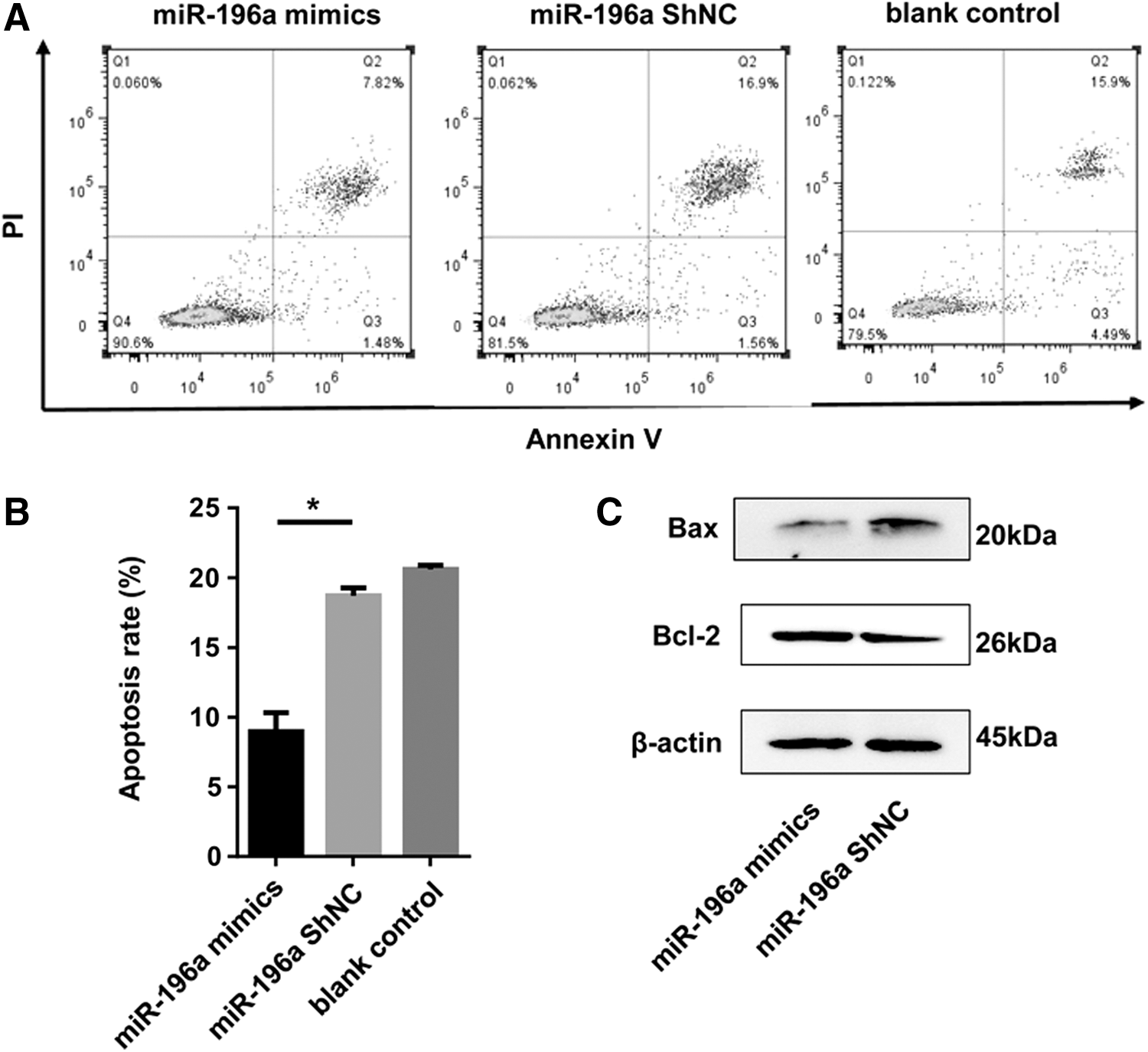
miR-196a overexpression inhibits cell apoptosis in hemin-induced K562 cells.
Prediction and identification of the miR-196a target gene in hemin-induced K562 cells
The present study further selected the target genes of miR-196a to explore the effect of miR-196a in hemin-induced K562 cells. The prediction of bioinformatic analysis showed that p27kip1 is a target gene of miR-196a (Fig. 4A). To confirm that, the seed sequence in the 3′UTR of p27kip1 was mutated (Fig. 4A). To validate this prediction, fragments of the 3′UTR of p27kip1 that contain the putative miR-196a-binding sites were inserted into the dual-luciferase reporter vector psiCHECK™2. The luciferase activity in hemin-induced K562 cells cotransfected with miR-196a mimics and the p27kip1-wt vector was markedly lower than that in hemin-induced K562 cells transfected with the p27kip1-mut/NC vector (Fig. 4B). These results show that p27kip1 is a direct target gene of miR-196a.

miR-196a targets the 3′UTR of p27kip1 in hemin-induced K562 cells.
miR-196a level negatively regulates the expression of p27kip1
It was further discovered that the miR-196a mimics could cause a decrease in p27kip1 expression at the protein level (Fig. 5). Apart from that, some important apoptotic or cell cycle control factors were changed as well on account of the overexpression of miR-196a (Figs. 3C and 5). The protein level of Bax decreased due to the overexpression of miR-196a relative to the control, whereas the level of Bcl-2, CCND2, and CDK4 increased. As is known that Bcl-2 is an antiapoptotic molecule, whereas Bax plays the role of activating apoptosis (Gharesi et al., 2017). CCND2 has been confirmed to cause cell cycle arrest at G1 (Chen et al., 2018; Zhang et al., 2018), and CDK4 have an inverse effect (Gong et al., 2017). This observation was consistent with recent results that miR-196a has a crucial effect on promoting G1/S transition, and inhibiting apoptosis in hemin-induced K562 cells. Thus, cell proliferation was promoted.
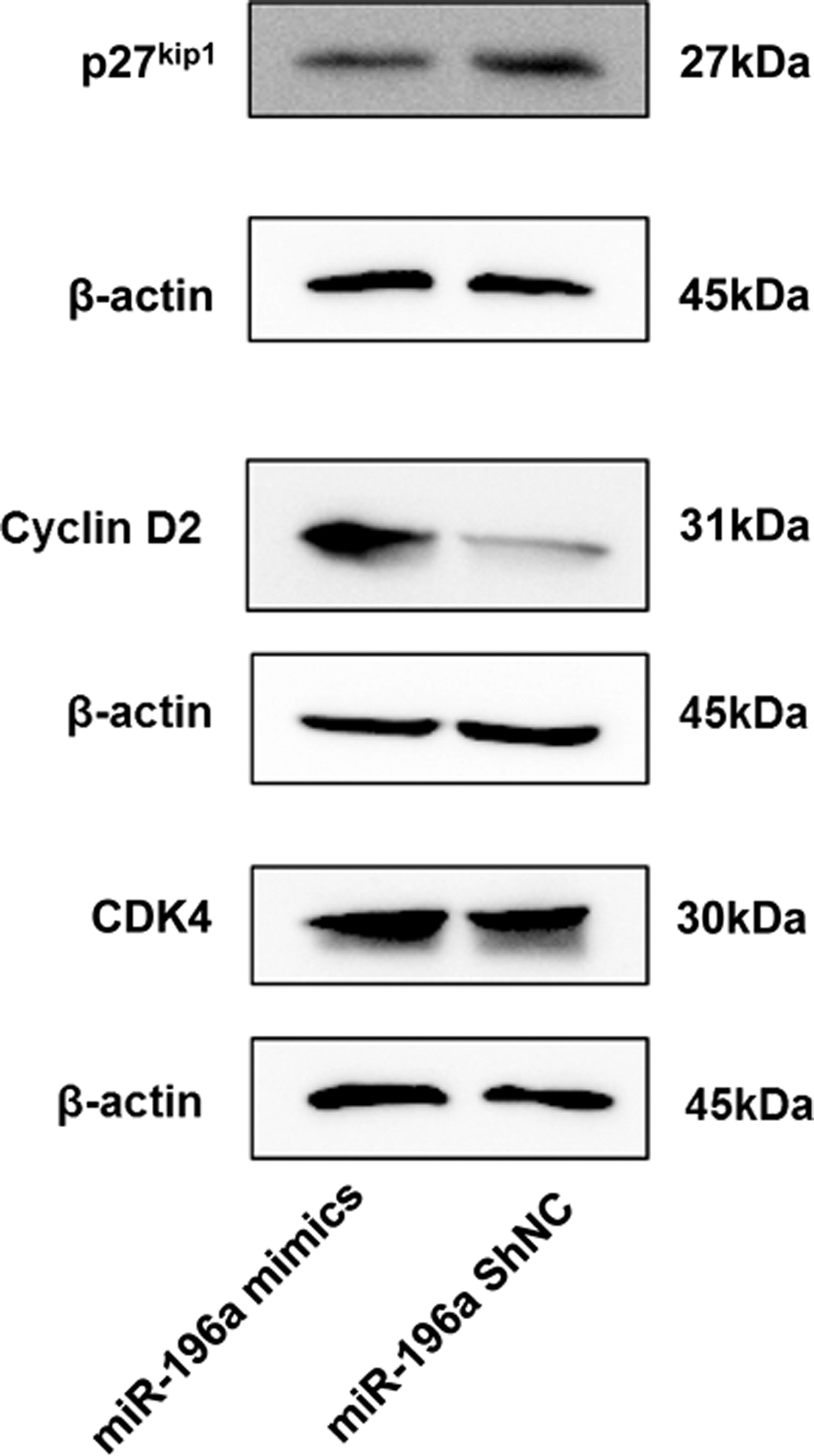
The protein level of p27kip1 is inversely correlated with miR-196a, whereas the protein level of CCND2 and CDK4 are directly correlated with miR-196a. These protein expressions were detected by Western blot in hemin-induced K562 cells transfected with miR-196a mimics or miRNA-ShNC, with β-actin as an internal control. Due to the approximate molecular weight of the three target proteins, it is difficult to incubate the primary antibodies on the same membrane, so the three proteins were separated on different gels with their own β-actin, respectively. All data are presented as mean ± SD (three independent replicates per group). CCND2, cyclin D2; CDK4, cyclin-dependent kinase 4.
miR-196a promotes cell proliferation and inhibits apoptosis by downregulating p27kip1
To confirm if miR-196a can facilitate cell proliferation and inhibit apoptosis through downregulating p27kip1 in hemin-induced K562 cells. Both psiCHECK™2-miR-196a mimics and psiCHECK™2-p27kip1 were transfected into the hemin-induced K562 cells, with the hemin-induced K562 cells cotransfected with psiCHECK™2-miR-196a mimics and psiCHECK™2-p27kip1-NC as a control group. After transfection, the protein level of p27kip1 increased (Fig. 6A). Next, the cell proliferation and apoptosis rate were measured again, with less cell growth rate and more cell apoptosis compared with the control (Fig. 6B–D). In summary, all these above results reveal that when miR-196a overexpressed in the hemin-induced K562 cells, the protein level of p27kip1 was downregulated, and the transition from the G0/G1 phase to the S phase would be accelerated. Hence, cell proliferation would promote, while apoptosis would decrease (Fig. 7).
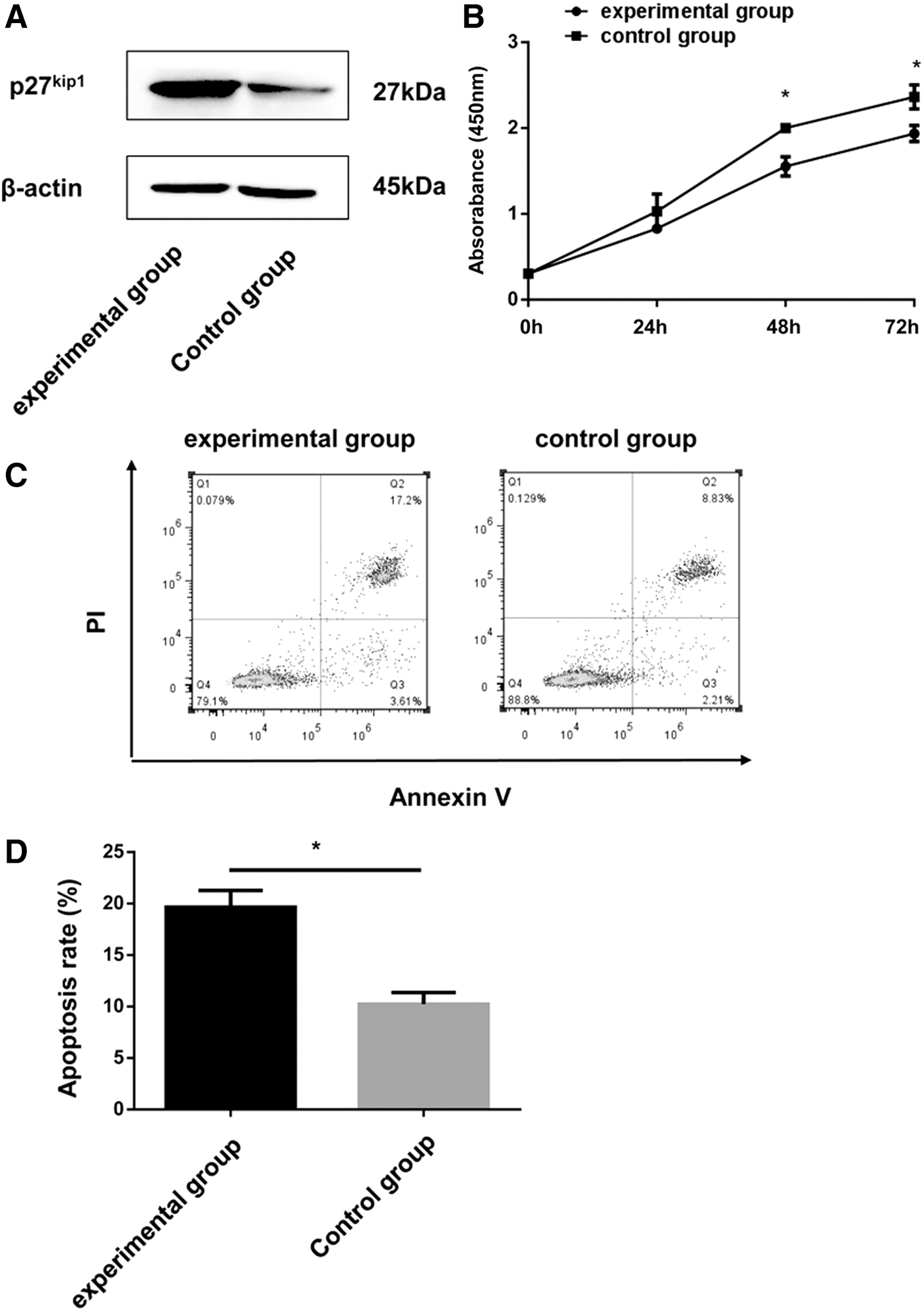
miR-196a promotes cell proliferation and inhibits apoptosis by downregulating p27kip1.
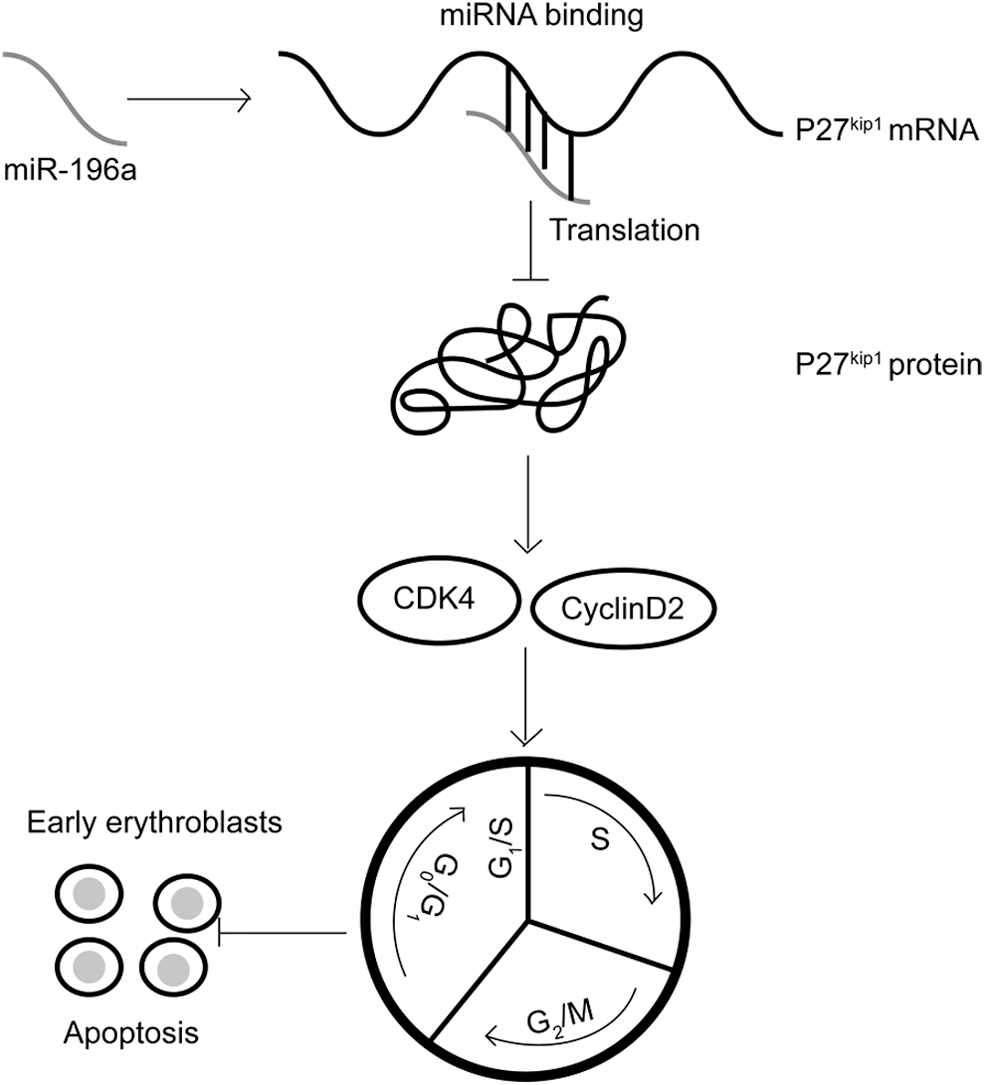
miR-196a negatively regulates the protein level of p27kip1 and its effects in hemin-induced K562 cells, the model of early erythroblasts. With the downregulation of the protein level of p27kip1 by miR-196a overexpression, the transition from the G0/G1 phase to the S phase would be accelerated. Then cell proliferation would be promoted and the apoptosis would be decreased.
Discussion
Emerging reports have demonstrated that miRNAs take part in the development and progression of erythroid lineage, especially for erythropoiesis (Kouhkan et al., 2014; Kim et al., 2015). Latest studies have indicated that when some storage lesions occur in mature erythrocytes during their storage, levels of some miRNAs like miR-150 and miR-196a have changed (Kannan and Atreya, 2010; Sarachana et al., 2015; Vu et al., 2017; Repsold and Joubert, 2018). Thus, the investigators make a hypothesis, in which miR-196a might regulate eryptosis (a special term for apoptosis of red blood cell) negatively. Owing to the special structure of mature erythrocytes, the purpose gene could not be transfected into them (Moras et al., 2017). Hence, K562 cells were used, which has been proven to be a model of early erythroblasts induced by 25 μM of hemin for 24 h, to transfect the lentiviral vector with the miR-196a mimics or miRNA-ShNC into them. Subsequently, an early erythrocyte apoptosis model with hemin-induced K562 cells was established.
Previous studies have mostly focused on the effect of miR-196a in tumorigenesis, including breast, gastric, pancreatic, colorectal, and cervical cancer (Li et al., 2015; Sarachana et al., 2015; Jiang et al., 2018; Zhong et al., 2018; Buscheck et al., 2019). However, in this study, the function of miR-196a, which promoted proliferation and inhibited apoptosis by binding the 3′UTR of p27kip1 in hemin-induced K562 cells, was discussed. That is, the function of miR-196a in early erythrocyte apoptosis was investigated.
It has been proven that CDKI p27kip1 is a key regulatory factor of the G1/S transition of the cell cycle. Moreover, the downregulation of p27kip1 leading to the acceleration of cell cycle progression (Satoh and Kaida, 2016; Gonçalves, 2018; Li et al., 2018a). This study bioinformatically predicted p27Kip1 as a possible target of miR-196a. The luciferase activity assay revealed that the decrease of the protein level of p27Kip1 was mediated by miR-196a through a direct interaction with 3′UTR of p27Kip1, respectively. Furthermore, upon miR-196a overexpression, fewer cells were arrested in the G0/G1 phase, whereas more cells remained in the S phase, which means that the overexpression of miR-196a could accelerate the transition from the G0/G1 phase to the S phase through the downregulation of p27kip1, and that the proliferation ability of hemin-induced K562 cells transfected with miR-196a mimics was enhanced, compared with the controls. Additional evidence suggests that other cell cycle-related proteins, such as CCND2 and CDK4, also participated in the regulation of cell cycle. Beyond the cell cycle, p27Kip1 can also regulate the apoptosis (Abbastabar et al., 2018). When miR-196a was forced, the apoptosis rate of hemin-induced K562 cells decreased, when compared with the control groups. Similar results were also reflected in the protein levels of Bax and Bcl-2. Then, the level of p27kip1 was restored in hemin-induced K562 cells, and subsequently the cell growth rate was descended while the apoptosis rate was ascended significantly. However, when miR-196a was inhibited, some results, such as cell proliferation and cell cycle transition, were not suppressed, and the apoptosis rate of hemin-induced K562 cells were not distinctly promoted. This could probably be because the inhibition of miR-196a was compensated by other molecular pathways.
A balance between cell proliferation and apoptosis should be maintained to avoid unnecessary tissue disorders (Clarke and Allan, 2009). When DNA damage occurred, the cell cycle can be blocked. Then, two results may appear, either completing the rest cell progress if the damage is repaired, or undergoing apoptosis if the damage is too severe to be repaired (Zhu et al., 2014). To sum up, cell proliferation and apoptosis are interrelated. Cell cycle regulators are involved not only in the regulation of cell proliferation, but also in the regulation of apoptosis (Bertrand et al., 2009). Therefore, this study conducted the following research by comprehensively analyzing the relationship between cell proliferation and apoptosis, and thus came to the conclusion.
In this study, the investigators aimed to discuss the mechanism of inhibiting apoptosis in early erythroblasts by upregulating miR-196a. Therefore, studies that reported miRNAs' participation in storage lesions, especially for the eryptosis of mature red blood cells, could be partly validated. However, the present study has some shortcomings. The cell model with other cell lines, such as CD34+ hematopoietic progenitor cells (Zhai et al., 2014; Zini et al., 2016; Liang et al., 2018) was not established, and a validation model of mature red blood cells could not be established due to its special membrane structure. Further studies on the function and structure of erythrocytes would help to solve this challenge (Talu et al., 2016; Himbert et al., 2017), and anemia might be improved through modulating miR-196a or other miRNAs.
Overall, this study shows that miR-196a has a crucial effect on the cell proliferation, cell cycle transition, and apoptosis of hemin-induced K562 cells, the model of early erythroblasts. In addition, miR-196a may regulate the biological progresses of hemin-induced K562 cells by targeting p27kip1. With the downregulation of the protein level of p27kip1, the transition from the G0/G1 phase to the S phase would be accelerated. Hence, cell proliferation would be promoted, while apoptosis would decrease. All of the results indicate that miR-196a may exhibit a therapeutic action on anemia.
Footnotes
Acknowledgments
The authors would like to thank Mr. J.S. for the financial support. The authors would also like to thank Ms. Y.X. and Mr. L.Y. for their technical support.
Disclosure Statement
No competing financial interests exist.
Funding Information
This research was funded by National Natural Science Foundation of China (grant nos. 81271865, 30771882).