
Abstract
Colorectal cancer (CRC) has been suggested to be one of the leading cancer types all over the world. Till now, the molecular mechanism by which circCCT3 regulates CRC remains to be clarified. To detect mRNA and protein levels of various genes, Reverse Transcription-quantitative PCR and western blot were used in our study. Luciferase reporter assay was utilized to probe direct interaction between genes. We used transwell assay to assess the invasion ability of CRC cells. For apoptosis detection, immunofluorescence of CRC cells by Annexin V staining was performed. We carried out bioinformatic analysis to show higher expression of circCCT3 in human clinical CRC tumors. Low level of circCCT3 was closely associated with higher disease-free survival of CRC patients. Moreover, we found that circCCT3 was linked to advanced stage of CRC. miR-613 is the target of circCCT3 and responsible for circCCT3-modulated invasion and apoptosis of CRC cells. In addition, we identified WNT3 and vascular endothelial growth factor A (VEGFA) as downstream effectors of miR-613 in CRC cells. WNT3 and VEGFA overexpression resulted in partial rescue of miR-613-mediated phenotypes of CRC cells. In conclusion, we propose that circCCT3 contributes to CRC metastasis via miR-613/WNT3 or miR-613/VEGFA, promoting the development of therapeutical approaches for treating CRC.
Introduction
Colorectal cancer (CRC) has been shown to be ranked among the top five leading causes of cancer-associated death, which is commonly caused by metastasis (Goldstein et al., 2016; Bray et al., 2018). According to previous reports, the distant metastatic site of CRC is the liver (Manfredi et al., 2006). About one-fourth of CRC patients are diagnosed with liver metastasis, and the 5-year survival rate is around 10% for metastatic patients (Chen et al., 2017). Based on these facts, it is crucial to illuminate the mechanism of CRC metastasis.
Circular RNAs (circRNAs) are defined as closed and single-stranded RNAs derived from back-spliced pre-mRNA (Jiang et al., 2018). Initially, circRNAs are discovered ∼20 years ago and believed as by-products of aberrant splicing (Nigro et al., 1991). The functions of circRNAs are rarely studied. CircCCT3 (circBase ID: hsa_circ_0004680) is originated from exons 3–5 and introns 3–4 of chaperonin containing TCP1 subunit 3 (CCT3) by back-splicing. circRNA has been suggested to be involved in various types of cancer (Kulcheski et al., 2016). CiRS7 promotes gastric and CRC by abrogating miR-7 (Hansen et al., 2013). Furthermore, circMTO1 sponges miR-9 to suppress hepatocellular carcinoma (Han et al., 2017).
microRNAs are 18–25 bp small noncoding RNAs (Bartel, 2004). They regulates cell proliferation, differentiation, apoptosis, and cancer metastasis through binding to 3′-UTR of related genes (Lin and Gregory, 2015). For instance, miR-133a inhibits MAPK signaling pathway and thereby modulating CRC progression (Wang et al., 2013). In mouse models of colon cancer, miR-29 loss enhances metastasis by increasing MMP2 expression (Schetter et al., 2012). It has been shown that miR-613 suppresses CRC metastasis via FMNL2 and RTKN (Li et al., 2016; Dong et al., 2019).
Vascular endothelial growth factor A (VEGFA) is one member of the VEGF family proteins (Cursiefen et al., 2004). It plays a critical role in cancer angiogenesis (Hirakawa et al., 2005). VEGFA is upregulated in breast cancer (Kurebayashi et al., 1999). George et al. (2001) indicated that VEGFA is implicated in CRC.
Wnt signaling is activated by WNT ligands (e.g., WNT3) binding to FRIZZLED and LRP receptor complexes, which is involved in diverse biological processes (Schatoff et al., 2017). Wnt3 downregulation significantly blocks gastric cancer cell proliferation (Wang et al., 2016). More importantly, some groups find that Wnt pathway contributes to CRC metastasis.
In this study, we aim to investigate the new mechanism underlying CRC progression. In addition, we also attempt to identify the downstream targets of circCCT3. Here, we show that circCCT3 regulates VEGFA and Wnt signaling to promote metastasis of CRC by sponging miR-613. The findings provide more insights into CRC progression and a potential biomarker or therapeutic strategy for CRC treatment.
Materials and Methods
Cell lines
Human CRC cells (HCT116 and SW480) and 293T cells were obtained from ATCC. These cells were all cultured in Dulbecco's modified Eagle's medium (DMEM) (Hyclone), which were added with 10% fetal bovine serum (FBS) and 1% penicillin/streptomycin.
Cell transfection
The WNT3 and VEGFA cDNAs were subcloned and ligated into pcDNA3.1 vectors. Then, we transfected recombinant vectors into cells using lipofectamine 2000 (Life Technologies). Besides, we used Lipofectamine RNAiMAX Reagent (Life Technologies) to introduce miR-613 mimic or miR-613 inhibitor into cells. miR-613 mimic or miR-613 inhibitor were synthesized by GenePharma.
Stable cell lines
Short hairpin RNAs (shRNAs) were annealed and inserted into pLKO.1-TRC vector. We made and harvested lentivirus through transfecting pLKO.1-shRNA, pVSVG, and pPAX2 into 293T cells. We collected the virus at 48 h. Upon treatment, we added the virus to cell medium at the ratio of 1:5. After additional 24 h, 2 μg/mL of puromycin were used to pick the resistant cells. sh-NC is the scramble vector. sh-NC: 5′-GTACCTGACTAGTCGCAGATT-3′; sh-circCCT3: 5′-TCTTGACTATTGCAGATATCATT-3′.
Luciferase reporter assay
Wild-type and mutant circCCT3, WNT3, and VEGFA were subcloned and ligated into pGL3-luciferase vector. Negative control (NC), miR-613 mimic, or miR-613 inhibitor was transfected with circCCT3, WNT3 3′-UTR, or VEGFA 3′-UTR-fused pGL3 vectors into 293T cells. Finally, the luciferase activities were determined using the Dual-Luciferase Reporter System (Promega).
Transwell assay
CRC cells (∼1 × 105) were resuspended with 200 μL of DMEM. The suspended cells were seeded into the top chamber containing Matrigel-coated membrane. About 400 μL media plus 20% FBS were added to the bottom chamber. After additional 48 h, the invaded cells were fixed and stained with 0.003% crystal violet. The pictures of stained CRC cells were taken, and the numbers were counted under the inverted microscope.
Immunofluorescence
We performed this experiment using CRC cells, as previously described (Schutters et al., 2013). In brief, the cells were fixed with 4% paraformaldehyde. Then, Triton X-100 was used to treat and permeabilize the CRC cells. Bovine serum albumin (BSA) was used to block the cells for 1 h at room temperature. We incubated cells with Annexin V-FITC antibody (BD Pharmingen). The photographs were captured under fluorescence microscope, and apoptosis rates were calculated as Annexin V-positive cells/total cells.
Reverse Transcription-quantitative PCR
The total RNAs were extracted and purified by treatment of TRIzol reagent (Invitrogen). First strand of cDNA was generated using reverse transcription reagent (TaKaRa, Japan). We used SYBR Green dye (Roche) in real-time polymerase chain reaction (PCR) experiment. GAPDH is used as internal controls for circCCT3, VEGFA, and WNT3, whereas U6 is used as internal controls for miR-613. The primers were used as follows: circCCT3-F: GGACCCAGGATGAAGAGGTT; circCCT3-R: CATTGGGTCCAAAAGCATCT; miR-613-F: GTGAGTGCGTTTCCAAGTGT; miR-613-R: GGGTCCCTTCACACTTGGAA; WNT3-F: CGTGTTAGTGTCCAGGGAGT; WNT3-R: CTACTACCATCTCCGAGGCG; VEGFA-F: GGCCAGCACATAGGAGAGAT; VEGFA-R: ACGCTCCAGGACTTATACCG; GAPDH-F: GAGAAGTATGACAACAGCCTC; GAPDH-R: ATGGACTGTGGTCATGAGTC; U6-F: CTCGCTTCGGCAGCACATATACTA; U6-R: ACGAATTTGCGTGTCATCCTTGCG
Western blotting
We resolved ∼50 μg of proteins onto SDS-PAGE (sodium dodecyl sulfate–polyacrylamide gel electrophoresis) gel, and then, the proteins were transferred to polyvinylidene fluoride membrane. After that, the membrane was incubated with corresponding primary antibodies at 4°C overnight. Secondary antibodies in BSA were incubated with the membrane. In this experiment, the following antibodies were used: anti-Wnt3 (Abcam), anti-VEGFA (Abcam), and anti-GAPDH (Cell Signaling Technology).
MS2-RNA immunoprecipitation assay
HCT116 cells containing pMS2-GFP were co-transfected with pcDNA3.1-MS2, pcDNA3.1-MS2-circCCT3-WT, or pcDNA3.1-MS2-circCCT3-MUT. After 48 h, RNA immunoprecipitation (RIP) experiment was carried out using GFP antibody and Magna RNA-Binding Protein Immunoprecipitation Kit (Millipore). The magnetic bead–antibody complexes and cell lysate were incubated overnight. After reverse transcription, Reverse Transcription-quantitative PCR (RT-qPCR) of miR-613 was performed to confirm miR-613 binding by circCCT3.
The Cancer Genome Atlas data sets
Clinical data of CRC patients were obtained from The Cancer Genome Atlas (TCGA). The RNA levels were measured by RNAseq with Illumina HiSeq. There were 349 normal subjects and 275 CRC patients analyzed in this study. The software Enguage Digitizer (Version 4.1) was used to extract the disease-free survival of CRC patients from TCGA. We performed box plot and survival curve by GraphPad Prism 6.0 Software.
Statistical analysis
In all experiments, three biological replicates were performed to calculate as mean ± standard deviation. Comparison of two experimental groups was performed by the two-tailed unpaired Student's t-test; one variance comparison of multiple groups was carried out by analysis of variance (ANOVA). *p < 0.05 was considered as statistically significant data.
Results
circCCT3 is upregulated in clinical CRC tissues
To explore whether circCCT3 had a role in CRC metastasis and progression, we reanalyzed TCGA data sets of CRC and examined the expression level of circCCT3 according to the published method. We revealed that circCCT3 levels in human CRC tumors were remarkably increased compared with normal tissues (Fig. 1A). Low level of circCCT3 in CRC patients correlated with higher disease-free survival rate (p = 0.016, n = 135) (Fig. 1B). Besides, we noted that circCCT3 level was relatively higher in advanced stage of human CRC tumors (F = 5.26, Pr = 0.00155) (Fig. 1C). In summary, we suggest that circCCT3 may act as an oncogene in CRC.

circCCT3 is upregulated in clinical CRC tissues and promotes CRC cell metastasis.
circCCT3 depletion inhibits CRC cell metastasis
To determine the role of circCCT3 on CRC metastasis, our group created stable circCCT3-knockdown HCT116 and SW480 cell lines by the shRNA method. RT-qPCR showed significantly reduced circCCT3 expression in the CRC cells (Fig. 1D). Next, we tested whether circCCT3 affected invasion of HCT116 and SW480 cells by transwell assay. The result showed that circCCT3 knockdown in HCT116 and SW480 cells led to smaller number of invaded cells (Fig. 1E). Furthermore, we used immunofluorescence in HCT116 and SW480 cells to examine apoptosis with Annexin V staining. We found that the number of Annexin V-positive sh-circCCT3 cells was more than that of sh-NC cells (Fig. 1F, G). Taken together, our data indicate that circCCT3 knockdown suppresses invasion and induces apoptosis of CRC cells.
miR-613 directly targets and regulates circCCT3
To make clear how circCCT3 regulates CRC metastasis, we first predicted the interacted microRNA of circCCT3 by online program StarBase v2.0 (Fig. 2A). Luciferase reporter assay indicated that miR-613 mimic reduced luciferase activity of cells with wild-type circCCT3-fused luciferase gene, whereas miR-613 inhibitor strikingly enhanced the luciferase activity (Fig. 2B). To validate this result, MS2-RIP experiment was performed and showed direct binding of miR-613 by wild-type circCCT3, not mutant circCCT3 (Fig. 2C).
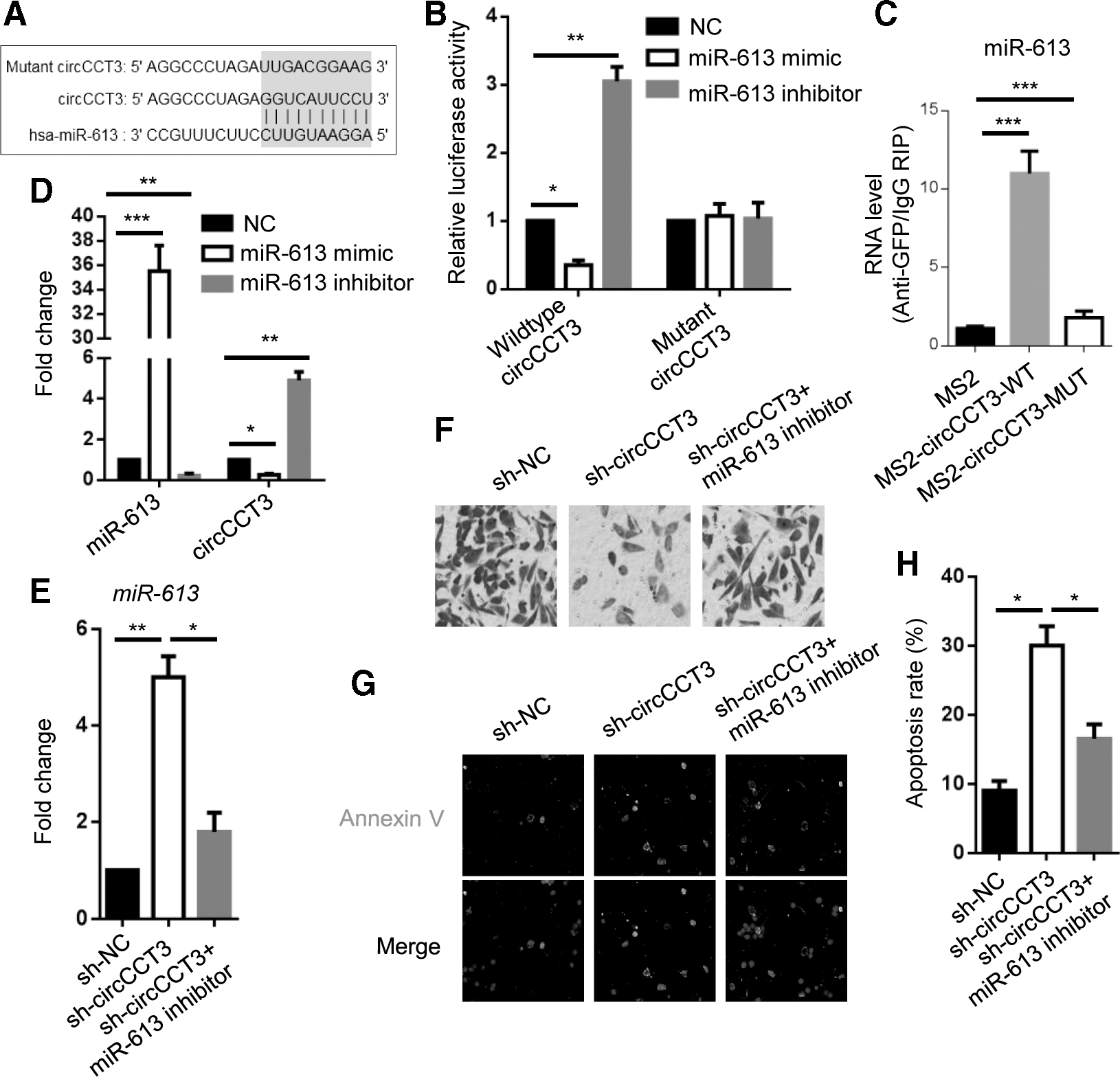
miR-613 mediates circCCT3 knockdown-attenuated CRC cell metastasis.
Afterward, we sought to detect circCCT3 level in miR-613 mimic or inhibitor HCT116 cells, miR-613 mimic greatly decreased circCCT3 expression level to ∼20–30%, whereas miR-613 inhibitor increased circCCT3 level by approximately four- to fivefolds (Fig. 2D). Taken together, our results suggest that miR-613 can directly target circCCT3.
miR-613 mediates circCCT3 knockdown-attenuated CRC cell metastasis
To investigate whether miR-613 mediated circCCT3-regulated metastasis of CRC cells, miR-613 inhibitor was transfected into sh-circCCT3 HCT116 cells (Fig. 2E). Transwell assay revealed enhanced invasion ability of circCCT3-knockdown HCT116 cells transfected with miR-613 inhibitor compared with sh-circCCT3 group cells (Fig. 2F). Immunofluorescence demonstrated that the number of Annexin V-positive HCT116 cells transfected with sh-circCCT3 plus miR-613 inhibitor was smaller than that of sh-circCCT3 cells (Fig. 2G, H). In conclusion, miR-613 is a key factor for mediating invasion and apoptosis of CRC cells regulated by circCCT3 knockdown.
miR-613 directly targets WNT3 and VEGFA
To identify the potential targets of miR-613, we employed online program TargetScan to perform prediction. The paired binding sequences were shown between miR-613 and WNT3 or VEGFA (Fig. 3A, C). To validate this result, we utilized luciferase reporter assay to examine the luciferase activity of HCT116 cells transfected with indicated vectors. The results showed that miR-613 mimic reduced the luciferase activity in HCT116 cells with wild-type WNT3 or VEGFA 3′-UTR-fused luciferase gene; by contrast, miR-613 inhibitor caused opposed trend (Fig. 3B, D). Next, we used western blot to detect Wnt3 and VEGFA protein levels in HCT116 cells transfected with miR-613 mimic or inhibitor. miR-613 mimic significantly attenuated Wnt3 or VEGFA expression, whereas miR-613 inhibitor greatly contributed to their levels (Fig. 3E). Taken together, our results suggest that miR-613 negatively affects Wnt3 and VEGFA expression.

miR-613 directly targets WNT3 and VEGFA.
WNT3 and VEGFA partially rescue miR-613-regulated CRC cell metastasis
Since Wnt signaling and VEGFA are reported to be important for a variety of cancers, we proposed that Wnt3 and VEGFA are involved in miR-613-modulated HCT116 cell metastasis. RT-qPCR analyses showed overexpression of WNT3 or VEGFA in miR-613 mimic HCT116 cells (Fig. 4A). The numbers of invaded Wnt3 or VEGFA-overexpressed miR-613 mimic HCT116 cells were increased in comparison to miR-613 mimic cells (Fig. 4B). Likewise, Wnt3 or VEGFA overexpression in miR-613 mimic HCT116 cells largely reduced apoptosis of miR-613 mimic HCT116 cells (Fig. 4C, D). Collectively, our data indicate that Wnt signaling and VEGFA are implicated in circCCT3/miR-613-regulated metastasis of CRC cells.
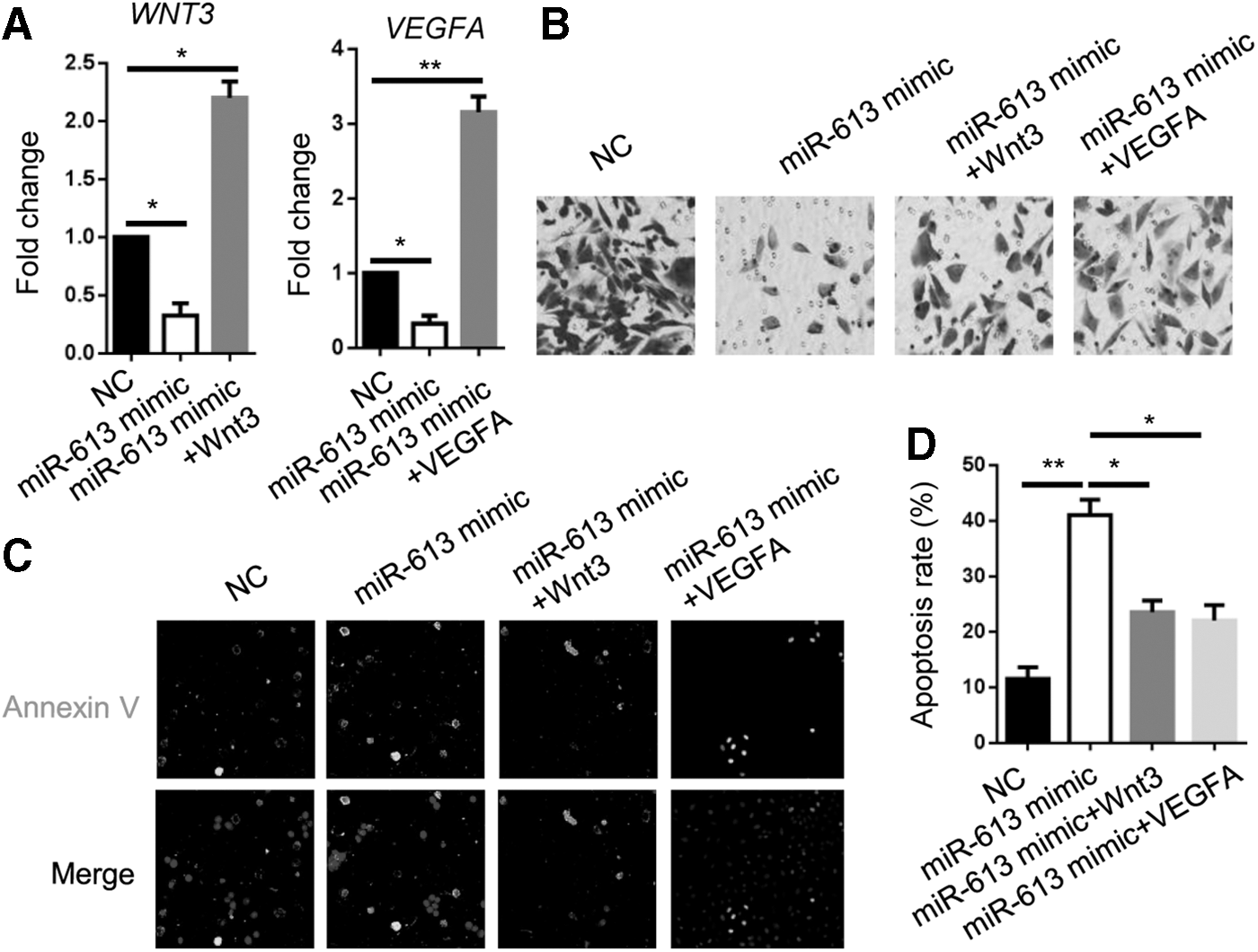
WNT3 and VEGFA partially rescue miR-613-regulated CRC cell metastasis.
Discussion
circCCT3 plays an oncogenic role in CRC metastasis through miR-613/VEGFA and Wnt signaling. Mechanistically, circCCT3 directly interacts with miR-613, then regulates VEGFA and WNT3 genes expression. Phenotypically, circCCT3 depletion attenuates invasion and induces apoptosis of CRC cells through miR-613/WNT3 or VEGFA. Bioinformatics show that circCCT3 is closely associated with CRC progression in clinical setting.
Cancer metastasis is widely considered as the major cause of cancer-related death (Shan et al., 2018). CRC metastasis to distant sites (most commonly is the liver) is generally regulated by many biological molecules (Cassidy and Syed, 2017; Kristensen et al., 2017). Currently, traditional methods of surgical resection, chemotherapy, and radiotherapy are most used in clinical patients. Thus, it is urgent to develop effective and targeted therapeutics for CRC. circRNAs have been shown as biomarkers for colorectal and other types of cancers (Meng et al., 2017; Li et al., 2018). CircITCH and circFOXO3 are reduced in esophageal squamous cell carcinoma and breast cancer (Li et al., 2015; Yang et al., 2016). In this study, we reveal that circCCT3 is upregulated and associated with malignancy of CRC. Besides, circCCT3 knockdown impairs invasion ability and increases apoptosis of CRC cells. Hence, we identify another circRNA as promoter for CRC metastasis. Unfortunately, animal experiment was not performed in our study to further confirm that CRC metastasis was regulated by circCCT3, which should be carried out in the future.
Previous studies have indicated that circRNA could act as competing endogenous RNA (ceRNA) of miRNA sponge to inhibit its biological function (Zhang et al., 2017; Ma et al., 2019). For instance, Huang et al. (2016) suggested that circMYLK binds miR-29a-3p and regulates DNMT3B, VEGFA, and ITGB1 in bladder cancer. CircZNF609 regulates AKT3 by functioning as ceRNA of miR-150-5p in Hirschsprung's disease (Peng et al., 2017). Similarly, our group demonstrates that circCCT3 directly interacts with miR-613 by luciferase reporter assay and bioinformatic analysis. Subsequently, circCCT3 serves as ceRNA of miR-613 to influence the CRC phenotype. Recently, in CRC, Wang et al. (2019) showed that circPVT1 promotes metastasis through miR-145. So, it is a common molecular phenomenon that circRNA functions as ceRNA for miRNA in CRC cells.
miRNAs are efficiently diagnostic and prognostic markers for cancers, including CRC (To et al., 2018). In basic researches, a number of groups have proposed that miRNAs regulate CRC tumorigenesis and metastasis through various signaling pathways. For example, miR-126 downregulation leads to PI3K/AKT signaling in CRC (Guo et al., 2008). miR-18a and miR-30b act as tumor suppressors via inhibiting KRAS (Tsang and Kwok, 2009; Liao et al., 2014). Additionally, miR-21 and miR-106a target TGF-β/Smad pathway to promote CRC migration and invasion (Feng et al., 2012; Yu et al., 2012). In our research, we investigate the role of miR-613 in CRC invasion and apoptosis. The results confirm tumor-suppressive role of miR-613 in CRC and serve as downstream effector for circCCT3 to mediate tumor phenotypes of CRC cells.
It is suggested that high levels of WNT pathway activity are observed in colon tumors (Cancer Genome Atlas Network, 2012; Novellasdemunt et al., 2015). We validate that miR-613 downregulates WNT3 expression level in HCT116 cells. In addition, transwell assay and Annexin V immunofluorescence test that WNT3 is the downstream target and restores the repressive role of miR-613 in CRC, which is consistent with previous study (Ying et al., 2008).
Conclusions
In summary, our research group explores the metastatic effects of circCCT3 on CRC. We identify a completely novel mechanism by which circCCT3 enhances invasion and induces apoptosis of CRC cells. We believe that targeting this regulation is a promising strategy for treating CRC.
Authors' Contributions
W.L., Youqi Xu, and Xiaodong Wang: designed and conducted the experiments. G.C., W.B., and Xia Wang: performed bioinformatic analyses. Z.F., Yao Xu, and Youqi Xu: wrote the article. M.D. and Q.T.: prepared the figures. All authors approved the final submission.
Footnotes
Disclosure Statement
No competing financial interests exist.
Funding Information
This work was supported by the Natural Science Foundation of Jiangsu Province (Basic Research Program, Grant No. BK20161605).