
Abstract
Silicosis is an irreversible occupational pulmonary disease that is characterized as progressed pulmonary fibrosis. In this study, we investigated the changes of UFSP2 and the related UFMylation in silica-induced pulmonary injury mice models. The experimental silicosis models were prepared by intratracheal injection of silica particles, and the lung samples were harvested at the first or the seventh day after treatment. We found that the UFSP2 expression in the 1-day models was comparable, whereas it was upregulated in the 7-day models. Consistently, the UFMylation in the lung tissues of the 7-day models was activated. In addition, we observed the CADM2, an adhesion molecule, was reported to associate with epithelial–mesenchymal transition, was upregulated in the lungs of 7-day models. In contrast, it remained comparable in the 1-day models. Our data indicated that the UFSP2/UFMylation pathway and the CADM2 might be involved in the silica-induced pulmonary injury.
Introduction
Silicosis is an irreversible occupational disease that is caused by the inhalation of silica particles and characterized as the formation of silicotic nodule and progressive pulmonary fibrosis (Leung et al., 2012). The pathological mechanism of silicosis includes multiple progresses, such as cell death, inflammation, oxidative stress, exhaustion of macrophages, fibroblasts activation, disorders of collagen production, and deposition (Schmidt et al., 1984; Fubini and Hubbard, 2003; Joshi and Knecht, 2013; Kawasaki, 2015; Benmerzoug et al., 2018). Epithelial–mesenchymal transition (EMT) has been known to be a crucial mechanism for the uncontrolled activation of fibroblast (Willis et al., 2005; Yamauchi et al., 2010; Li et al., 2015). Recently, endoplasmic reticulum (ER) stress has been reported to be involved in the EMT of alveolar epithelial cells (AEC) (Delbrel et al., 2019). In addition, the ER stress and the related unfolded protein response have been linked to lung fibrosis by the regulation of AEC apoptosis, myofibroblast differentiation and the M2 macrophage polarization in idiopathic pulmonary fibrosis (Burman et al., 2018). Therefore, ER stress may be a potential target for the treatment of silicosis.
The UFMylation system is a ubiquitination-like pathway that promotes the conjugation of the UFM1 to its target protein (Gerakis et al., 2019). Although the biological function of the UFMylation remains to be investigated, it has been reported to be closely associated with the ER homeostasis and ER stress (Lemaire et al., 2011; Li, 2018b; Xie et al., 2019; Wang, 2020a). UFSP2 is a key enzyme for the activation of UFM1 by removing the C-terminal two amino acid residues and also mediate the de-UFMylation by removing the conjugated UFM1 from the target proteins (Komatsu et al., 2004; Kang et al., 2007; Ha et al., 2011). Therefore, the expression of UFSP2 is indispensable for the UFMylation and de-UFMylation. In this study, we explored whether UFSP2 and UFMylation were involved in silicosis.
Materials and Methods
Ethics statement and animal study
The animal experiments were approved by the Ethics Committee of Peking University People's Hospital (2018PHC069). Twenty C57BL/6J male mice about 6 weeks of age were weighed and randomly divided into two groups: silica injected group and phosphate-buffered saline (PBS)-injected group, 10 mice/group. Then the mice were housed under climate-controlled conditions in a 12 h light/12 h dark cycle and allowed free access to food and water. The silica-induced lung injury experiment was carried out as previously reported (Liu, 2019a). In brief, the silica particles (Sigma-Aldrich, St. Louis, MO) were baked at 180°C overnight for sterilization and suspended in sterile, lipopolysaccharide-free PBS at a concentration of 40 mg/kg and sonicated for 20 min with an ultrasonic oscillator (Kunshan Instrument Co., Jiangsu, China). The mice were anesthetized with pentobarbital sodium. The silica suspension was slowly administered through intratracheal injection at a dose of 0.15 g/kg, and the controlled mice were treated with equal volume of sterile PBS. The body weight changes were measured daily.
Histological analysis
The mice were sacrificed at the first and seventh day after treatment, respectively. The pulmonary tissues were cut and fixed in 10% of formalin. The tissues were embedded in paraffin, cut into 5 μm thick slices and stained with hematoxylin and eosin. For immunohistochemistry staining, the slices were heated with citrate buffer for 30 min and the endogenous peroxidases were blocked with 3% hydrogen peroxide. After being washed with PBS for three times, the slices were incubated with diluted primary antibodies: rabbit anti-CADM2 (Sigma-Aldrich) or UFSP2 (Abcam, Cambridge, MA) at 4°C overnight. Subsequently, the slices were washed with PBS for five times and incubated with HRP-tagged goat anti-rabbit secondary antibody (ZSGB-BIO, Beijing, China) at room temperature for 1 h. The slices were then washed with PBS for five times and stained with diaminobenzidine. The expressions of CADM2 and UFSP2 were viewed through a microscope.
Protein analysis
The expression of proteins in lung tissues was determined through western blotting. In brief, lung tissues were homogenized with RIPA lysis buffer and quantified by using a BCA method. The samples were mixed with loading buffer and boiled at 100°C for denaturation. Equal proteins were loaded onto an SDS-PAGE. The proteins were then separated by electrophoresis at 120 V for 1.5 h. The proteins were then blotted onto a nitrocellulose membrane at 300 mA for 2 h and blocked with 5% nonfat milk for 1 h at room temperature. Subsequently, the membrane was incubated in diluted primary antibodies: rabbit anti-CADM2 (Sigma-Aldrich), UFSP2, UFM1, UFL1, and GAPDH (Abcam) at 4°C overnight. After that, the membrane was washed with tris buffered saline with 0.05% Tween-20 (TBST) for four times and incubated with HRP-tagged secondary antibody for 1 h at room temperature. The membrane was then washed with TBST for four times, followed by detecting the bands with SuperSignal West Femto Maximum Sensitivity Substrate (Pierce, Rockford, IL). The band intensities were quantified with Image J software V1.52a (National Institute of Health, MD).
Statistical analysis
The statistical procedures were performed using SPSS19.0 software (SPSS, Inc., Chicago, IL). The data were expressed as means ± standard deviation. A two-tailed Student's t-test was used for analyzing differences between two groups. p < 0.05 was considered as statistically significant.
Results
UFSP2 is increased in silica-induced pulmonary injury
To investigate the expression of UFSP2 in silica-induced pulmonary injury, we initially construct the mice models by intratracheal injection of silica suspension. The lungs were collected at the first or the seventh day after treatment. The body weight changes are shown in Figure 1A. Data of histological analysis of the 7-day models indicated infiltration of inflammatory cells and the formation of silicotic nodules (Fig. 1B). By using western blotting, we found the expression of UFSP2 was significantly upregulated in the lung tissues of the 7-day models (Fig. 2A, B). However, UFSP2 was comparable in lung of 1-day models (Fig. 2A, B). Consistently, data from immunohistochemistry staining showed that UFSP2 was increased in the lung of silica-treated mice (Fig. 2C), suggesting that UFSP2 was involved in silica-induced pulmonary injury.

The construction of silica-induced pulmonary mice models (1-day model: silica group: n = 3, control group: n = 5; 7-days model: silica group: n = 3, control group: n = 5).
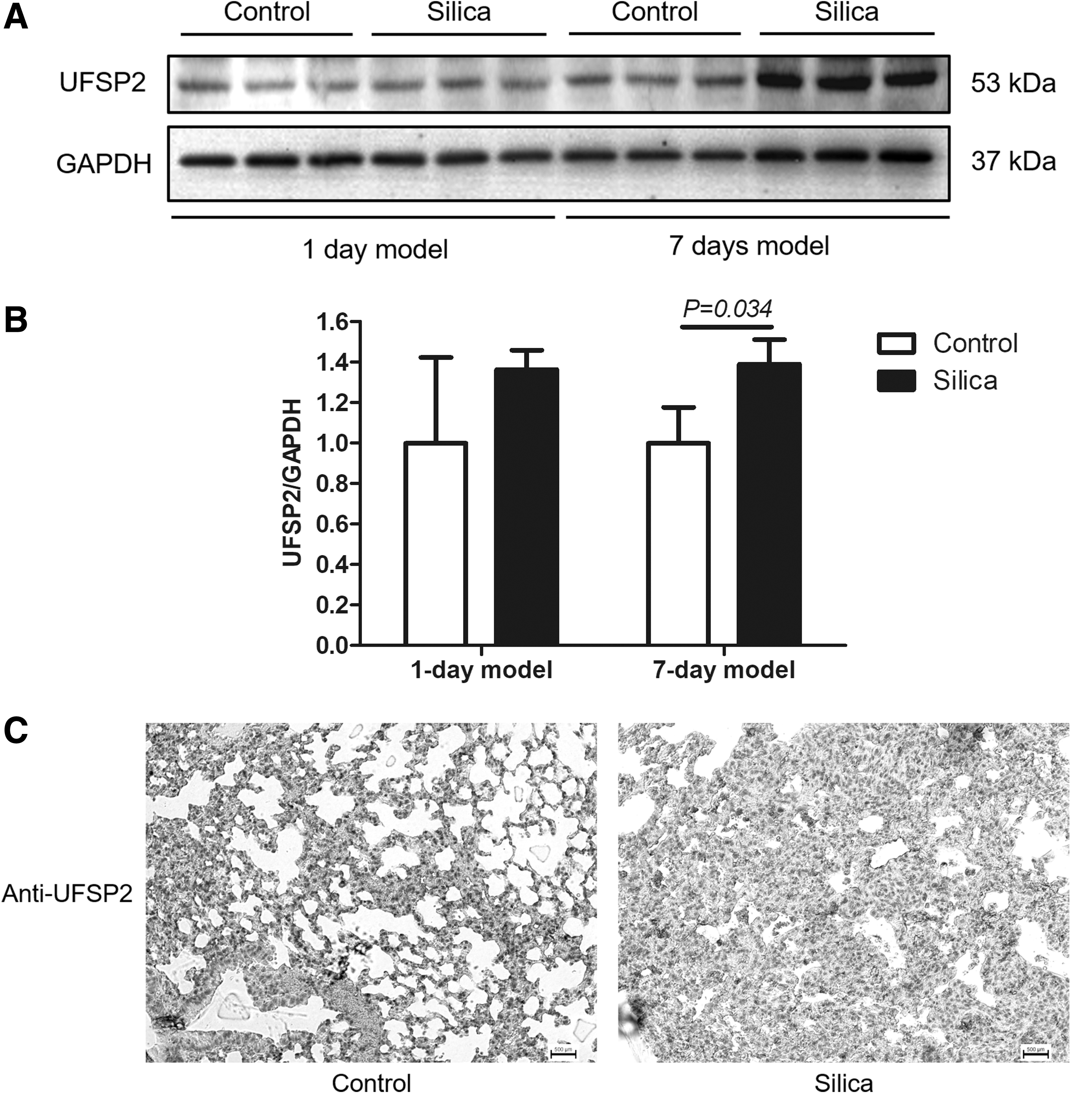
The expression of UFSP2 in silica-induced pulmonary mice models. The expression of UFSP2 was detected using western blotting
The UFMylation system is activated by silica inhalation
The UFSP2 plays multiple roles in the UFMylation system. By removing the two amino acid residues of the pro-UFM1, the UFSP2 enzyme catalyzes the maturation of UFM1 and thus promotes UFMylation, and UFSP2 also medicates the de-UFMylation process by the removal of the conjugated form of UFM1 from the modified proteins (Kang et al., 2007; Ha et al., 2011). In the lung tissues of 7-day silica-induced pulmonary injury models, the UFMylation was significantly upregulated (Fig. 3). However, there was no obvious change in the 1-day models. The expressions of UFL1, an UFM1 ligase (E3) of the UFMylation system, were comparable in either the 1-day or 7-day models (Fig. 3). These data indicated that UFMylation could be activated in silica-induced pulmonary injury.
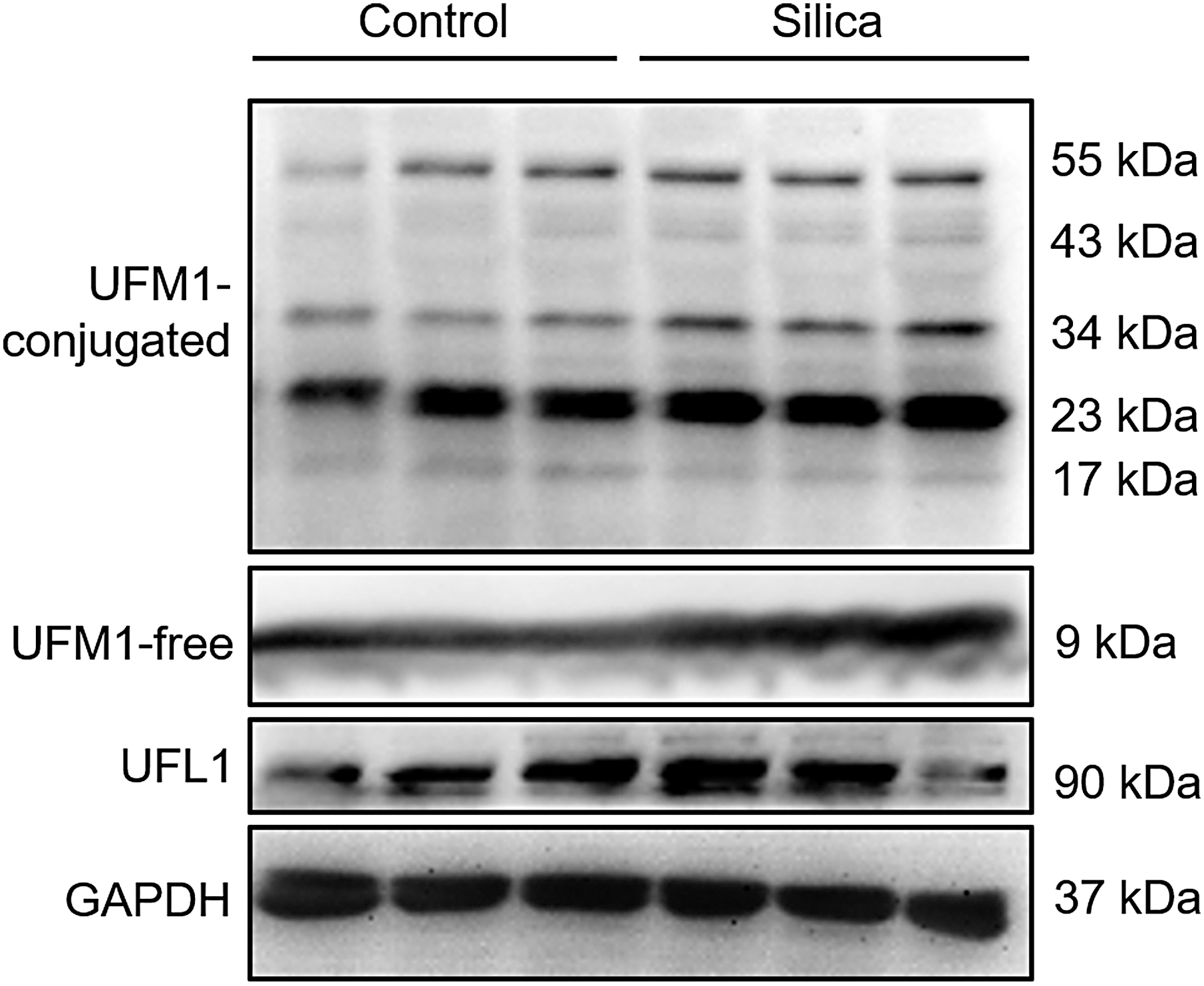
Silica inhalation activates UFMylation. The UFMylation status was represented with the conjugated form of UFM1. The UFM1 and the specific enzyme of UFMylation UFL1 was detected with western blotting. Each experiment was performed three times.
The expression of CADM2 is upregulated in silica-induced pulmonary injury
As EMT is a key process of pulmonary fibrosis, we next investigate the expression of CADM2, a molecule that is known to participate in the EMT regulation, in silica-induced pulmonary injury. In the 1-day mice models, the expression of CADM2 was not significant between the silicosis and the control mice (Fig. 4A). In contrast, the expression of CADM2 was significantly upregulated in the lungs of the 7-day silicosis mice (Fig. 4B, C). These data indicated that CADM2 may participate in silica-induced lung injury.
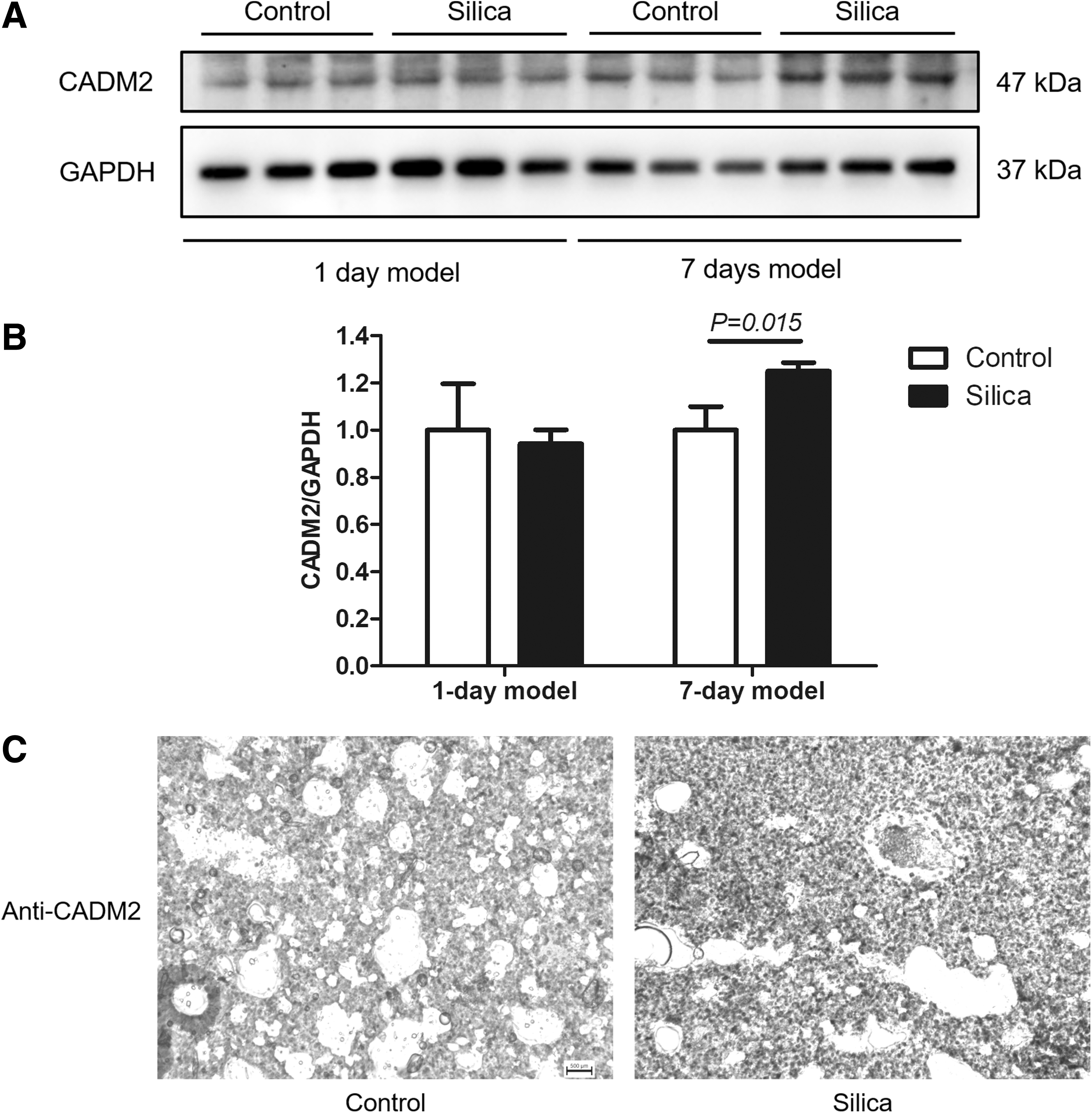
The expression of CADM2 in the silicosis models. The expression of CADM2 was determined with western blotting
Discussion
In this study, we analyzed the changes of UFSP2 and the associated UFMylation in silica-induced pulmonary injury. We found that the UFSP2 was significantly upregulated in the lung tissues of silica-treated mice. Consistently, the UFMylation was significantly activated. These data indicated that the UFSP2/UFMylation pathway might be involved in silicosis.
The UFMylation system is a novel identified post-translational modification that is known to associate with the ER homeostasis and ER stress (Liu et al., 2017; Wang, 2020a). It has been reported that UFMylation can be activated during ER stress, whereas inhibition of UFMylation induces ER stress (Lemaire et al., 2011; Cai et al., 2019). UFL1, the E3 enzyme of UFMylation, was reported to protect against heart failure (Li, 2018b). The disorder of UFMylation is also known in some cancers (Yoo et al., 2014; Wang, 2020b), and may associate with drug resistance (Terai et al., 2018). The histone H4 protein can be UFMylated, which participates in the DNA damage response (Qin et al., 2019). Recently, UFMylation was reported to participate in ER-phagy and protein quality control (Liang et al., 2020). Although the relationship between UFMylation and silicosis is unknown, ER stress is known to participate in the development of silicosis (Cheng et al., 2019). In this study, we, for the first time, observed that the UFMylation was activated in the silicotic lung. The UFSP2 is a required enzyme for the maturation of UFM1 by splicing the pro-UMF1; meanwhile, UFSP2 also mediate the de-UFMylation by cleaving the conjugated UFM1 from the targets (Ha et al., 2011; Kang et al., 2007). Mutations in the UFSP2 coding gene were recently reported to associate with spondyloepimetaphyseal dysplasia (Di Rocco et al., 2018; Zhang et al., 2020). We also observed that the UFSP2 was upregulated in the silica-treated pulmonary injury. Therefore, we speculate the upregulation of UFSP2 might increase the recycle of UFM1. However, the roles of the UFSP2/UFMylation in silicosis remains to be investigated.
The uncontrolled activation of pulmonary fibroblasts is the eventual progress for the collagen deposition in the lung. The EMT increased the number of activated fibroblasts in the lung in response to silica-induced inflammation (Li, 2018c). Previous studies have revealed that ER stress in the AEC participates in EMT in response to hypoxic microenvironment (Tanjore et al., 2011; Delbrel et al., 2019). Herein, we found the CADM2, an adherent molecule, was upregulated in silica-induced pulmonary injury. CADM2 was reported to regulate the EMT and associate with cancer, obesity, and energy homeostasis (Rathjen et al., 2017; Greenbaum et al., 2019; Liu, 2019b; Yan et al., 2018). The roles of CADM2 in cancers remain controversy. In most cancers, the expression of CADM2 was downregulated and plays a tumor suppressor role (Li, 2018a). However, CADM2 promotes tumor metastasis in non-small cell lung cancer by inducing EMT (Dai et al., 2020). Based on our data we speculate that the increase of CADM2 expression in silicosis models may suggest an activation of EMT.
In conclusion, we found the UFSP2/UFMylation and the CADM2 was activated in silica-induced pulmonary injury. However, the roles of them remain to be explored in the future study.
Footnotes
Disclosure Statement
The authors declare no conflicts of interest.
Funding Information
This study was supported by the Capital Clinical Application Research (Z181100001718156).