
Abstract
Long noncoding RNAs (lncRNAs) play vital roles in development and progression of various cancers. To investigate the relationship between three tag single-nucleotide polymorphisms (SNPs) (rs13252298, rs1016343, and rs1456315) in lncRNA prostate cancer-associated noncoding RNA 1 (PRNCR1) and lung cancer (LC) risk, we conducted this study. First, we performed a case–control study, including 576 LC patients and 612 cancer-free controls. Second, a meta-analysis was used to evaluate the association of selected SNPs with risk of overall cancer. We found that rs13252298 and rs1456315 were strongly correlated with risk of LC, nonsmall cell lung cancer (NSCLC), and lung adenocarcinoma. For rs13252298, individuals carrying GG genotype had increased risks of LC compared with those carrying AA genotype (adjusted odds ratio [OR] = 1.565, 95% CI = 1.091–2.245, p = 0.015). A significant result was also found in recessive model with adjusted OR of 1.719. Individuals with GG genotype of rs1456315 were at increased risks of LC compared with those carrying AA genotype. Similar results were found in NSCLC patients. Meta-analysis showed that rs1016343 and rs13252298 were associated with overall cancer. But for rs1016343, no significant association was observed in Asians. In conclusion, rs13252298 and rs1456315 in PRNCR1 may be genetic susceptibility factors for LC in Chinese population. These results need to be confirmed by further studies.
Introduction
Lung cancer (LC), the leading cause of cancer morbidity and mortality in both men and women, is one of the most common malignancies worldwide (Torre et al., 2016). Using the GLOBOCAN 2018 compiled by the International Agency for Research on Cancer, researchers concluded that there would be an estimated 2.1 million new LC cases and 1.8 million deaths predicted in 2018, representing close to one in five (18.4%) cancer deaths in the world (Bray et al., 2018). Similarly, it is also the leading cause of cancer deaths in Chinese men and women, and is a major public health problem. Chen et al. (2016) used high-quality data from the existing population registry of the National Central Cancer Registry of China to observe that trends in mortality rates of LC (age standardized to the SEGI standard population) were stable in the whole population. It is well known that cancer is affected by both genetic and environmental factors (Du et al., 2020; Gu et al., 2020; Yang et al., 2020), as is LC (Romaszko and Doboszynska, 2018; Qin et al., 2019). In addition, studies have reported that many single-nucleotide polymorphisms (SNPs) in long noncoding RNA (lncRNA) genes were associated with multiple cancer susceptibility. Therefore, they were potential biomarkers for predicting cancer risk (Lv et al., 2017).
LncRNAs are a class of noncoding RNAs (ncRNAs) >200 nucleotides in length, containing 200 to 100,000 nucleotides. Lacking a specific and complete open reading frame, lncRNA is an endogenous transcribed RNA molecule located in the nucleus or cytoplasm with a number of 7000 to 23,000 in vivo (Moran et al., 2012; Parolia et al., 2015). As a vital regulator of various biological functions and disease processes, lncRNA has complex secondary and tertiary structure, which could bind to proteins, RNA, and DNA and exert its regulatory functions (Xu et al., 2017a). It is dysregulated in many cancer types and is thought to play a key role in regulating several characteristics of cancer biology.
Prostate cancer-associated noncoding RNA 1 (PRNCR1), first found in prostate cancer by Chung et al. (2011) based on genome-wide association studies (GWASs) in 2011, is a ncRNA ∼13 kilobase introns. PRNCR1, also known as PCAT8 and CARLo3, is located in the “gene desert” region of chromosome 8q24 (128.14–128.28 Mb). The chromosome 8q24 region contains numerous regulatory factors, including enhancers and superenhancers. And it has been reported that chromosome 8q24 has become an important region of genetic susceptibility to various human diseases and cancers (Barry et al., 2014). PRNCR1 was upregulated in some prostate cancer cells and prostatic intraepithelial neoplasia lesions. In recent years, with the in-depth study of lncRNAs, lncRNA PRNCR1 has gradually been found to be related not only to prostate cancer (Wang et al., 2013) but also to osteolysis after hip replacement (Gong et al., 2018), eclampsia (Jiao et al., 2018), colorectal cancer (Wu et al., 2016), gastric cancer (Zhang et al., 2020), breast cancer (Pang et al., 2019), and so on. In 2018, Cheng et al. (2018) demonstrated that PRNCR1 upregulated HEY2 by competitive binding to miR-448, promoting progression of nonsmall cell lung cancer (NSCLC). In the study, miR-448 was highly expressed in NSCLC tissues and cell lines. In addition, low expression of miR-448 could predict poor prognosis in patients with NSCLC. Then, to study the mechanism of lncRNA PRNCR1 and NSCLC, and explore the effect of PRNCR1 on LC cell migration and apoptosis, Wang et al. (2019) used lentiviral transfection technology to construct lncRNA PRNCR1 shRNA interference expression vector in 2019. The results showed that shRNA lncRNA PRNCR1 could reduce the migration rate of NSCLC cells and increase the apoptosis rate. These findings provided innovative ideas and methods for the treatment of LC, and they also indicated that the PRNCR1 gene had certain research value in LC.
As a newly discovered lncRNA in recent years, PRNCR1 affecting tumor cell proliferation, invasion, and apoptosis has been gradually discovered, and it has become a research hotspot for early diagnosis and treatment of various cancers. Many studies have confirmed that SNPs in PRNCR1, such as rs13252298, rs1016343, and rs1456315, were associated with the risk of multiple cancers (Teerlink et al., 2016; Chu et al., 2017; Huang et al., 2018). Based on the role of PRNCR1 in the development of LC and the regulation of SNPs on gene expression function of lncRNAs, we hypothesized that SNPs in PRNCR1 might alter the risk of LC. Therefore, we conducted a hospital-based case–control study to investigate the relationship between three tag SNPs (rs13252298, rs1016343, and rs1456315) in PRNCR1 and LC susceptibility. In addition, a systematic and comprehensive meta-analysis was used to explore the relationship between these variants and overall cancer susceptibility.
Materials and Methods
Study subjects
This hospital-based case–control study was carried out in Shenyang, located in Liaoning province in China. This study was approved by the Institutional Review Board of China Medical University, and all subjects signed written informed consent. The case group including 576 newly histologically diagnosed LC patients was from the First Affiliated Hospital of China Medical University, the Fourth Affiliated Hospital of China Medical University, and the General Hospital of the Northern War Zone of the Chinese People's Liberation Army from January 2011 to December 2013. All LC patients were diagnosed by histological diagnosis before radiotherapy and chemotherapy. The inclusion and exclusion criteria of LC cases and the control subjects were the same as reported previously (Yang et al., 2018b). A total of 612 cancer-free controls were derived from healthy people who were in the same hospital for medical examination at the same time.
We used Quanto1.2.4 statistical software in conjunction with the sample size calculation formula to calculate the sample size. The frequency of the control group was matched by the age of the case ±5 years old. All subjects donated 5 mL of venous blood, and an epidemiological survey was conducted on the subjects. Moreover, background information such as gender, age, and tobacco exposure was collected. We recorded all the problems encountered during the investigation and produced a manual containing detailed collection, analysis, preservation, and transportation. Nonsmokers were defined as individuals with a total number of lifetime smoking of <100.
SNP selection and genotyping
We selected three tag SNPs (rs13252298, rs1016343, and rs1456315) in PRNCR1, which were obtained from the database of 1000 Genomes Project for the Chinese Han Beijing (CHB) population by using haploview4.2 software and reading previous studies on cancer susceptibility to PRNCR1 gene polymorphisms. HaploRegv4.1 database was used to predict the potential function of candidate SNPs in PRNCR1 (Shen et al., 2020). The minimum allele frequencies of these three SNPs were >0.05. Anticoagulant genomic DNA was extracted from venous blood of all subjects using the classical phenol-chloroform method. Genotyping of three SNPs conducted by using an Applied Biosystems 7500 FAST Real-Time PCR System (Foster City, CA) utilizing TaqMan® allelic discrimination (Applied Biosystems) with commercial primer-probe set. For quality control, 10% of samples were randomly selected for repeated genotyping, and the results of duplicate sets were concordant.
Statistical analysis
Student's t-test, χ 2-test, and logistic regression analysis were used to examine the differences in age, gender, and smoking status between the LC case group and the cancer-free control group. The goodness-of-fit χ 2-test was performed to test the Hardy–Weinberg equilibrium (HWE) of the genotypes. Unconditional logistic regression analysis was used to calculate the odds ratios (ORs) and their 95% confidence intervals (95% CIs) to assess the relationship between rs13252298, rs1016343, and rs1456315 and risk of LC. The linear-by-linear association of χ2-test was conducted to calculate cumulative effects of variant alleles rs13252298-G and rs1456315-G. The interaction between smoking exposure and these three loci polymorphisms and LC susceptibility was analyzed by using the additive interaction model and the multiplicative interaction model. Statistical power was calculated by using Quanto1.2.4 (University of Southern California, Los Angeles, CA). All results were statistically analyzed using SPSS25.0 (IBM SPSS, Inc., Chicago, IL) statistical software. The tests were bilateral, and the test level was set to 0.05. If p ≤ 0.05, it was considered statistically significant. Haploview4.2 software was used to select tag SNPs and conduct the analysis of linkage disequilibrium (LD) to get LD plot. When r 2 > 0.8, it was considered that there was a LD.
Methodology of meta-analysis
Relevant literature was obtained by a comprehensive search in PubMed, Web of Science, and CNKI databases up to July 20, 2020. The search strategy was “PRNCR1 OR PCAT8” AND “polymorphism OR mutation OR variant OR rs13252298 OR rs1016343 OR rs1456315” AND “cancer OR carcinoma OR tumor OR neoplasms.” Eligible studies need to meet the following inclusion criteria: (1) original studies; (2) case–control studies evaluating the relationship between rs13252298, rs1016343, and rs1456315 and cancer susceptibility; (3) studies with sufficient genotype frequency data to calculate pooled ORs and 95% CIs. Exclusion criteria were as follows: (1) studies excluded were not case–control studies on these three variants and cancer risk; (2) duplicate documents; (3) meeting papers, abstracts, case reports, reviews, and meta-analysis; and (4) those lacking valid data. Data extraction and literature quality assessment were conducted independently by two researchers. When differences were encountered, they resolved them through negotiation. The data to be extracted were as follows: first author, year, country, ethnicity, source of control, type of cancer, genotyping methods, number of cases and controls, genotype frequencies in cases and controls, p value of HWE in controls, and the score of Newcastle–Ottawa Scale (NOS). Studies with NOS greater than five were included. All statistical analyses were performed using Stata 12.0 software, and p < 0.05 was considered statistically significant. We used random-effects model to calculate pooled ORs and 95% CIs under five different genetic models. The heterogeneity was determined by the results of χ 2 based Q-test and I 2-test. Moreover, the funnel plot, Begg's test, and Egger's test were used to detect publication bias. Sensitivity analysis was implemented by eliminating any single study by turns.
Results
Baseline characteristics
The basic information of subjects is shown in Table 1. There were 576 LC patients, including 263 lung adenocarcinoma (AD), 208 lung squamous cell carcinomas (SQ), nine large cell lung carcinomas, 87 small cell lung cancer, and nine others. The average age of cases was 58.77 ± 12.40 (mean ± standard deviation [SD]). The results of Student's t-test and χ 2-test showed no significant differences in the distribution of age between the case and control group (p = 0.065 and p = 0.944, respectively). Likewise, the gender distribution of two groups was not statistically significant (p = 0.829). However, there was a statistically significant difference in smoking status between two groups (p = 0.000). Obviously, the smoking rate in cases was higher than that in controls. There were 612 cancer-free controls, and their average age was 57.32 ± 14.55 (mean ± SD). Table 2 shows the basic information and allele frequencies of PRNCR1 SNPs in this study. The genotype frequency distributions of these SNPs were consistent with HWE in the control group (p = 0.428 for rs13252298; p = 0.598 for rs1016343; p = 0.933 for rs1456315). LD plot of three tag SNPs is shown in Figure 1.
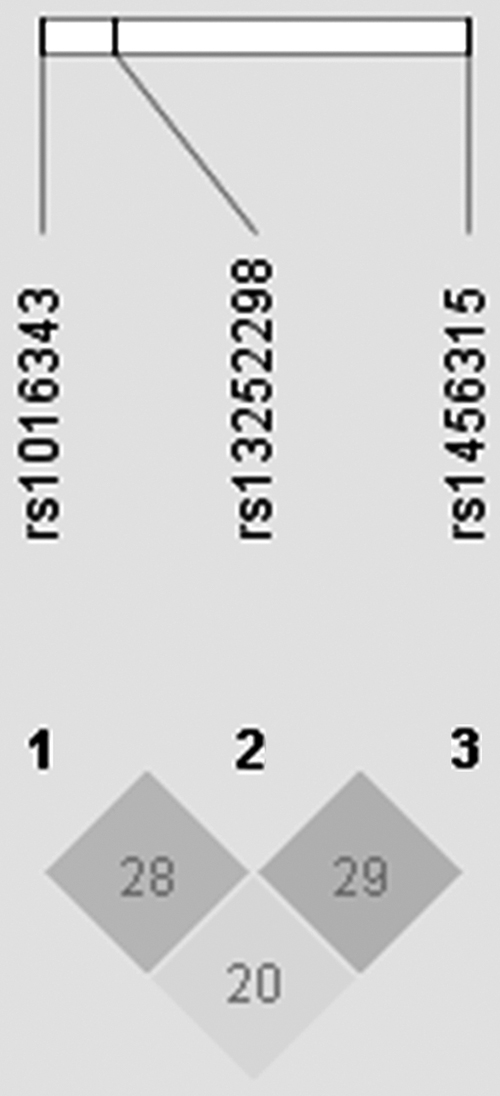
LD plot about LD for the three tag SNPs (rs13252298, rs1016343, and rs1456315) in PRNCR1. Numbers in squares indicate 100-fold r 2 values for each pair of SNPs. LD, linkage disequilibrium; PRNCR1, prostate cancer-associated noncoding RNA 1; SNP, single-nucleotide polymorphisms.
Demographic Characteristics of Lung Cancer Cases and Control Groups
p < 0.05 indicates statistical significance; bold values indicate that p < 0.05.
AD, lung adenocarcinoma; LCLC, large cell lung cancer; SCLC, small cell lung cancer; SD, standard deviation; SQ, lung squamous cell carcinoma.
Basic Information and Allele Frequencies of Prostate Cancer-Associated Noncoding RNA 1 Single-Nucleotide Polymorphisms in This Study
Adjusted by gender and smoking status.
Adjusted by gender and smoking status; p < 0.05 indicates statistical significance.
HWE, Hardy–Weinberg equilibrium; MAF, Minor allele frequency; OR, odds ratio; SNP, single-nucleotide polymorphisms; 95% CI, 95% confidence interval.
Genotype distribution and LC susceptibility
Table 3 and Supplementary Table S1 list the relationship between these three SNPs and susceptibility to LC and its subtypes. PRNCR1 rs1016343 was found to be in no significant association with risk of LC, NSCLC, AD, and SQ in all models. However, we found that rs13252298 and rs1456315 variants were significantly associated with LC risk (GG vs. AA: adjusted OR = 1.565, 95% CI = 1.091–2.245, p = 0.015 and GG vs. AG+AA: adjusted OR = 1.719, 95% CI = 1.226–2.410, p = 0.002 for rs13252298; GG vs. AA: adjusted OR = 1.488, 95% CI = 1.015–2.180, p = 0.042 and GG vs. AG+AA: adjusted OR = 1.484, 95% CI = 1.032–2.134, p = 0.033 for rs1456315). In addition, strong associations were found between them and NSCLC risk (Table 3). Moreover, rs13252298 and rs1456315 polymorphisms were strongly related to the risk of AD. For rs13252298, individuals carrying the G allele had a greater risk of AD and SQ than that carrying the A allele.
Association Between the Three Single-Nucleotide Polymorphisms and Risk of Lung Cancer and Nonsmall Cell Lung Cancer
Bold values indicate that p < 0.05.
Adjusted by age, gender, and smoking status.
Adjusted by age, gender, and smoking status; p < 0.05 indicates statistical significance.
As shown in Table 4, compared with carriers with three or less risk alleles of the two SNPs, individuals with four alleles had significantly increased risk of LC and NSCLC (LC: adjusted OR = 1.934, 95% CI = 1.171–3.192, p = 0.010; NSCLC: adjusted OR = 2.235, 95% CI = 1.342–3.723, p = 0.002). However, cumulative effects were not found in a dose-dependent manner (LC: p = 0.684; NSCLC: p = 0.954).
Cumulative Effects of rs13252298-G and rs1456315-G on Lung Cancer and Nonsmall Cell Lung Cancer Susceptibility
Adjusted by age, gender, and smoking status.
Calculated by linear-by-linear association of χ 2-test.
Relationship of SNPs in PRNCR1 with LC risk stratified by smoking status, age, and gender
By stratifying smoking status, we obtained that SNPs in PRNCR1 had no relationships with LC and NSCLC susceptibility in ever smoking and never smoking groups, except rs13252298 in never smoking group (Supplementary Table S2). In population of ≤60 age group, rs13252298 was significantly associated with risk of LC and NSCLC (Supplementary Table S3). These results were obtained similarly to results of >60 age group. In female stratified population, rs13252298 and rs1456315 mutations could significantly promote the occurrence of LC and NSCLC (Supplementary Table S4). Yet, we did not find that PRNCR1 SNPs were meaningfully associated with the risk of LC and NSCLC in male population, and rs1016343 had the same results in female population.
Interaction between SNPs and smoking exposure
In Supplementary Table S5, we observe never smoking status with a protective genotype of two SNPs (AG+AA genotype of rs13252298, AA genotype of rs1456315) as the reference group to investigate the gene–environmental interaction, separately. The results of the crossover analysis revealed that smoking exposure with both protective and dangerous genotypes significantly increased the risk of LC and NSCLC compared with never smoking with protective genotypes. These results implied that there might be gene–environmental interactions, so, we need to further validate them using multiplicative model and additive model. Though, we failed to find meaningful results of the gene–environmental interaction on the additive scales (Supplementary Table S6). However, the OR value of multiplicative interaction of rs13252298 risk genotypes with smoking exposure less than one indicated that there was a negative multiplicative interaction, with a p value of 0.039 (p < 0.05) (Supplementary Table S7).
Results of Meta-Analysis
Figure 2 shows the literature selection process. Finally, a total of 15 articles containing this study were included (Salinas et al., 2008; Zheng et al., 2010; Chung et al., 2011; Ho et al., 2012; Li et al., 2013, 2015; Hui et al., 2014; Chang, 2016; He et al., 2017; Sattarifard et al., 2017; Xu et al., 2017b; Yang et al., 2018a; AlMutairi et al., 2019; Hong et al., 2019), including 11 on rs1016343, 11 on rs13252298, and seven on rs1456315. The main characteristics of eligible studies in this meta-analysis are shown in Table 5. After statistical analysis, we indicated that rs1016343 and rs13252298 were related to overall cancer susceptibility, while rs1456315 had nothing to do with the risk of overall cancer (Table 6 and Fig. 3). In the subgroup analysis, we found that rs1016343 was associated with increased overall cancer susceptibility of Caucasian population, while no association was found with cancer susceptibility of Asian population. The sensitivity analysis results revealed that this study was stable and reliable (Fig. 4). As shown in Figure 5, the shape of Begg's funnel plots looked relatively symmetrical. Moreover, the results of Begg's test confirmed that there was no publication bias in this meta-analysis (p = 0.876 for rs1016343; p = 0.64 for rs13252298; p = 0.23 for rs1456315).
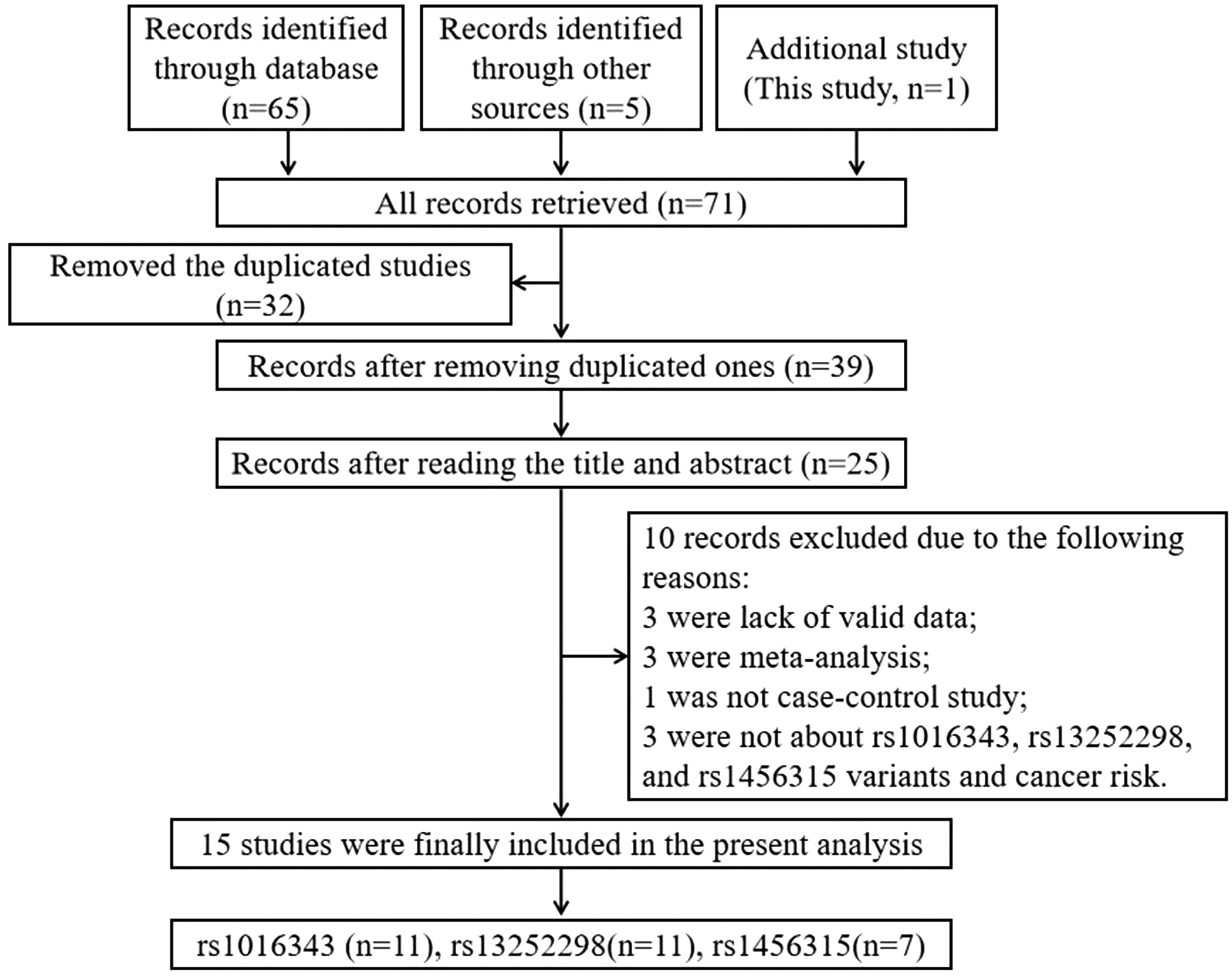
Flowchart illustrates the study selection process of this meta-analysis.
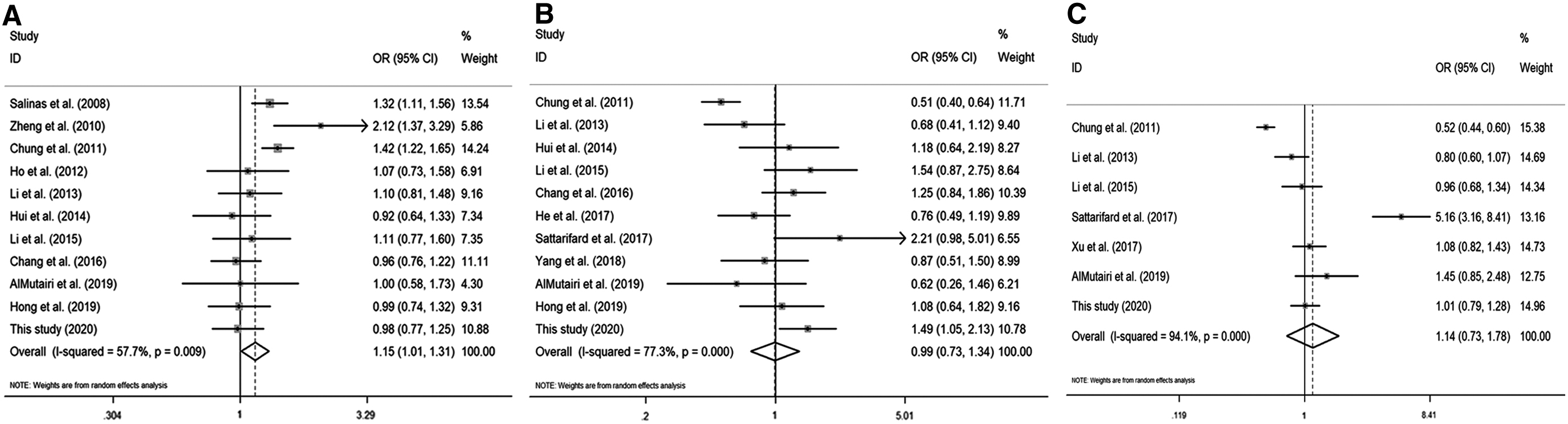
Forest plots of cancer risk associated with the PRNCR1 polymorphisms.

Sensitivity analysis for the meta-analysis of PRNCR1 gene polymorphisms and cancer risk.
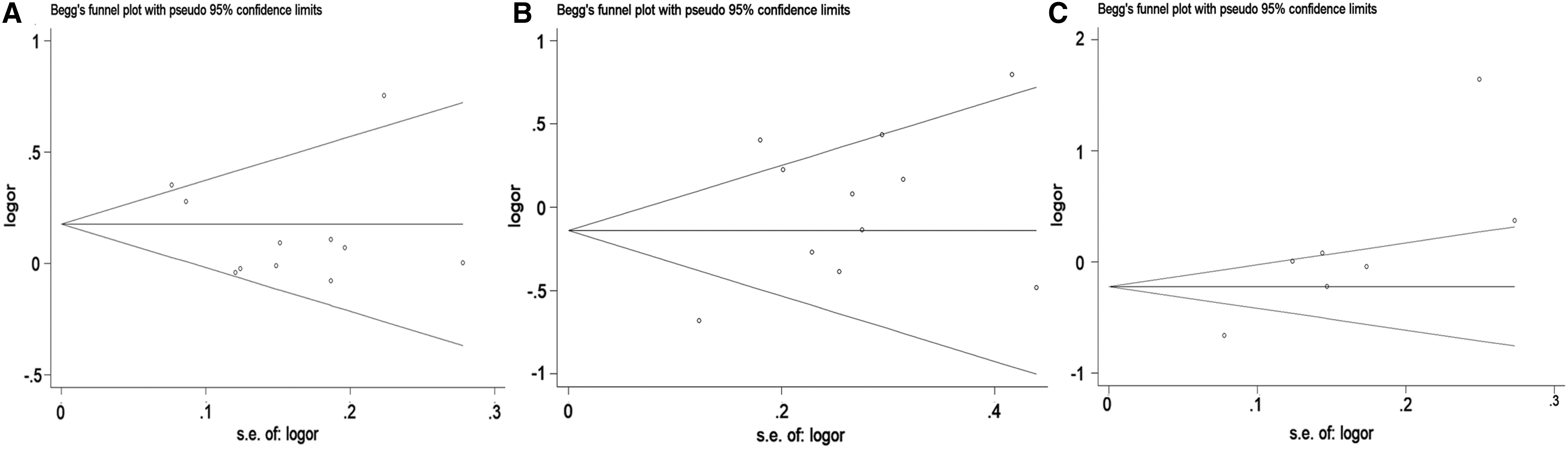
Begg's funnel plots of PRNCR1 polymorphisms and cancer risk.
Meta-Analysis of the Association of PRNCR1 Polymorphisms with Risk of Cancer
Bold values in the table indicate that OR is statistically significant.
Main Characteristics of Eligible Studies in This Meta-Analysis
HB, hospital based; NOS, Newcastle–Ottawa assessment scale; PB, population based; PCR-HRM, polymerase chain reaction-high resolution melting curves; PCR-RFLP, polymerase chain reaction-restriction fragment length polymorphism; PHC, primary hepatic carcinoma.
Discussion
This is the first case–control study preliminarily reporting the relationship between SNPs in PRNCR1 and LC susceptibility in Chinese northeast population. We found no statistically significant association between rs1016343 and susceptibility to LC and its subtypes. However, rs13252298 and rs1456315 were found to significantly increase susceptibility to LC, NSCLC, and AD. The results of the crossover analysis provided the risk of LC increased by smoking exposure and the dangerous genotype carriers of these two SNPs. Moreover, the interaction of rs13252298 risk genotypes with smoking exposure was a negative multiplicative interaction. Results of meta-analysis shed light on that rs1016343 could increase risk of overall cancer, but no significant association was observed in Asians. The overall cancer risk of individuals with rs13252298 AG genotype was 0.822 times higher than that with AA genotype (p = 0.021). However, no association was found between rs1456315 and overall cancer susceptibility.
Several studies have shown that ncRNAs had multiple biological behaviors during tumorigenesis and progression. As two subgroups of the ncRNAs family, microRNAs (miRNAs) could interact with lncRNAs to form a ceRNA network (Chen et al., 2018). LncRNAs could regulate many biological processes of human cancer through various mechanisms, thereby affecting the development of cancer. Researchers confirmed that PRNCR1 could promote the progression of NSCLC. Functionally, miR-448 had an anticancer effect in NSCLC by inhibiting cell proliferation, invasion, migration, and epithelial–mesenchymal transition. In terms of mechanism, lncRNA PRNCR1 exerted ceRNA function in NSCLC by regulating miR-448 and HEY2. In addition, PRNCR1 was highly overexpressed in invasive prostate cancer and might be an essential component of anticastration of prostate tumors (Yang et al., 2013). Moreover, the upregulation of PRNCR1 promoted cell proliferation and cell cycle progression in colorectal cancer (Yang et al., 2016). PRNCR1-siRNA could downregulate the expression of PRNCR1, effectively inhibited the proliferation and invasion and migration of gastric cancer cells, and laid a theoretical foundation for gene therapy of gastric cancer with PRNCR1 as a target (Cai, 2017).
SNPs in lncRNAs might affect the splicing process and the stability of the mRNA conformation, leading to modification of its interacting partners. Therefore, further identification of lncRNA PRNCR1-related SNPs might open a new way for functional analysis of cancer susceptibility loci identified by GWAS (Li et al., 2013). Since PRNCR1 is located in 8q24 region of chromosomes, whether its genetic variation is associated with multiple cancers has become a hot topic in recent years (Han et al., 2016; Xu et al., 2017b). Based on these studies, Salinas et al. (2008) conducted a population-based study of Caucasians and African Americans, which showed that rs1016343 was significantly associated with Caucasian prostate cancer risk. However, studies have also shown that rs1016343 was not associated with susceptibility to prostate cancer in Scottish population, gastric cancer in Korean population, and colorectal cancer in Chinese population (Ho et al., 2012; Li et al., 2013; Hong et al., 2019). Similarly, rs1016343 was found to be unrelated to LC susceptibility in Chinese northeast population in this case–control study. And, results of this meta-analysis showed that no significant association was observed between rs1016343 and over cancer in Asians. For rs13252298, related studies have shown that it was not statistically associated with gastric cancer risk in Chinese population and Korean population (He et al., 2017; Yang et al., 2018a; Hong et al., 2019). However, it was found to be associated with a reduction in colorectal cancer in Chinese population but not in the risk of colorectal cancer in Saudi population (Li et al., 2013; AlMutairi et al., 2019). For rs1456315, it was connected with a reduced risk of colorectal cancer in Chinese population, but it could significantly increase the risk of colorectal cancer in Saudi population (Li et al., 2013; AlMutairi et al., 2019). In our case–control study, the mutant GG genotypes of rs13252298 and rs1456315 were more likely to increase the risk of LC than the wild homozygous AA genotypes in a Chinese northeast population. Therefore, the mutant GG genotypes were dangerous genotypes of rs13252298 and rs1456315 in LC. In addition, the interaction of rs13252298 GG risk genotype with smoking exposure was statistically significant and was a negative multiplicative interaction. But, rs1456315 was not related to overall cancer risk in our meta-analysis. The controversial results of these studies might be caused by insufficient sample size, different races, and different genotyping methods.
After reviewing relevant literature, we found that this is the first case–control study to investigate the SNPs in PRNCR1 and the susceptibility to LC in China. However, this study also has certain limitations. First, the research objects in Shenyang may not be representative. Second, our sample size may not be large enough, especially after stratification analysis. Third, this experiment is only a related study, and there is no deeper functional research. Therefore, we should not only explore more PRNCR1 polymorphisms and other aspects of cancers, including chemotherapy susceptibility, metastasis, and recurrence, but also through functional studies to explore the potential of PRNCR1 polymorphisms in tumorigenesis mechanism in the future research. Hence, a larger number of studies should be conducted in the future to verify our conclusions.
Conclusion
In summary, rs13252298 and rs1456315 variants in PRNCR1 were associated with risk of LC in this case–control study. Moreover, the interaction of the rs13252298 GG risk genotype with smoking exposure was statistically significant and was a negative multiplicative interaction. Besides, our meta-analysis revealed that rs1016343 and rs1456315 were not related to over cancer risk in Asians subgroup, except for rs13252298. Thus, SNPs in PRNCR1 may influence genetic susceptibility to LC and NSCLC in Chinese northeast population. All of these results might need to be further validated by larger sample size studies.
Footnotes
Authors' Contributions
N.L. contributed to conceptualization, methodology, and writing original article; Z.C., M.G., S.L., M.S., Y.W., L.T., Y.B., Z.Z., S.W., and B.Z. contributed to methodology; Z.Y. contributed to review writing and editing, supervision, and funding acquisition.
Acknowledgments
The authors thank all the study subjects for their participation and personnel in the hospitals for their assistance.
Disclosure Statement
No competing financial interests exist.
Funding Information
This study was funded by the National Natural Science Foundation of China (No. 81673261).
Supplementary Material
Supplementary Table S1
Supplementary Table S2
Supplementary Table S3
Supplementary Table S4
Supplementary Table S5
Supplementary Table S6
Supplementary Table S7
References
Supplementary Material
Please find the following supplemental material available below.
For Open Access articles published under a Creative Commons License, all supplemental material carries the same license as the article it is associated with.
For non-Open Access articles published, all supplemental material carries a non-exclusive license, and permission requests for re-use of supplemental material or any part of supplemental material shall be sent directly to the copyright owner as specified in the copyright notice associated with the article.