
Abstract
In late December 2019, a new kind of Coronavirus called severe acute respiratory syndrome Coronavirus 2 (SARS-CoV-2) was officially identified in Wuhan, China. In March 2020, SARS-CoV-2 was declared a pandemic by the World Health Organization (WHO), and it has infected millions of people worldwide. SARS-CoV-2 is a highly contagious Coronavirus, which has led to an outbreak of acute respiratory tract infection called “Coronavirus disease 2019” (COVID-19), resulting in mild to severe respiratory infections in humans. The design of appropriate therapeutic approaches is dependent on the understanding of molecular and cellular pathways of Coronavirus infections. In this study, we summarized the characteristic features of SARS-CoV-2. In addition, we considered the recent information regarding COVID-19 molecular immune pathogenesis, diagnosis, and potential treatment, which may provide novel perspectives and therapeutic goals in combating SARS-CoV-2.
Introduction
Over the last 20 years, numerous viral epidemics have been recognized, such as severe acute respiratory syndrome Coronavirus (SARS-CoV), from 2002 to 2003, and H1N1 in 2009 (Chen et al., 2020a). In December 2019, SARS-CoV-2 was discovered in Wuhan, China. SARS-CoV-2 is a type of RNA virus, leading to Coronavirus disease 2019 (COVID-19) (Bogoch et al., 2020). Provisionally, the 2019 novel Coronavirus was named 2019-nCoV. According to the evaluation of the virus by experts of the International Committee on Taxonomy of Viruses (ICTV), and because of its genetic similarity to that of SARS (SARS-CoVs), the World Health Organization (WHO) officially named this pathogen as SARS-CoV-2, and its disease called COVID-19 (Cascella et al., 2021).
On January 30, 2020, the WHO announced that the epidemic of SARS-CoV-2 was a public health emergency, and on March 11, 2020, it was declared as a pandemic (Neher et al., 2020; Adu-Amankwaah et al., 2021). The pandemic has rapidly spread all over China and several countries (Chen et al., 2020c; Li et al., 2020a; Lu et al., 2020). As of January 22, 2021, nearly 100 million patients have been infected with SARS-CoV-2, and more than 2 million patients have died from COVID-19 worldwide.
The rapid growth of reported cases of COVID-19 raises an urgent concern for the effective diagnosis and therapeutic strategies against SARS-CoV-2 (Huang et al., 2020). Due to this crucial circumstance, extensive understanding is necessary for the better control and treatment of the disease.
Therefore, this study aimed to throw more light on COVID-19 molecular immune pathogenesis, diagnosis, and therapies. In addition, this review will aid in making suggestions toward the control and drug discovery of SARS-CoV-2 infection by focusing on the latest developments regarding SARS-CoV-2 and the research evidence gained from studies on Middle East respiratory syndrome (MERS)-CoV and SARS-CoV.
Search Methodology
We performed systematic literature searches from the bibliographic databases, including Medline (PubMed), Web of Science, and Embase, regarding COVID-19 up to January 10, 2021. We used COVID-19, SARS-CoV-2, severe acute respiratory syndrome Coronavirus 2, Wuhan Coronavirus, 2019-nCoV, and Coronavirus disease 2019 as search terms for our survey.
SARS-CoV-2 Pathophysiology
Coronaviruses are enveloped and have a nonsegmented, positive-sense, single-strand (ss) RNA genome, varying from 26,000 to 37,000 bases, making them the largest group of RNA viruses identified to date (Weiss and Navas-Martin, 2005). COVID-19 is caused by a new Coronavirus called SARS-CoV-2 (previously named 2019-nCoV).
Coronaviruses have extremely large genomes of RNA, flanked by 5′, and 3′ UTRs included cis-acting secondary RNA structures that are essential for the synthesis of RNA. The SARS-CoV-2 genome consists of six functional open reading frames (ORFs) organized from 5′ to 3′ in order. There are two large ORFs, including ORF1a and ORF1b at the 5′’ end of Coronaviruses RNA, comprising two-thirds of the whole SARS-CoV-2 genome (Tavares et al., 2021). These two large ORFs encode nonstructural proteins, which are necessary for the preservation of the Coronavirus large genome, such as papain-like protease (PLpro), 3-chymotrypsin-like protease (3CLpro), and RNA-dependent RNA polymerase (RdRp). Other ORFs of the SARS-CoV-2 genome generally encode nonstructural proteins, structural proteins, and various accessory proteins (Li and De Clercq, 2020).
The structural proteins are composed of the spike (S) protein, the envelope (E) protein, the membrane (M) protein, and the nucleocapsid (N) protein (Fig. 1A). The structural genes at the 3′ end of the genome are interspersed with accessory genes (Fehr and Perlman, 2015).

Several studies have demonstrated that they play a significant role in the pathogenesis of Coronavirus disease (Zhao et al., 2012). The S proteins bind to host cells through a receptor called angiotensin-converting enzyme 2 (ACE2). The M and E proteins have critical functions in the assembly of the virus, and the N proteins are required for the synthesis of RNA (Song et al., 2019).
Based on whole-genome phylogenetic analysis, structural proteins of SARS-CoV-2 are more similar to SARS-CoV than to MERS structural proteins (Fig. 1B, C; Ahmed et al., 2020). Although the specific pathophysiological pathways affecting the development of SARS-CoV-2 remain unknown because of pending laboratory studies, genomic similarities with SRS-CoV may explain the incidence of severe pneumonia (Fehr and Perlman, 2015). The exact mechanism of SARS-CoV-2 continues hypothetical until such experimental studies are performed.
Molecular Mechanisms of COVID-19 in the Host Cell
The S protein of Coronavirus has been identified as a key factor for the virus entry into host cells (de Wit et al., 2016). The existence of four structural domains determines the SARS S protein. Although S1 domain and S2 ectodomains are necessary to bind the virus to the receptor and its fusion to the membrane, respectively, the cytoplasmic domains plays an important role in the assembly of various enveloped viruses (Buonvino and Melino, 2020). The receptor-binding domain of SARS-CoV-2 is on the surface-exposed S1 of the virus spike that directly binds to the receptor of the host cells, thereby determining the tropism and pathogenicity of the virus host cell. There are heptad repeat regions and the fusion peptide in the transmembrane S2 domain, leading to the viral fusion with cellular membranes through broad conformational rearrangements (Letko et al., 2020).
There are two functional subunits of Spike, including S1, binding to the receptor of the host cell, and S2, leading to the fusion of viral and cellular membranes. In the various CoVs, the S protein breaks between S1 and S2 boundary subunits, which stay noncovalently bound in the perfusion conformation (Belouzard et al., 2009). Walls et al. (2020) found that the SARS-CoV-2 entry into the host cells through S protein might be facilitated by ACE2, making it an efficient receptor for this newly emerged Coronavirus.
Zhou et al. (2020a) conducted virus infectivity experiments using HeLa cells that either expressed or did not express ACE2 protein, from humans, civet, mice, Chinese horseshoe bats, and pigs. They demonstrated that SARS-CoV-2 can enter into HeLa cells, which express all of the ACE2 proteins, excluding mice ACE2, but may not enter cells that lack ACE2, confirming that the virus needs ACE2 as its receptor for entry (Fig. 1C). Although SARS-CoV-2 uses ACE2 with identical binding affinity as SARS-CoV, its transmissibility and infectivity are entirely different. Therefore, the distinct protease-mediated S protein cleavage may cause different virus transmission and infectivity among SARS-CoV-2 and SARS-CoV (Zhou et al., 2020a).
The formation of a polybasic cleavage site (PRRAR) at the S1–S2 boundary facilitates effective cleavage by the prototype proprotein convertase furin, which is a special characteristic of the SARS-CoV-2 S protein.
Cleavage contributes to increased infection and is considered a vital process in the development of SARS-CoV-2, because the efficient S protein cleavage is essential for active infection and is a key determinant of eliminating barriers to species (Hoffmann et al., 2020a; Ou et al., 2020).
The principal host cell proteases on the cell membrane are the transmembrane protease serine 2 (TMPRSS2; Zhou et al., 2015). The SARS-CoV-2 S protein has an insertion sequence 680-SPRR-683 at the cleavage site of S1/S2 in comparison with SARS-CoV. Meng et al. (2020) demonstrated that R682, R683, and R685 were the most suitable substrates for TMPRSSs, which could improve the efficiency of TMPRSS's S protein cleavage, facilitate its activation, and enhance the infection of SARS-CoV-2. In addition to the entrance by ACE2, Wang et al. (2020) showed that S protein was bound to a specific receptor on the host cells, CD147, leading to the invasion of viruses (Wang et al., 2020). In addition to membrane fusion, SARS-CoV entrance is also facilitated through clathrin-dependent and -independent endocytosis (Wang et al., 2008). The complete SARS-CoV-2 pathogenicity mechanism in the host cell is well illustrated in Figure 2.

The SARS-CoV-2 starts its infection cycle in host cells once S protein attaches to the ACE2 receptor. Then, the cellular serine protease TMPRSS2 causes S protein priming for its activation (the S1/S2 cleavage sites of SARS-CoV-2 for TMPRSS2 includes R815 like SARS-Cov and new potential sites R682, R683, and R685). After that, the change in conformation of the S protein encourages the fusion of the SARS-CoV-2 envelope with the cell membrane through the endosomal pathway and the virus RNA genome releases into the host cell. The translated viral genome creates viral replicase polyproteins pp1a/1ab, which is then cleaved by viral proteinases, to smaller components. Also, the positive-strand RNA is encoded to a negative one, which is introduced as a template for the synthesis of viral mRNAs and then translated to specific viral proteins. Subsequently, viral RNA genome and proteins formed virions into ER and Golgi and finally transported through vesicles and released its components outside of the cell. Also, the potential therapeutic drugs for COVID-19 in the host pathways were illustrated. ACE2, angiotensin-converting enzyme 2; ER, endoplasmic reticulum; TMPRSS2, transmembrane protease serine 2
Molecular Survey on Acute Respiratory Distress Syndrome in COVID-19
Acute respiratory distress syndrome (ARDS), as a leading cause of death in COVID-19 patients, is an immunopathologic event for SARS-CoV-2 occurring in about 5% of COVID-19 patients. Short/rapid breathing and cyanosis are among the signs and symptoms of ARDS. Patients hospitalized in intensive care units require mechanical ventilators, and those who are not in a condition to breathe also entail to be linked through extracorporeal membrane oxygenation to sustain their lives (Saitz et al., 2014).
The antigen-presenting cells (APCs), which have a crucial function in the antiviral immunity of the body, can obtain the viral antigen. Unfortunately, studies on SARS-CoV-2 remain incomplete. Regarding the SARS-CoV, antigenic presentations rely primarily not only on the major histocompatibility complex class-I (MHC-I) molecules but also on the MHC class II (MHC II), which can often lead to its presentation (Liu et al., 2010).
The presence of antigen then activates the humoral and cellular immunity of the body regulated by virus-specific B and T cells (Fig. 3A). There are several studies of the cell defense of Coronavirus relative to the humoral responses. Immunologists found that the antigen-binding grove conformation along with the related antigen peptides can be recognized through the CD4+ or CD8+ T cell antigen receptors. After activation of the innate immune system and the adaptive immune system by SARS-CoV-2, a massive quantity of pro-inflammatory cytokines and chemokines is produced from immune effector cells, including interleukin-6 (IL-6), which is released through activation of the NF-κB (nuclear factor kappa-light-chain-enhancer of activated B cells) pathway.
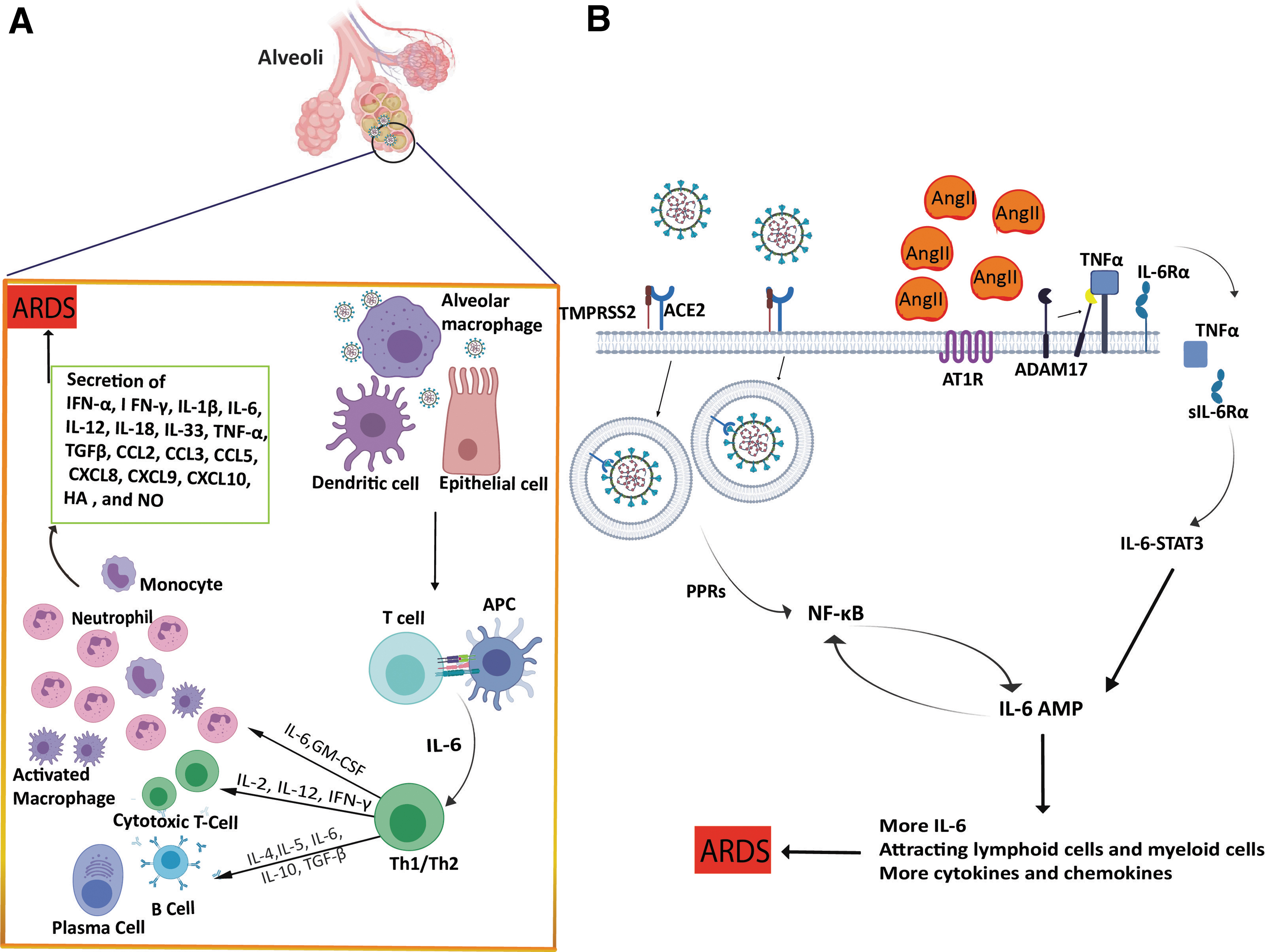
Suggested immune processes after SARS-CoV-2 infection.
Hariharan et al. (2020) found that ARDS could be induced by the hyperactivation of the NF-κB pathway. They mentioned the two different mechanisms for activation of the NF-κB pathway in response to SARS-CoV-2. In the first mechanism, SARS-CoV-2 infection activates the MyD88 pathway by pattern recognition receptors, inducing activation of several proinflammatory cytokines such as IL-6, tumor necrosis factor-alpha (TNFα), and chemokines, which can activate the NF-κB pathway. The second mechanism is dependent on ACE2 receptors.
ACE2 is endocytosed with SARS-CoV, leading to a decrease in ACE2 receptors on the cell surface, thus causing an increase in serum AngII that can bind to angiotensin receptor type 1 (AT1R). AngII-AT1R can activate NF-κB and disintegrin and metalloprotease 17 (ADAM17), raising TNFα and epidermal growth factor receptor ligands, two stimulators of NF-κB. Induction of ADAM17 changes the membrane form of the IL-6 receptor (IL-6Rα) to the soluble one and the sIL-6Rα-IL-6 complex can activate signal transducer and activator of transcription 3 (STAT3). Thus, the infection of SARS-CoV-2 can activate the IL-6 amplifier (IL-6 Amp) following trigger NF-κB and STAT3 in the respiratory system.
The IL-6 Amp produces different proinflammatory cytokines and chemokines, including IL-6, which triggers positive feedback of IL-6 signaling through attracting lymphoid cells and myeloid cells such as activated T cells and macrophages to the lesion (Fig. 3B; Hirano and Murakami, 2020).
Because of lymphocytopenia in patients with severe COVID-19, leukocytes other than T cells have to mediate for cytokine release induced by SARS-CoV-2. Chemokines and cytokines are vital producers of neutrophils. Cytokine storm may result in ARDS, which is a major cause of death in severe acute patients (Lew et al., 2003; Kim et al., 2016).
Wu et al. (2020) demonstrated that the neutrophil counts in patients with COVID-19 pneumonia were considerably greater than those in patients without ARDS. The neutrophil activation may lead to the immune response to the virus infection, contributing to the cytokine storm. This may partly explain the positive association between high fever and ARDS found at the early stages of COVID-19 (Fig. 3; Wu et al., 2020). On the other hand, the pictures of cytoplasmic tail scans showed that the typical white spots named “ground glass,” containing fluid in the lungs, are common (Bai et al., 2020). The latest studies have verified that a clear liquid jelly, the same as the wet drowning lungs, is present in lung autopsies (Xu et al., 2020b).
Although the origin of the clear jelly is not yet known, hyaluronan (HA) is related to ARDS. In addition, the regulation of HA synthesis is impaired during SARS infection (Hallgren et al., 1989).
The amounts of inflammatory cytokines like IL-1 and TNF are significantly higher in the lungs of COVID-19 patients. These kinds of cytokines are the most powerful causative agents for HA-synthase-2 in the endothelium of CD31+, lung alveolar epithelial cells, EpCAM+, and fibroblasts. It is noteworthy to mention that HA has the capacity to absorb water up to a thousand times its molecular weight. Thus, the reduction or inhibition of the HA production in the lungs of COVID-19 patients would be promising to help them to breathe.
Also, the multiple human leukocyte antigen (HLA) haplotypes are correlated with various susceptibilities to diseases. The HLA molecule repertoires composed of a haplotype can affect their longevity. Consequently, HLA molecules on the cell surface of antigen-presenting cells with improved binding specificities with SARS-CoV-2 peptides appear to be advantageous (Shi et al., 2020b).
It is also necessary to investigate whether particular HLA loci are related to the production of anti-SARS-CoV-2 immunity, and if this occurs, then classify alleles, groups I or II, indicating defense immunity induction. Simple detection kits can be established when the dominant alleles have been identified. Such findings will provide useful knowledge on the appropriate nature of SARS-CoV-2 vaccines.
Potential Treatments for COVID-19
Currently, there are no proven efficient vaccines or therapeutic agents against COVID-19, but certain therapies have demonstrated potential improvements in certain patient subpopulations or in some endpoints. Large-scale clinical studies are being performed by researchers and producers to test different treatments for COVID-19. Based on relevant literature and clinical studies, we reviewed all available data on possible treatments for SARS-CoV-2, which are presented in the following.
Entry inhibitors of SARS-CoV-2
For entry into host cells, ACE2 acts as the receptor, and human proteases act as the activator. The virus then integrates its membrane with the host cell membrane and invades it. Therefore, a possible cure for COVID-19 can be therapies interacting with host cell entry. Arbidol has been demonstrated to suppress several enveloped viruses by inhibiting the viral membranes entering/fusing with cellular membranes (Blaising et al., 2014).
In vitro studies found that it has potential activity toward SARS-CoV-2, and present clinical reports suggest that it could be more efficient than lopinavir and ritonavir in the treatment of COVID-19 (Wang et al., 2020d). The serine protease as the suppressor of camostat mesylate, accepted in Japan for treating different illnesses, has been presented to suppress the action of TMPRSS2, and thus may be an important candidate (Kawase et al., 2012). A current research found that camostat mesylate inhibits SARS-CoV-2 from entering human lung cells (Hoffmann et al., 2020b). Therefore, it could be a possible antiviral drug toward COVID-19 disease, but there is inadequate medical evidence to confirm its usefulness to date.
However, the controversial therapies that interact with SARS-CoV-2 entry are chloroquine and hydroxychloroquine (HCQ). The obvious effectiveness and appropriate protection of chloroquine phosphate, an old malaria drug, are shown in multicenter clinical trials in China against COVID-19-induced pneumonia. Data from more than 100 patients have proven that chloroquine phosphate can be advantageous for controlling pneumonia exacerbation, improving pulmonary imaging findings, encouraging a virus-negative conversion, and decreasing the duration of the disease (Gao et al., 2020).
Chloroquine and HCQ are similar in the mechanism of action and structure, except for a hydroxy moiety in one terminal at HCQ. Both of them act as weak bases that may change the pH of intracellular acidic organelles like endosomes/lysosomes, which are essential for the fusion of membrane. Both agents are considered to be useful for combating SARS-CoV-1 and SARS-CoV-2 (Colson et al., 2020). No scientific consensus has recently been obtained regarding their effectiveness in the COVID-19 therapy. Some experiments indicated that they could prevent infection of SARS-CoV-2 in vitro, but there is insufficient evidence (Wang et al., 2020c; Yao et al., 2020).
In patients treated with chloroquine or HCQ, two clinical trials were not correlated with a mortality rate in patients who did not receive the drug. On the other hand, since a greater risk of heart attack was observed in treated patients, this might raise the risk of death (Geleris et al., 2020; Rosenberg et al., 2020). The U.S. Food and Drug Administration (FDA) withdrew the emergency permission to use chloroquine and HCQ for the COVID-19 therapy on June 15, 2020, due to the adverse effects found in clinical studies (Hu et al., 2020).
Blocking of SARS-CoV-2 replication
Favilavir (T-705), remdesivir (GS-5734), ritonavir, lopinavir, and ribavirin are inhibitors of replication. All replication inhibitors target RdRp, except for ritonavir and lopinavir blocking 3CLpro (McKee et al., 2020). First, remdesivir was administered intravenously by the Washington Health Department, reporting that remdesivir could be safe for SARS-CoV-2 disease (Holshue et al., 2020). Through treating hospitalized patients with acute COVID-19, the FDA has given an emergency use permission for remdesivir. In addition, remdesivir is the first European Union-approved alternative therapy of patients with supplementary oxygen-requiring pneumonia.
The safety and effectiveness of remdesivir for the COVID-19 therapy must be assessed in various international Phase III clinical experiments (Hu et al., 2020). Hence, other analogs of nucleosides such as favipiravir, ribavirin, or galidesivir may also help protect against SARS-CoV-2 (De Clercq, 2019).
Recently, a clinical trial found that favilavir significantly decreased the duration for the clearance of SARS-CoV-2 and improved signs of COVID-19 on patients' chest imaging (Cai et al., 2020). Also, ribonucleoside analog β-
Authorized inhibitors of protease (including ritonavir, disulfiram, and lopinavir) have been identified to be effective for SARS and MERS (Servidio and Stellacci, 2021). Disulfiram, as an approved medication for alcohol abuse, has been documented to block the MERS and SARS PLpro in cell cultures, but there is a lack of clinical evidence. To find whether inhibitors of HIV protease (such as lopinavir and ritonavir) are effective for the SARS-CoV-2 identified cases, clinical trials have been initiated (e.g., ChiCTR2000029539).
Primarily, lopinavir and ritonavir were proposed to suppress SARS and MERS 3CLpro and seemed related to enhanced clinical outcomes in a nonrandomized open-label trial for SARS patients (Zumla et al., 2016). In patients with COVID-19, lopinavir and ritonavir alone had no therapeutic efficacy, but tended to be more advantageous when used in combination with other compounds, including interferon beta-1b, and ribavirin (Hung et al., 2020). It is evident that more research needs to be done to find new antiviral drugs to control COVID-19 infections. Also, there are several potential therapeutic approaches for COVID-19.
Immunomodulatory drugs
SARS-CoV-2 induces an intense immune system response that can lead to cytokine storm syndromes (Huang et al., 2020). A possible adjunctive treatment for COVID-19 can then be immunomodulatory compounds that suppress the uncontrolled inflammatory reaction. An important study revealed that dexamethasone decreased death rates approximately in a third of patients hospitalized with COVID-19, who needed invasive mechanical ventilation, and one-fifth of patients who needed oxygen. On the other hand, dexamethasone showed no effect on COVID-19 patients who did not need respiratory support (Horby et al., 2020).
The COVID-19 can be prevented by reducing unnecessary inflammatory responses as an alternative therapy. At the cellular stage, Zhou et al.(2020b) showed that CD4+T cells are quickly primed to secrete granulocyte-macrophage colony-stimulating factor (GM-CSF) and other inflammatory cytokines following SARS-CoV-2 infection. This further triggered the activation of CD14+ and CD16+ monocytes with elevated expression IL-6. Blocking the receptor of GM-CSF or IL-6 will also theoretically decrease SARS-CoV-2 immunopathology.
The effectiveness and protection of tocilizumab (an IL-6 receptor-specific antibody) in patients with COVID-19 are now being investigated in a multicenter, randomized, and controlled clinical trial (ChiCTR2000029765; Fu et al., 2020). In their study, Bian et al. (2020) demonstrated that mepolizumab effectively increased recovery for SARS-CoV-2 pneumonia patients with an appropriate safety profile (Table 1).
A List of Candidate Antiviral Drugs for Coronavirus Disease 2019
ARDS, acute respiratory distress syndrome; CAM, cell adhesion molecules; RdRP, RNA-dependent RNA polymerase; SARS-CoV-2, severe acute respiratory syndrome Coronavirus 2; TMPRSS2, transmembrane protease serine 2.
Following treatment with bevacizumab as an antivascular endothelial growth factor, pulmonary edema in patients with SARS-CoV-2 might be decreased (Rosa and Santos, 2020).
Primary studies data demonstrated that eculizumab as an anti C5 monoclonal antibody can decrease C-reactive protein (hs-CRP) and inflammatory markers levels, indicating its role in critical COVID-19 patient therapy (Diurno et al., 2020).
The expression of different interferon-stimulated genes is triggered by interferons, which may disturb any phase of replication in the virus. Previous studies have reported that type I interferons can act as a potential candidate for SARS therapy (Stockman et al., 2006). Recent experimental studies revealed that SARS-CoV-2 was much more responsive to Type I interferons than SARS-CoV, indicating the possible efficacy of type I interferons in early COVID-19 therapy (Mantlo et al., 2020). As guidance in China, interferon alpha vapor inhalation may have potential roles in the treatment of patients with COVID-19 (Sallard et al., 2020).
Treatment with immunoglobulin
In an emergency for the application of new drugs, FDA provided guidance in the expanded use of convalescent plasma for the treatment of patients with COVID-19. Nevertheless, through inducing antibody-mediated infection enhancement, transfusion-related acute lung injury, and allergic transfusion reactions, this procedure can have negative impacts.
The major advantage of monoclonal antibodies compared with convalescent plasma is that monoclonal antibodies can be produced in higher amounts to satisfy therapeutic needs. Therefore, they have the potential to prevent or treat COVID-19. In selected patients, treatment with monoclonal antibody is efficient immunotherapy for curing several viral infections (Ng et al., 2020). Recently, researchers have indicated that particular monoclonal antibodies neutralize infection with SARS-CoV-2 in vitro and in vivo (Shi et al., 2020a; Wang et al., 2020a). Relevant data for vaccine design are also given by the neutralizing epitopes of these monoclonal antibodies. However, the extensive use of monoclonal antibody therapy can be limited by high cost, the low potential of production, and the issue of bioavailability.
The Laboratory Diagnostic Methods for COVID-19
Early detection of COVID-19 is essential for the treatment and control. The testing of nucleic acid (NA) is documented to be essential to verify infections by SARS-CoV-2. Whole-genome sequencing provides one of the perfect techniques for detecting viral NAs. Five normal ORFs, including ORF1ab polyprotein, N protein, spike glycoprotein, E protein, and M protein, have been found by the first detailed SARS-CoV-2 genome sequences (Li et al., 2020b).
Following this, researchers are also uploading the GenBank with certain SARS-CoV-2 genomic sequences collected by utilizing Illumina or Nanopore platforms. Due to the expense, time consumption, and complexity of the entire genome sequence, it is not appropriate for urgent and large-scale studies (Chan et al., 2020; Chen et al., 2020b).
Examination of viral NA utilizing the reverse transcription polymerase chain reaction (RT-PCR) method can act as a significant part in the assessment of hospitalization and isolation of specific patients. Real-time RT-PCR is specific, fast, and cost-effective. Nonetheless, deficiency insensitivity, adequate stability, and reasonably long processing time have been detrimental to controlling epidemic disease (Ai et al., 2020). The reasons behind the low power of detection of viral NA may contribute to the premature expansion of viral NA detection technologies, differences in the detection rate from various manufacturers, poor viral load in patients, or incorrect clinical sampling (Fang et al., 2020). Han et al. (2020) showed that sputum induction is more effective than throat swab to classify SARS-CoV-2 RNA in recuperating patients.
Dong et al. found that digital PCR (dPCR) greatly increased accuracy and decreased false-negative SARS-CoV-2 diagnostics levels, which can be an effective complement to existing RT-quantitative PCR (qPCR) (Dong et al., 2015). dPCR is the technique that separates NA molecules over several smaller reactions, and at the end acquires amplification data from each partition at the end point dependent on fluorescence intensity (Hindson et al., 2011; Manoj, 2016). Quantification is done by utilizing Poisson statistics to the proportion of negative partitions to account for positive partitions that first included more than one target molecule. The dPCR may provide more precision than qPCR and is considerably easier to use in the quantification of copy numbers since the binary nature of the partitions is considered positive or negative. Moreover, the dPCR is more tolerant of PCR suppression in comparison with qPCR owing to partitioning and it is an endpoint PCR measurement, and subsequently, less reliant on high PCR efficacy (Strain et al., 2013).
Colorimetric loop-mediated isothermal amplification (LAMP) offers an easy and quick method for the diagnosis of SARS-CoV-2 RNA. Not only pure RNA as a sample input can be used for this method but also direct tissue or cell lysate may be used without a need for RNA purification. LAMP is a technique for the rapid amplification of NA utilizing four to six specifically designed primers and a DNA polymerase with chain displacement activity (Bst) at a steady temperature. This method can be associated with a pH indicator in the reaction mix that enables the amplification reaction to be interpreted by a shift in color. The combination of a rapid sample preparation method with an easy detection process can enable the development of portable field detection, in addition to rapid screening for point-of-need testing applications (Yu et al., 2020; Zhang et al., 2020).
Another interesting solution to RNA virus identification is the use of programmable RNA sensors. Green et al. (2014) have established sensors for the toehold to bind and detect practically any RNA sequence. To date, the researchers use a freeze-dried, paper-based, and cell-free protein expression platform, which offers a clean, abiotic manner for the storage and delivery system for these genetic circuits at room temperature, to deploy such switching sensors outside the test laboratory (Green et al., 2014).
Toehold switch sensors mostly consist of two cognate RNAs: a transducer RNA (like the mRNA of the green fluorescent protein), which encodes for the system's output signals, and the trigger RNA, which modulates the output signal. The production signal suppresses the translation by sequestrating the ribosome binding site (RBS) in a hairpin of the transducer RNA. This hairpin is unwound upon binding of a cognate trigger RNA, revealing the RBS, and does downstream protein translation (Pardee et al., 2014).
The clustered regularly interspaced short palindromic repeats (CRISPR) is a biotechnology tool renowned for its usage in editing a gene. In particular, CRISPR has become an effective and precise molecular diagnostic technique for in vitro NA detection (Gootenberg et al., 2017; Chen et al., 2018).
Among the CRISPR-Cas effector groups, Cas12 as an RNA-guided DNase is a member of class II type V-A system that triggers indiscriminate single-stranded DNA (ssDNA) collateral cleavage after the target identification. This causes the degradation of ssDNA reporters, emits a fluorescence signal on cleavage, or can be observed on a paper strip (through lateral flow) in a portable way (Myhrvold et al., 2018). Thus, CRISPR-Cas12 techniques can be used as a method to diagnose the SARS-CoV-2 quickly.
Immunoassay techniques based on the antibody coupled with a range of signal detection systems are usually considered to be effective detection methods. So far, numerous antibody discovery procedures have been accepted by the National Medical Products Administration (NMPA) for the diagnosis of SARS-CoV-2, including kits for total Ab quantity, IgM, or IgG antibodies against SARS-CoV-2, using enzyme-linked immunosorbent assay, chemiluminescence, or colloidal gold methods. Both characteristics of sensitivity and specificity are high by all the three methods for detection of total Ab and IgM, and the sensitivity for total Ab is higher compared with IgM identification (Xia et al., 2020).
Conclusions
COVID-19 disease is now a global threat to the general population. Nonetheless, there is little information about the new virus. The emergence and incidence of SARS-CoV-2 depend on how the virus interacts with the host immune system. Currently, many medications in clinical trials are tested and developed for COVID-19 therapy worldwide. All the above approaches can effectively be used in various situations for the diagnosis and treatment of COVID-19. Furthermore, the correct diagnosis of COVID-19 in the early phases tends to control the transmission of the disease. It is hoped that this study will provide some useful information regarding the battle against the deadly infectious, which is highly prevalent in the world.
Footnotes
Authors' Contributions
Z.R. contributed to prepare figures, the table, and drafted the article. F.S. was a major contributor in writing the article. L.M. edited the article. All authors read and approved the final article.
Disclosure Statement
No competing financial interests exist.
Funding Information
No funding was received for this article.