
Abstract
In response to neuronal activity changes, the adult hippocampal circuits undergo continuous synaptic remodeling, which is essential for information processing, learning, and memory encoding. Glial cells, including astrocytes and microglia, actively regulate hippocampal synaptic plasticity by coordinating the neuronal activity-induced synaptic changes at the circuit level. Emerging evidence suggests that the crosstalk between neurons and glia in the adult hippocampus is region specific and that the mechanisms controlling this process are critically dependent on secreted factors. Interleukin-33 (IL-33), a cytokine of the IL-1 family, is a key factor that modulates such glia-driven neuromodulations in two distinct hippocampal circuits. The activation of IL-33 and its receptor complex is important for maintaining the excitatory synaptic activity in the cornu ammonis 1 subregion and the remodeling of dentate gyrus synapses through activity-dependent astrocyte–synapse and microglia–synapse interactions, respectively. Meanwhile, the dysregulation of this signaling is implicated in multiple neurological disorders, especially Alzheimer's disease. Further investigations of how IL-33/ST2 signaling is regulated in a region-specific manner as well as its diverse functions in glia–synapse communications in the adult hippocampal circuitry will provide insights into the nature of hippocampal synaptic plasticity and homeostasis in health and disease.
Introduction
The hippocampus is critical for information encoding as well as memory storage and retrieval. It can be divided into the cornu ammonis 1 (CA1), CA2, CA3, and dentate gyrus (DG) subregions (Basu and Siegelbaum, 2015). Of note, the DG–CA3–CA1 synaptic circuit is essential for hippocampus-dependent memory formation in response to experience (Basu and Siegelbaum, 2015). The neurons in this circuit can alter the structure and strength of synapses in a neuronal activity-dependent manner, which is termed “synaptic plasticity”. Such activity-driven regulation of synaptic efficacy ultimately shapes the connectivity of neuronal circuits in the adult hippocampus, thereby facilitating learning and memory (Bannerman et al., 2014; Magee and Grienberger, 2020). In particular, homeostatic plasticity is a unique form of synaptic plasticity, wherein neurons adapt to offset excessive excitation or inhibition induced by chronic changes in synaptic activity, which is important for maintaining circuit homeostasis (Turrigiano, 2008; Pozo and Goda, 2010). However, improper induction and expression of hippocampal synaptic plasticity contribute to the synaptic dysfunction and memory deficits in several neurodegenerative diseases, including Alzheimer's disease (AD) (Chen et al., 2019). Thus, to better understand how the hippocampus handles information as well as its related memory functions in both physiological and pathological conditions, detailed knowledge about the synaptic plasticity in distinct hippocampal circuits is required.
Emerging evidence suggests that hippocampal synaptic plasticity is regulated not only by the bidirectional communication between presynaptic and postsynaptic neurons but also through the interactions between neurons and their enveloping glia (Allen and Lyons, 2018; Wang et al., 2021a). Astrocytes and microglia, the major types of glial cells in the adult hippocampus, can interact with synapses through their intricate processes (Perea et al., 2009; Perez-Alvarez et al., 2014; Allen and Lyons, 2018). In particular, the specialized processes of different glia express distinct receptors, which enable them to detect changes in synaptic activity in a specific circuit, thereby enabling these cells to regulate hippocampal synaptic plasticity in a subregion-specific manner (Oliet et al., 2001; Bernardinelli et al., 2014; De Pitta et al., 2016). Therefore, activation of proper glial signaling is critical for the circuit homeostasis and functioning of adult hippocampus, whereas the dysregulation of glial signaling is implicated in multiple neurodegenerative diseases (Phatnani and Maniatis, 2015; von Bernhardi et al., 2015; Wang et al., 2021a). However, it remains unclear how glia sense changes in neuronal activity and consequently affect hippocampal circuit connectivity. Nevertheless, it was recently proposed that the cytokine interleukin-33 (IL-33) is involved in such glia–neuron communication in the adult hippocampal circuitry and that IL-33-mediated signals are important for hippocampal synaptic plasticity and homeostasis (Nguyen et al., 2020; Wang et al., 2021b).
IL-33 was initially identified as an alarmin of the IL-1 family crucial for maintaining tissue homeostasis by regulating both innate and adaptive immune responses (Liew et al., 2010, 2016). Upon release, IL-33 binds to its receptor complex comprising ST2 and IL-1RAcP (IL-1 receptor accessory protein) to activate the nuclear factor-kappa B and mitogen-activated protein kinase pathways, thereby driving multiple biological processes (Liew et al., 2016). Besides the basic functions of IL-33 in immune responses, recent findings have extended the physiological roles of IL-33 to include synaptic remodeling. Specifically, IL-33 exhibits enriched expression in the neural cells in the adult hippocampus and regulates hippocampal synaptic functions in an activity- and circuit-dependent manner (Nguyen et al., 2020; Wang et al., 2021b).
Astrocyte-Secreted IL-33 Regulates Synaptic Homeostasis in the Hippocampal CA3–CA1 Circuit in an Activity-Dependent Manner
We recently identified an activity-dependent, IL-33-mediated interaction between astrocytes and neurons in the adult hippocampal CA3–CA1 circuit (Fig. 1A, B) (Wang et al., 2021b). Pharmacological blockade of neuronal activity by tetrodotoxin increased IL-33 expression and secretion in acute hippocampal slices. Of note, such activity-dependent regulation of IL-33 only occurred in a specific subpopulation of hippocampal CA1 astrocytes, suggesting that astrocyte-secreted IL-33 might mediate the activity blockade-induced hippocampal homeostatic plasticity in a circuit-specific manner. Moreover, upon optogenetic stimulation to inactivate hippocampal CA1 excitatory neurons in vivo by directly suppressing their activity or activating their neighboring parvalbumin interneurons (Klausberger and Somogyi, 2008; Ferguson and Gao, 2018), we still observed significantly increased IL-33 expression in CA1 astrocytes, suggesting that astrocyte-derived IL-33 has a unique role in the regulation of synaptic connectivity and homeostasis in the adult hippocampal CA1 circuitry.
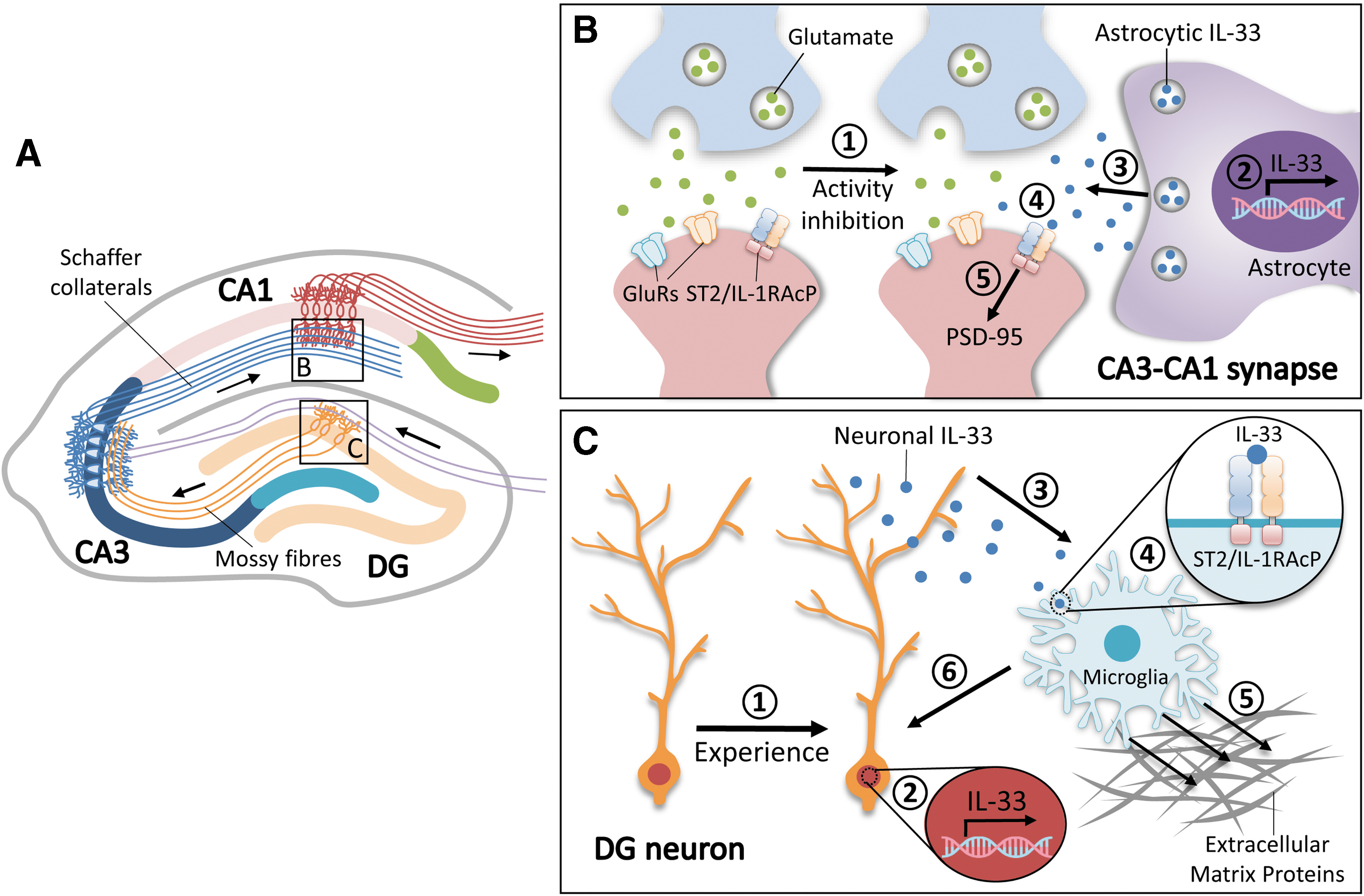
IL-33/ST2 signaling regulates hippocampal synaptic plasticity and homeostasis through circuit-dependent glia–synapse interactions.
Indeed, IL-33 administration in young adult mice promotes excitatory synapse formation and enhances synaptic transmission in the CA1 excitatory neurons. Accordingly, this synaptogenic effect of IL-33 is mediated by ST2-mediated postsynaptic mechanisms, including the phosphorylation-dependent synaptic accumulation of the postsynaptic scaffold protein PSD-95 and the subsequent strengthening of synaptic transmission. Given that chronic changes in the abundance of postsynaptic PSD-95 regulated by Ser295 phosphorylation possibly contribute to homeostatic plasticity (Kim et al., 2007), activity-induced astrocyte-secreted IL-33 might be a critical signaling regulator of this process. Meanwhile, conditional knockout of IL-33, specifically in CA1 astrocytes, decreases the number of excitatory synapses, further indicating that astrocyte-secreted IL-33 is required for the synaptic remodeling of the hippocampal CA3–CA1 circuit. These findings reveal a cellular process, whereby hippocampal CA1 astrocytes can sense and respond to cues from the CA3–CA1 circuit by the activation of IL-33 signaling. Nonetheless, it remains unclear how IL-33 expression or secretion is regulated in the astrocytes surrounding CA1 excitatory synapses or how it exerts synaptogenic effects during learning and memory. Given that diverse astrocytic ion channels and neurotransmitter receptors are responsible for the activity-dependent astrocyte–synapse interactions (Olsen et al., 2015; Allen and Lyons, 2018), further efforts to determine the specific receptors involved in this process would help explain this activity-driven IL-33 regulation in CA1 astrocytes.
Our study revealed an IL-33-driven synaptic adaptation event in the hippocampal CA1 circuit wherein activity deprivation of CA1 excitatory neurons stimulates IL-33 secretion from neighboring astrocytes; this secreted IL-33 acts on its neuronal receptor complex to regulate the homeostatic synaptic plasticity in these excitatory neurons by increasing the numbers of structural and functional excitatory synapses, thereby maintaining hippocampal network stability (Fig. 1B). Besides IL-33, studies including our RNA sequencing analysis have identified several secreted factors that are regulated by neuronal activity and involved in synaptic functions, including Chrdl1, SPARCL1, and TNFα (Stellwagen and Malenka, 2006; Kucukdereli et al., 2011; Blanco-Suarez et al., 2018; Wang et al., 2021b). Thus, IL-33 might coordinate with multiple well-known cellular signals to constitute a homeostatic feedback loop whereby CA1 neurons increase synaptic excitability, enabling them to receive more inputs from CA3 presynaptic neurons during activity inhibition; this would not only help maintain CA3–CA1 synaptic homeostasis but also facilitate a stable circuit activity in preparation for the subsequent encoding of learning experiences. As such, further examination of the activity-induced signaling cascades between astrocytes and neurons in the CA1 hippocampal circuitry will help clarify the nature of hippocampal homeostatic plasticity. Concordantly, perturbation of this IL-33-dependent homeostatic regulation by the IL-33 decoy receptor, soluble ST2, results in hippocampus-associated cognitive impairments (Wang et al., 2021b). Hence, astrocyte-secreted IL-33 is a key synaptic regulator of hippocampal homeostatic plasticity, and the homeostasis of the CA3–CA1 circuit maintained by such astrocyte–neuron IL-33/ST2 signaling is required for information processing in the adult hippocampus.
Neuron–Microglia IL-33/ST2 Signaling Is Required for Experience-Dependent Synaptic Remodeling of the Hippocampal DG Circuit
In addition to our finding that astrocyte-secreted IL-33 exhibits a region-specific effect on hippocampal homeostatic plasticity, Nguyen et al. demonstrated that in response to experience (i.e., an enriched environment), IL-33 mRNA expression is increased in a subtype of hippocampal neurons in the DG circuit; in turn, IL-33 protein binds to its ST2 receptor in nearby microglia to induce the phagocytosis of extracellular matrix proteins (Fig. 1A, C) (Nguyen et al., 2020). Given that extracellular matrix proteins can restrict the induction and expression of synaptic plasticity (Frischknecht et al., 2009), such clearance of extracellular matrix proteins during experience due to neuron–microglia IL-33/ST2 signaling is likely essential for synapse formation and plasticity in the DG, demonstrating the importance of such signaling for memory performance in young adult mice. Accordingly, dysregulated neuron–microglia IL-33/ST2 signaling in the adult hippocampal DG is implicated in aging, which is associated with memory deficits (Nguyen et al., 2020). Meanwhile, the activation of IL-33/ST2 signaling in the aging mouse brain rescues synaptic dysfunctions. Nevertheless, it remains unclear how DG neurons transduce external experience into the internal signaling cascade that includes the secretion of neuronal IL-33 and the subsequent activation of adjacent microglial phagocytosis. Given that microglia-mediated engulfment and clearance of excessive synapses (i.e., synaptic pruning) are critical for circuit development and maintenance (Paolicelli et al., 2011), it is of interest to further examine why such enriched environment-induced microglial phagocytosis only targets matrix proteins and not synapses. In addition, the DG–CA3–CA1 synaptic circuit is critical for information encoding during learning experience (Basu and Siegelbaum, 2015). Thus, determining the effects induced by enriched environments in other hippocampal subregions, including the CA3 and CA1, would help clarify the nature of such activity-dependent remodeling of hippocampal circuits.
Region-Specific Control of Adult Hippocampal Circuit Homeostasis by IL-33/ST2 Signaling
In the adult brain, IL-33 is abundantly expressed by astrocytes and oligodendrocytes (Hudson et al., 2008; Yasuoka et al., 2011). The expression patterns and levels of IL-33 differ among brain regions owing to their distinct transcriptional mechanisms (Fairlie-Clarke et al., 2018), including the expression of different enriched transcription factors (Lozzi et al., 2020) and dynamic chromatin modulations (Klemm et al., 2019). Such differential expression of IL-33 suggests that it might have distinct functions in neuronal circuits in certain hippocampal regions. Indeed, we found that the IL-33 secreted by a subpopulation of astrocytes in the hippocampal CA1 region is specifically involved in activity blockade-induced homeostatic synaptic plasticity (Fig. 1B) (Wang et al., 2021b). Meanwhile, IL-33 secreted by neurons in the DG regulates synaptic remodeling by microglial engulfment (Fig. 1C) (Nguyen et al., 2020). Thus, these findings collectively reveal a key role of IL-33 in the activity- and circuit-specific control of synaptic functions in the adult hippocampus.
Moreover, the recent emergence of novel single-cell techniques has revealed that neural cells, especially astrocytes and microglia, have highly heterogeneous genetic profiles, morphology, and functions—even within the same brain region (Mederos et al., 2018; Masuda et al., 2020). The heterogeneity of neural cells might explain their different responses to the activity-induced expression and secretion of IL-33, namely the postsynaptic responses in the CA1 subregion that regulate homeostatic plasticity as well as the microglial activation in the DG that modulates synaptic remodeling. Such distinct mechanisms elicited by IL-33/ST2 signaling enable diverse avenues for the homeostatic regulation of synaptic activity and functions in the adult hippocampal circuits in response to different learning experiences. Therefore, further single-cell transcriptomic profiling of different neural cell types in various hippocampal subregions would help reveal the cellular basis of the activity-driven neuron–glia communication in the adult hippocampal circuitry, thereby providing insights into the molecular mechanisms by which glia modulate hippocampal synaptic plasticity.
Dysregulation of IL-33/ST2 Signaling in AD
Of note, both of the abovementioned studies on IL-33-mediated glia–synapse interactions imply that dysregulated hippocampal IL-33/ST2 signaling is associated with memory deficits (Nguyen et al., 2020; Wang et al., 2021b). Concordantly, IL-33/ST2 signaling dysfunction is implicated in the progression of several neurological disorders such as AD (Chapuis et al., 2009; Allan et al., 2016; Fu et al., 2016; Liew et al., 2016). Accordingly, human genetics studies have revealed several IL-33 single-nucleotide polymorphisms related to AD (Chapuis et al., 2009). Compared to healthy controls, patients with mild cognitive impairment or AD have fewer synapses in the hippocampus (Dickerson et al., 2005; Yassa et al., 2010) and decreased IL-33/ST2 signaling in the central nervous system (Chapuis et al., 2009; Fu et al., 2016). Given that both hyperactivated microglia and astrocytes have been observed in the brains of patients with AD and transgenic mouse models of AD (Keren-Shaul et al., 2017; Fakhoury, 2018; Hemonnot et al., 2019; Habib et al., 2020), the dysregulated IL-33 signaling observed in AD is likely involved in the impaired glia–synapse interactions in the adult hippocampal circuitry. Therefore, it is of great interest to further investigate the functional roles of IL-33 signaling in glia-driven synaptic remodeling in AD. Of note, we found that replenishment of IL-33 in a transgenic mouse model of amyloid deposition (i.e., APP/PS1 mice) ameliorates synaptic plasticity deficits and cognitive impairment (Fu et al., 2016), which suggests that IL-33 indeed has a beneficial effect against AD pathology.
Conclusion and Perspectives
The findings discussed above collectively suggest the existence of a diverse IL-33/ST2 signaling-mediated biological event in the brain that extends its functions to immune cells, thereby regulating glia–neuron communication and impacting synaptic plasticity and homeostasis in the adult hippocampal circuitry. Thus, the coordination of distinct functions of IL-33/ST2 signaling in different hippocampal subregions is essential for experience-driven information processing in the adult hippocampus. Accordingly, further investigation of the molecular and cellular mechanisms of the IL-33/ST2 signaling-driven synaptic adaptations in neuronal circuits will advance our understanding of hippocampus-associated memory processes, which will also provide insights into the development of therapeutic strategies for cognitive deficits in neurological disorders.
Footnotes
Disclosure Statement
No competing financial interests exist.
Funding Information
Research conducted in the Ip Laboratory was supported, in part, by the National Key R&D Program of China (2018YFE0203600), the Research Grants Council of Hong Kong (16149616, 16102717, and 16102019; Collaborative Research Fund C6027–19GF), the Area of Excellence Scheme of the University Grants Committee (AoE/M-604/16), the Theme-based Research Scheme (T13–605/18W), the Innovation and Technology Commission (ITCPD/17–9), the Guangdong Provincial Fund for Basic and Applied Basic Research (2019B1515130004), NSFC-RGC Joint Research Scheme (32061160472), the Guangdong Provincial Key S&T Program (2018B030336001), the Shenzhen Knowledge Innovation Program (JCYJ20180507183642005 and JCYJ20200109115631248), and the Guangdong–Hong Kong–Macao Greater Bay Area Center for Brain Science and Brain-Inspired Intelligence Fund (2019001 and 2019003).