
Abstract
Long noncoding RNA growth-arrest-specific transcript 5 (GAS5) has been proved to play a crucial role in cancer chemoresistance. However, the function of GAS5 and its underlying molecular mechanism in hepatocellular carcinoma (HCC) chemoresistance remain unknown. In this study, we aimed to investigate its function and underlying molecular mechanism in HCC cisplatin (CDDP) resistance. The results demonstrated that GAS5 was significantly downregulated in HCC tissues and cells, especially in CDDP-resistant HCC tissues and cells. Low GAS5 expression was tightly correlated with shorter survival in patients with HCC. Functionally, GAS5 overexpression sensitized CDDP-resistant HepG2/CDDP and Huh7/CDDP cells to CDDP. Mechanically, GAS5 improved the sensitivity of HCC cells to CDDP through sponging miR-222. Taken together, these observations suggested that overexpression of GAS5 overcame CDDP resistance of HCC cells by regulating miR-222, providing a potential therapeutic target for overcoming the chemoresistance of HCC cells.
Introduction
Hepatocellular carcinoma (HCC), as one of the most common malignancies, threatens peoples' health and has an extremely high mortality rate (El-Serag and Mason, 1999; Shen et al., 2016; Zhang et al., 2017). Despite remarkably therapeutic advances that have been made in HCC, the prognosis of patients with HCC is still poor due to the acquisition of multidrug resistance (Lebwohl and Canetta, 1998). Cisplatin (CDDP) is one of the most common and effective chemotherapeutic drugs in the treatment of various cancers (Giaccone, 2000; Galanski, 2006). However, CDDP-based chemotherapy is often limited by intrinsic and acquired resistance, which ultimately leads to chemotherapy failure (Koberle et al., 2010; Johnsson et al., 2014). Therefore, it is urgent to understand the pathogenesis of HCC chemoresistance and develop novel effective therapeutic strategies for improving the outcome of HCC patients.
Long noncoding RNAs (lncRNAs) are nonprotein-coding RNAs with more than 200 nucleotides (Martens-Uzunova et al., 2014; Sun et al., 2015). Recently, aberrant expression of lncRNAs is observed in multiple cancers, and their roles in tumorigenesis, progression, and drug resistance have been elucidated (Palmieri et al., 2017; Lin and Yang, 2018). Growth-arrest-specific transcript 5 (GAS5), located at chromosome 1q25, was originally identified in a subtraction complementary DNA (cDNA) library prepared for sequencing RNA expressed in growth arrested cells (Schneider et al., 1988). Subsequently, accumulating documents have indicated that GAS5 is commonly downregulated in various malignancies, such as lung cancer, gastric cancer, and ovarian cancer (Zhang et al., 2016; Tan et al., 2017; Zhao et al., 2018). Growing evidence suggests that GAS5 was linked to cell growth and metastasis and played a crucial role in tumor progression and chemoresistance (Liu et al., 2018; Chen et al., 2019; Huo and Chen, 2019). Moreover, GAS5 was reported to be downregulated and act as an independent prognostic factor for patients with HCC (Tu et al., 2014). However, limited knowledge is available concerning the functional role and underlying mechanism of GAS5 in HCC CDDP resistance.
MicroRNAs (miRNAs), as a new class of small (18–25 nucleotides) noncoding RNAs, exert their roles by directly pairing to the 3′ untranslated region of the mRNAs, leading to inhibition of mRNA translation and degradation (Carrington and Ambros, 2003; Bartel, 2004; Guo et al., 2015). It has been shown that lncRNAs could interact with miRNAs and then modulate the expression of miRNA target genes (Wang et al., 2014; Feng et al., 2017). For instance, GAS5 might function as a competing endogenous RNA (ceRNA) to upregulate anti-metastasis protein ANGPTL1 expression through sponging miR-182, thus decreasing migratory and invasive abilities of HCC cells (Chen et al., 2019). Moreover, GAS5 could overcome CDDP resistance in many cancers through functioning as miRNA sponges (Cao et al., 2017; Wen et al., 2017; Yao et al., 2019). However, whether GAS5 could regulate CDDP resistance in HCC through regulating miRNA expression remains unclear.
In this study, our purpose was to investigate the functional role of GAS5 and its underlying molecular mechanism in the resistance of HCC cells to CDDP. Our research discovered that GAS5 was downregulated in HCC tissues and cells. Our research found that GAS5/miR-222 axis might be a target for overcoming CDDP resistance in HCC.
Materials and Methods
Tissue samples and cells
We collected 38 paired tumor and adjacent normal tissues from HCC patients who underwent surgical resection at the First Affiliated Hospital of Henan University. The detail information of patients was shown in Supplementary Table S1. This study was approved by the Ethics Committee of the First Affiliated Hospital of Henan University (20140316). All patients signed the informed consent.
Human normal liver cell line (Lo-2) and human liver cancer cell line (HepG2 and Huh7) were obtained from the Chinese Academy of Sciences Cell Bank (Shanghai, China). All cell lines were cultured in RPMI-1640 medium (HyClone, Logan, UT) supplemented with 10% fetal bovine serum (Gibco) at 37°C with 5% CO2. The CDDP-resistant HepG2/CDDP and Huh7/CDDP cells were established by exposing their parent cells to increasing concentrations of CDDP for 12 months.
Cell transfection
Empty pcDNA3.0 vector (Vector) and GAS5 overexpression vector pcDNA3.0-GAS5 (GAS5) were synthesized by GeneCopoeia (Guangzhou, China). miR-con, miR-222, anti-miR-con, or anti-miR-222 was purchased from GenePharma (Shanghai, China). The cells were cotransfected with Vector or GAS5 and miR-con or miR-222 or anti-miR-con or anti-miR-222 using Lipofectamine 2000 (Invitrogen, Shanghai, China).
Quantitative real-time PCR
We extracted total RNA from HCC tissues or cells using TRIzol reagent (Invitrogen). Total RNA was reverse transcribed into cDNA using PrimeScript RT Reagent Kit (TaKaRa, Tokyo, Japan). Quantitative real-time PCR (qRT-PCR) was performed to determine the expression of GAS5 and miR-222. Glyceraldehyde 3-phosphate dehydrogenase (GAPDH) and U6 were used as the internal normalizers for GAS5 and miR-222, respectively. Then the relative expression of GAS5 and miR-222 was calculated by 2−ΔΔCt method.
CDDP sensitivity assay
To determine the viability of cells after CDDP treatment, MTT assay was performed. The transfected cells were seeded in 96-well plates. After 24 h of incubation, HepG2/CDDP and Huh7/CDDP cells were treated with different concentrations (0.5, 1, 2, 4, 8, 16, 32, 64 μg/mL) of CDDP for 48 h. Finally, optical density value at 490 nm wavelength was measured. The sensitivity of HCC cells to CDDP was assessed using the half of the maximum inhibitory concentration (IC50) of CDDP.
Flow cytometry
Flow cytometry was used to assess cell apoptosis. The transfected HepG2/CDDP and Huh7/CDDP cells were incubated with 2 μg/mL of CDDP for 48 h. Then cells were collected and washed with 1 × phosphate buffered saline. The cells were then stained with 5 μL of Annexin V-fluorescein isothiocyanate and 5 μL propidium iodide for 10 min at room temperature in the dark. After that, the apoptosis of the cells was measured by fluorescence activated cell sorting.
Comet assay
Comet assay was performed as described previously (Zeng et al. 2018). Briefly, HepG2/CDDP and Huh7/CDDP cells that received CDDP treatment were analyzed by DNA Damage Detection Kit (KeyGEN BioTECH, China). Images were visualized using a Leica fluorescence microscope. Tail length was measured using CometScore software (TriTek Corporation, Sumerduck, VA).
Luciferase reporter assay
The luciferase reporters containing the wild-type GAS5 (GAS5-WT) or mutant-type GAS5 (GAS5-MUT) were generated to analyze the interaction between GAS5 and miR-222. HepG2/CDDP and Huh7/CDDP cells were seeded in a 24-well plate for 24 h. Cells were cotransfected with above luciferase plasmids and miR-222 mimics or miR-con using Lipofectamine 2000 (Invitrogen). At 48 h post-transfection, the luciferase activity was measured using the Dual Luciferase Reporter Assay System (Promega).
Western blot analysis
Western blotting was performed as described previously (Lei et al., 2007). The antibodies against vascular endothelial growth factor (VEGF), VEGFR2, and GAPDH were purchased from Cell Signaling Technology (Danvers, MA).
Statistical analysis
All data were described as mean ± standard deviation from at least three independent experiments. Statistical analysis of experimental results was carried out using SPSS 20.0 (IBM, NY) software. A p-value of <0.05 was considered statistically significant.
Results
lncRNA GAS5 was downregulated in CDDP-resistant HCC tissues and cells
To investigate the functional role of GAS5 in HCC, the expression level of GAS5 in HCC tumor tissues (n = 38) and adjacent normal tissues (n = 38) was evaluated by qRT-PCR analysis. The result showed that the GAS5 expression was obviously downregulated in HCC tissues compared with that in adjacent normal tissues (Fig. 1A). Then, the expression of GAS5 in tumor tissues of CDDP-sensitive and CDDP-resistant HCC patients was further detected by qRT-PCR. Consistently, the results showed that the expression of GAS5 was remarkably decreased in CDDP-resistant patients compared with that in CDDP-sensitive patients (Fig. 1B). In addition, to assess GAS5 expression in HCC cell lines, qRT-PCR was performed in HCC parental cells (HepG2 and Huh7), their CDDP-resistant cells (HepG2/CDDP and Huh7/CDDP), and human normal liver cell line (Lo-2). Compared with Lo-2 cells, GAS5 expression was significantly lowered in HepG2 and Huh7 cells (Fig. 1C, D). More importantly, the expression of GAS5 in HepG2/CDDP and Huh7/CDDP cells was lower than those in their parent cells (Fig. 1C, D). Kaplan–Meier survival analysis showed that the survival rate of HCC patients with high CACS2 expression level was higher than those with low GAS5 expression level (p = 0.0089) (Fig. 1E). Taken together, these results suggested that GAS5 may be involved in CDDP resistance in HCC.

Expression of GAS5 was downregulated in HCC tissues and cells.
GAS5 overexpression enhanced the sensitivity of CDDP-resistant HCC cells to CDDP
To evaluate whether GAS5 overexpression overcame CDDP resistance of HepG2/CDDP and Huh7/CDDP cells, IC50 value of CDDP was determined by MTT assay in HepG2/CDDP and Huh7/CDDP and their parental cells. As demonstrated in Figure 2A and B, the IC50 values of CDDP were obviously improved in HepG2/CDDP and Huh7/CDDP cells than those in HepG2 and Huh7 cells. To deeply investigate the functional roles of GAS5 in CDDP-resistant HCC cells, we established two stably GAS5-overexpressing resistant cells by transfecting vector (GAS5) into HepG2/CDDP and Huh7/CDDP cells, respectively. The results of qRT-PCR analysis showed that the expression levels of GAS5 were demonstrably increased in GAS5-transfected HepG2/CDDP and Huh7/CDDP cells (Fig. 2C, D). As expected, the results revealed that GAS5 overexpression remarkably improved the sensitivity of HepG2/CDDP and Huh7/CDDP cells to CDDP (Fig. 2E, F). Next, we evaluated the effects of GAS5 on CDDP-induced apoptosis in HepG2/CDDP and Huh7/CDDP cells using flow cytometry. As shown in Figure 2G and H, HepG2/CDDP and Huh7/CDDP cells transfected with GAS5 exerted a significant promotion in CDDP-induced apoptosis compared with cells introduced with Vector. Subsequently, DNA damage was measured by comet assay. Similarly, GAS5 overexpression generated a remarkable increase in DNA strand break damage and comet tail percentage in HepG2/CDDP and Huh7/CDDP cells (Fig. 2I, J). These findings indicated that overexpression of GAS5 could overcome CDDP resistance in CDDP-resistant HCC cells.
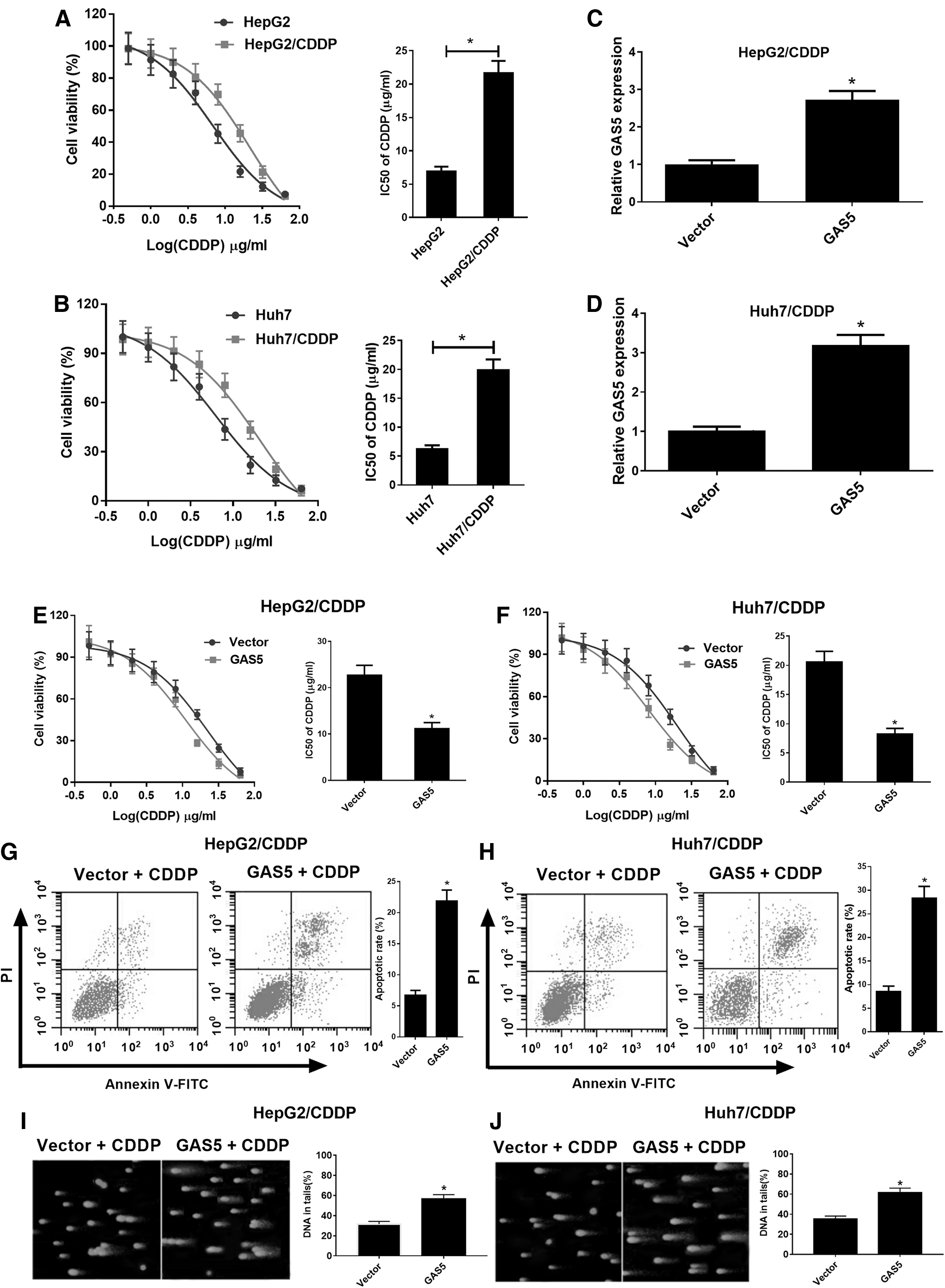
GAS5 overexpression enhanced CDDP sensitivity of HepG2/CDDP and Huh7/CDDP cells.
GAS5 sponges miR-222 in HCC cells
To confirm the mechanism of GAS5-mediated CDDP resistance in HCC cells, we identified miR-222 as a potential target of GAS5. The online software miRcode was used to calculate the miRNA targets of GAS5. The predicted result showed that GAS5 sequence contained a miR-222 binding site (Fig. 3A). To validate the accuracy of prediction results, luciferase reporter plasmids containing GAS5-WT or GAS5-MUT were transfected into HepG2/CDDP and Huh7/CDDP cells together with miR-222 or miR-con. The results indicated that the luciferase activity of GAS5-WT reporter vector was markedly suppressed by miR-222 overexpression (Fig. 3B, C), but the luciferase activity of GAS5-MUT reporter vector was unaffected (Fig. 3B, C). To further explore the regulatory role of GAS5 in miR-222 expression, qRT-PCR analysis was conducted in HepG2/CDDP and Huh7/CDDP cells transfected with Vector, GAS5, or GAS5-MUT. Notably, miR-222 expression was markedly inhibited by transfection of GAS5 but not GAS5-MUT in HepG2/CDDP and Huh7/CDDP cells (Fig. 3D, E). As shown in Figure 4F, the relative level of miR-222 expression in HCC tumor tissues was significantly higher than that in adjacent normal tissues. Spearman's correlation analysis discovered that miR-222 expression was negatively correlated with GAS5 expression in HCC tumor tissues (Fig. 3G). In summary, these results suggested that GAS5 could act as a miRNA sponge to regulate miR-222.
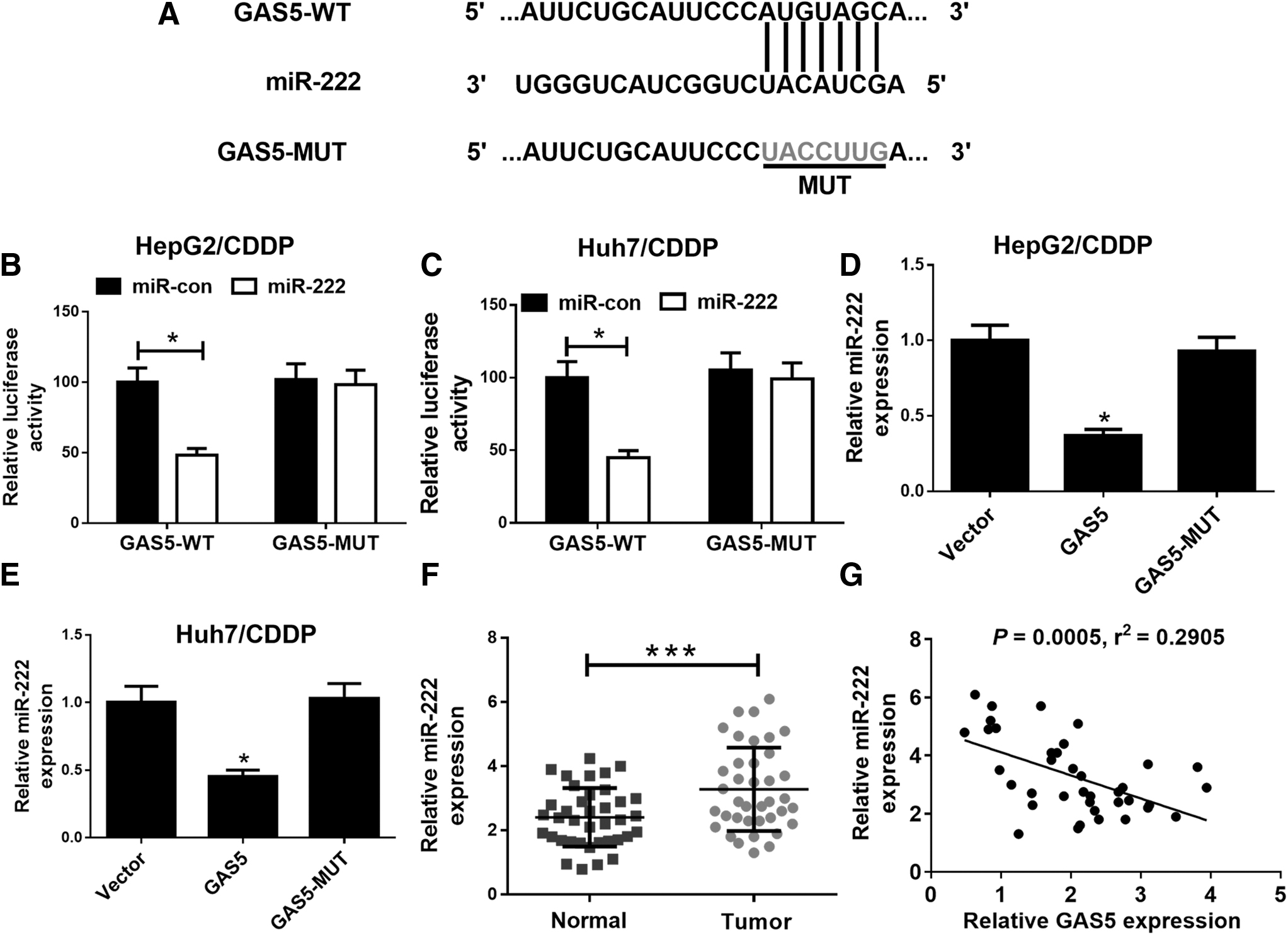
GAS5 acted as a miR-222 sponge in HCC cells.

GAS5 overexpression overcame CDDP resistance in CDDP-resistant HCC cells through suppressing miR-222 expression. HepG2/CDDP and Huh7/CDDP cells were transfected with Vector or GAS5 and miR-con or miR-222 or anti-miR-con or anti-miR-222, followed by determination of miR-222 expression by qRT-PCR analysis
Overexpression of GAS5 overcame CDDP resistance in CDDP-resistant HCC cells through suppressing miR-222 expression
The above results indicated that miR-222 can directly bind to GAS5, and we further explored the role of CACS2 in enhancing the sensitivity of HCC cells to CDDP through regulating miR-222 expression. Result of qRT-PCR showed that transfection of anti-miR-222 could inhibit miR-222 expression in HepG2/CDDP and Huh7/CDDP cells. miR-222 overexpression abolished the inhibitory effect of GAS5 on miR-222 expression in HepG2/CDDP and Huh7/CDDP cells (Fig. 4A, B). The results of MTT assay demonstrated that inhibition of miR-222 improved the sensitivity of HepG2/CDDP and Huh7/CDDP cells to CDDP (Fig. 4C, D). We further determined the effect of GAS5 and miR-222 on CDDP-induced apoptosis. Flow cytometry analysis was conducted in HepG2/CDDP and Huh7/CDDP cells exposed to 2 μg/mL CDDP. As shown in Figure 4E and F, anti-miR-222 transfection significantly increased cell apoptosis induced by CDDP in HepG2/CDDP and Huh7/CDDP cells. Upregulation of miR-222 blocked GAS5-induced HepG2/CDDP and Huh7/CDDP cell apoptosis (Fig. 4E, F). These findings demonstrated that overexpression of GAS5 improved CDDP sensitivity in CDDP-resistant HCC cells through suppressing miR-222 expression.
Overexpression of GAS5 decreased the activity of VEGF signaling pathway in CDDP-resistant HCC cells
To explore the relevant signal pathway, we performed western blotting to measure the expression levels of VEGFR2 and VEGF in Vector or GAS5 transfected HepG2/CDDP and Huh7/CDDP cells with CDDP exposure. Results showed that GAS5 upregulation dramatically decreased the expression levels of VEGF and VEGFR2 proteins in HepG2/CDDP and Huh7/CDDP cells (Fig. 5A, B). Collectively, GAS5 overcame CDDP resistance in CDDP-resistant HCC cells through inactivation of VEGF signaling pathway.
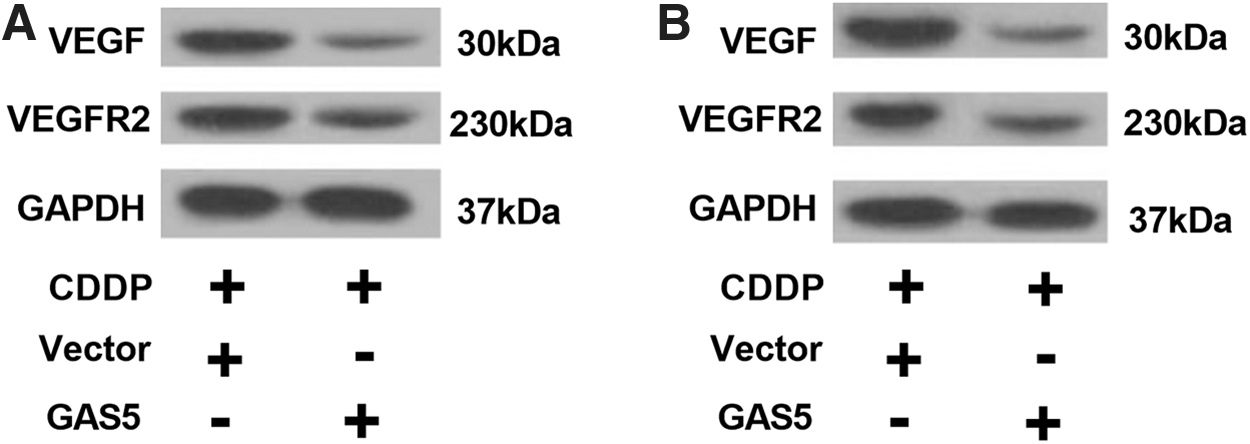
GAS5 overexpression decreased the activation of VEGF signaling pathway in CDDP-resistant HCC cells. HepG2/CDDP and Huh7/CDDP cells were transfected with Vector or GAS5 and 2 μg/mL CDDP.
Discussion
The roles of dysregulated lncRNAs in the chemoresistance of various cancers have received recent attention (Wang et al., 2012, 2019; Zhao et al., 2016; Jiang et al., 2018; Jin et al., 2018). Studies have suggested that GAS5 plays a key role in CDDP resistance. In the present study, we discovered that the expression of GAS5 was significantly decreased in CDDP-resistant HCC tissues and cells. HCC patients with low GAS5 expression have a shorter survival time, which prompted us to further study the potential role of GAS5 in the chemotherapy of HCC. CDDP treatment also resulted in a decreased expression of miR-222 in human HCC cells. We observed that GAS5 overexpression enhanced CDDP sensitivity of HepG2/CDDP and Huh7/CDDP cells by promoting cell apoptosis. Hence, upregulation of GAS5 could enhance the sensitivity of HCC cells to CDDP, providing a promising therapeutic target for the CDDP resistance of HCC.
Elucidating the molecular mechanism underlying CDDP resistance contributed to developing reasonable and effective therapies to overcome CDDP resistance. Our results revealed that GAS5 was downregulated in HCC tissues and cells. GAS5 is a verified tumor suppressor gene and has been found to be downregulated in many cancers (Zhang et al., 2016; Tan et al., 2017; Zhao et al., 2018). Apart from our findings, previous studies reported that abnormal expression of GAS5 was implicated in chemoresistance in cancers. For instance, GAS5 acted as a ceRNA to elevate STAT3 expression by sponging miR-21, subsequently enhancing the sensitivity of cervical cancer to CDDP (Yao et al., 2019). Overexpression of GAS5 enhanced the sensitivity of glioma cells to CDDP through the activation of mTOR signaling (Hou and Chen, 2019). Furthermore, GAS5 functioned as a ceRNA by sponging miR-21 to increasing PTEN expression in nonsmall cell lung cancer to overcome CDDP resistance of tumor cells (Cao et al., 2017). Our study demonstrated that overexpression of GAS5 contributed to the sensitivity of HepG2/CDDP and Huh7/CDDP cells to CDDP. The precise mechanism by which downregulation of GAS5 led to CDDP resistance in liver cancer remains unclear. Hence, we identified the functional role of GAS5/miR-222 axis in the sensitivity of HCC cells to CDDP. ceRNA hypothesis indicated that lncRNAs functioned as miRNA sponges to repress the function of miRNA (Wang et al., 2017; Jiang et al., 2018), and the online software miRcode was utilized to predict the potential target miRNAs of GAS5. Growing evidence suggested that miR-222 functioned as a key oncogene. For instance, miR-222 overexpression might be a key contributor to tumorigenesis (Wong et al., 2010). Moreover, miR-21, miR-31, miR-122, miR-221, and miR-222 in liver cancer were significantly upregulated (Galardi et al., 2007; Karakatsanis et al., 2013). Recent research demonstrated that acquired CDDP resistance was also associated with upregulated miR-222 (Jiang et al., 2014). miR-222 overexpression dramatically antagonized CDDP-induced autophagy in bladder cancer cells, attenuating CDDP-induced cell death (Zeng et al., 2016). In the present study, our data showed that GAS5 acted as a miR-222 sponge in HepG2/CDDP and Huh7/CDDP cells. Overexpression of miR-222 reversed the inductive effect of GAS5 upregulation on the sensitivity of HepG2/CDDP and Huh7/CDDP cells to CDDP. We also showed the inverse association of miR-222 and GAS5 expression in HCC tissues and cells. VEGF signaling pathway was reported to be involved in CDDP resistance (Liao et al., 2017). Notably, our finding revealed that GAS5 could downregulate VEGF signaling pathway in HCC cells. All these data demonstrated that GAS5 overexpression sensitized CDDP-resistant HCC cells to CDDP through sponging miR-222 and inactivating VEGF signaling pathway in HCC.
Conclusion
In conclusion, we identified the key role of GAS5 in regulating CDDP resistance in HCC cells and its underlying mechanism. Overexpression of GAS5 enhanced the CDDP sensitivity of HCC cells to CDDP through sponging miR-222, providing a promising therapeutic strategy to overcome CDDP resistance in HCC treatment.
Ethics Approval and Consent to Participate
This study was approved by the Ethics Committee of the First Affiliated Hospital of Henan University (20140316).
Footnotes
Disclosure Statement
No competing financial interests exist.
Funding Information
This work was supported by the Functional and Molecular Imaging Innovative Technology Team Project of Kaifeng (201709).
Supplementary Material
Supplementary Table S1
References
Supplementary Material
Please find the following supplemental material available below.
For Open Access articles published under a Creative Commons License, all supplemental material carries the same license as the article it is associated with.
For non-Open Access articles published, all supplemental material carries a non-exclusive license, and permission requests for re-use of supplemental material or any part of supplemental material shall be sent directly to the copyright owner as specified in the copyright notice associated with the article.