
Abstract
Herein we detected single nucleotide polymorphisms in MEF2B and UCP3 by DNA sequencing and the KASPar technology and analyzed their association with sheep growth traits. Two synonymous mutations, g.1826 C > T and g.10266 G > C, were detected, respectively, and they were found to be significantly associated with sheep growth traits (p < 0.05). In case of MEF2B g.1826 C > T, the average body weight and chest and cannon circumference of sheep with the CC genotype were significantly higher than those of sheep with the CT and TT genotypes (p < 0.05). Moreover, in case of UCP3 g.10266 G > C, the average body weight and chest and cannon circumference of sheep with the GG genotype were significantly higher than those of sheep with the GC and CC genotypes (p < 0.05). Moreover, the average body weight of sheep with the CC/GG genotype was higher compared with those of other genotype combinations. We also assessed MEF2B and UCP3 expression in different sheep tissues, confirming their expression in all examined tissues. To summarize, we believe that the polymorphisms identified in MEF2B and UCP3 can serve as molecular markers for sheep growth traits.
Introduction
Sheep, an important economic livestock species, provides meat and fur. Sheep meat (i.e., mutton) has a high nutritional value and low cholesterol content, making it highly popular (Montossi et al., 2013). Growth traits play a vital role in mutton production. In sheep, these are complex and defined by, for example, body weight, body size, and average daily gain. Furthermore, genetic factors, feeding conditions, and environmental factors influence growth traits.
In traditional breeding methods, performance measurement involves a substantial investment of manpower and resources and is also associated with errors as measurements are manually obtained. Marker-assisted selection is one of the most widely used new molecular technologies. It is reported that it can significantly reduce breeding years and improve the yield of high-quality livestock(Georges, 1999; Chen et al., 2019). Therefore, identifying genes or quantitative trait loci seems to be an effective method to improve sheep growth traits as well as production performance (Duffy, 2008; Bolormaa et al., 2017).
MEF2B belongs to the myocyte enhancer family (MEF2A, MEF2B, MEF2C, and MEF2D) (Zhang et al., 2016) and the MADS [MCM1, Agamous, Deficiens, and serum-response factor] box family of transcription factors (Martin et al., 1993). It mainly affects muscle growth and development and also evidently plays a key role in myocyte differentiation. It is located on chromosome 5 and shows some expression in various tissues (Firulli et al., 1996; Iida et al., 2005). Lin et al. reported that MEF2B expression was significantly upregulated on knocking out MEF2C, indicating that MEF2B and MEF2C have overlapping functions (Lin et al., 1997).
Furthermore, Morisaki et al. (1997) found that MEF2B was expressed at low levels in the cardiac and skeletal muscles of adult mice but at high levels in those of humans; MEF2B accordingly has been reported to be the most divergent member, showing low homology in different species (Morisaki et al.). MEF2B mutations are seemingly associated with CREBBP and EP300 mutations, as well as EZH2 Y641 mutation (Morin et al., 2010). Moreover, a recent study based on diffuse large B cell lymphoma found MEF2B expression to be positively correlated with BCL6 and CD10 expression (Salama et al., 2020).
Uncoupling proteins (UCPs), which are mitochondrial inner membrane carrier proteins, are widely distributed in various animal tissues. They play a pivotal role in promoting body heat production, maintaining normal metabolism, regulating body weight, and maintaining body temperature. The UCP family includes five members: UCP1, UCP2, UCP3, UCP4, and UCP5/BMCP1 (Pohl et al., 2019). UCP3 mainly exists in the skeletal muscle and adipose tissue, and is closely related to exercise. UCP3 and UCP2 exist on chromosome 15 as gene clusters (Stone et al., 2015), and they are key potential targets for the treatment of aging, degenerative diseases, diabetes, and perhaps even obesity (Brand and Esteves, 2005).
The role of UCP3 has been widely studied in humans and rodents; however, in the field of livestock and poultry, most studies have focused on pigs and birds, and only a few on sheep.
Raimbault et al. reported that avian UCP mRNA expression was restricted to the skeletal muscle and its abundance was increased in a chicken line with diet-induced thermogenesis; their results implied that avian UCP plays a role in avian energy expenditure(Raimbault et al., 2001). In addition, Cieslak et al. observed that the g.946 C > T missense mutation in UCP3 was significantly associated with backfat thickness in two breeds of pigs (Cieslak et al., 2009).
In the present study, we aimed to determine the relationship between single nucleotide polymorphisms (SNPs) in MEF2B and UCP3 and growth traits of sheep, as well as their expression levels in different sheep tissues.
Materials and Methods
Ethics statement
The study was carried out as per animal care and experiment procedures in accordance with the regulations and guidelines of the Government of Gansu People's Congress. The program has been approved by the Animal Conservation and Ethics Committee of Gansu Agricultural University (License No. 2012-2-159). Institutional review board approval of our hospital was obtained for this study.
Animals and DNA extraction
Lambs (1160 male Hu and 82 male Dorper lambs) were procured from four commercial sheep farms (Jinchang Zhongtian Sheep Industry Co. Ltd., Gansu Sanyang Jinyuan Husbandry Co. Ltd., Gansu Zhongsheng Huamei Sheep Industry Development Co. Ltd., and Wuwei Pukang Breeding Co. Ltd. in Gansu Province, China). At 56 days of age, they were immunized by following standardized procedures before weaning. All lambs were kept indoors in separate enclosures (0.8 × 1 m) until they were 180 days old.
In short, all lambs were exposed to a period of adaptation of 14 days; during this period, the dietary proportion of pellet feed (Gansu Sanyang Jinyuan Husbandry Co. Ltd.) was gradually increased by 7.1% every day and the forage proportion was simultaneously decreased until the pellet feed proportion became 100%. The experimental period was 100 days and the pre-experimental period was 10 days. All animals had ad libitum access to water and pellet feed. They were weighed using a calibrated electronic scale before feeding in the morning. Birth weight, weaning weight, and growth traits (80, 100, 120, 140, 160, and 180 days) were recorded. The measured growth traits included body weight, body height, body length, chest circumference, and cannon circumference.
The feeding methods and the environment were kept consistent at all times during the experiment. At the end of the experimental stage, 5 mL venous jugular blood samples were obtained from each animal, and DNA was isolated using a kit (TransGen Biotech, Beijing, China). The extracted DNA was then dissolved in an elution buffer (10 mM Tris hydrochloride and 1 mM ethylenediaminetetraacetic acid, pH 8.0). DNA samples were quantified through spectrophotometry and 1.5% agarose gel electrophoresis was performed to confirm their integrity. They were subsequently stored at −20°C until needed.
Phenotypic data of the 1160 Hu sheep were collected across five different batches: 188 in the first batch, 236 in the second, 188 in the third, 357 in the fourth, and 191 in the fifth. Phenotypic data of the 82 Dorper sheep were simultaneously collected at the same time as the fourth batch of the Hu sheep.
SNP recognition and genotyping
Specific PCR primers for MEF2B and UCP3 (GenBank accession no. NC_040256.1 and NC_040266.1, respectively) were designed (Oligo 7). Mixed sheep DNA was used as the template for PCR, and SNPs in MEF2B and UCP3 were identified in the resultant amplicons. For sequencing, PCR was performed in a 35 μL reaction volume comprising 2 × TSINGKE Master Mix (TSINGKE Biological Technology, Beijing, China), 10 μM forward and reverse primers each, and 50 ng template DNA. The cycling conditions were as follows: 3 min at 94°C; followed by 30 s at 94°C, 30 s at 45°C–65°C, and 30 s at 72°C for 35 cycles; and finally, 5 min at 72°C.
For genotyping the SNPs found in MEF2B and UCP3, genotype-specific primers were used for competitive PCR (KASPar), as reported by a previous study (Zhang et al., 2020). In the entire experimental group, 1146 and 1155 sheep with MEF2B and UCP3 were successfully genotyped.
Tissue samples and expression analysis
The animals were slaughtered at 180 days of age; tissue samples were subsequently obtained (the heart, liver, spleen, lung, kidney, rumen, duodenum, muscle, lymph, and tail fat) and temporarily stored in liquid nitrogen, before being stored at −70°C until needed.
In the experimental population, six animals were randomly selected. RNA was extracted from each tissue using TransZol Up and chloroform. RNA integrity was detected by agarose gel electrophoresis and concentration by near-infrared spectroscopy. cDNA was then obtained using a Retrotranscription Kit (TransGen Biotech). Specific primers and cDNA extracted from different tissues were used for fluorescent quantitative PCR. mRNA expression levels of MEF2B and UCP3 were analyzed by real-time fluorescence quantitative PCR; β-actin served as the internal control. Four technical replicates were assessed for each sample. Data were analyzed using the 2−ΔΔCt method, and average relative expression levels of genes were calculated.
Statistical analyses
SPSS 23.0 was used for statistical analyses. The general linear model was applied: the phenotypic data of the experimental population corresponding to the genotype served as the dependent variable, and the genotype, batch effect, and variety effect were the fixed factors. The correlation between the genotypic and phenotypic data was analyzed with Tukey's and Duncan's tests.
Results
Descriptive statistics and correlation analysis of growth traits
Table 1 shows the descriptive statistics of body weight, body height, body length, chest circumference, and cannon circumference for experimental sheep at each stage during 80–180 days. All growth traits showed an increase with age. The correlation analyses of growth traits (Fig. 1) showed that the correlation between chest circumference and body weight was the highest, whereas body height showed a weak correlation with the other growth traits. Nevertheless, a positive correlation was observed among all growth traits.

Correlation analysis performed using body weight (BW), body height (Height), body length (Length), chest circumference (Chest), and cannon circumference (Cannon) of sheep.
Descriptive Statistics of Growth Traits
SNPs in MEF2B and UCP3
Table 2 shows the primers and PCR conditions used to amplify a 303-bp fragment of MEF2B and 487-bp fragment of UCP3 (Fig. 2). On sequencing the amplicons, two synonymous mutations were found in MEF2B and UCP3: g.1826 C > T and g.10266 G > C, respectively. The SNP g.1826 C > T in MEF2B was located in the first intron, and the SNP g.10266 G > C in UCP3 was located in the seventh exon (Fig. 3). The two SNPs were genotyped using the KASPar technology (Table 3), and three genotypes were consequently identified: CC, TT, and CT and GG, CC, and GC, respectively (Fig. 4).

PCR amplification of sheep MEF2B
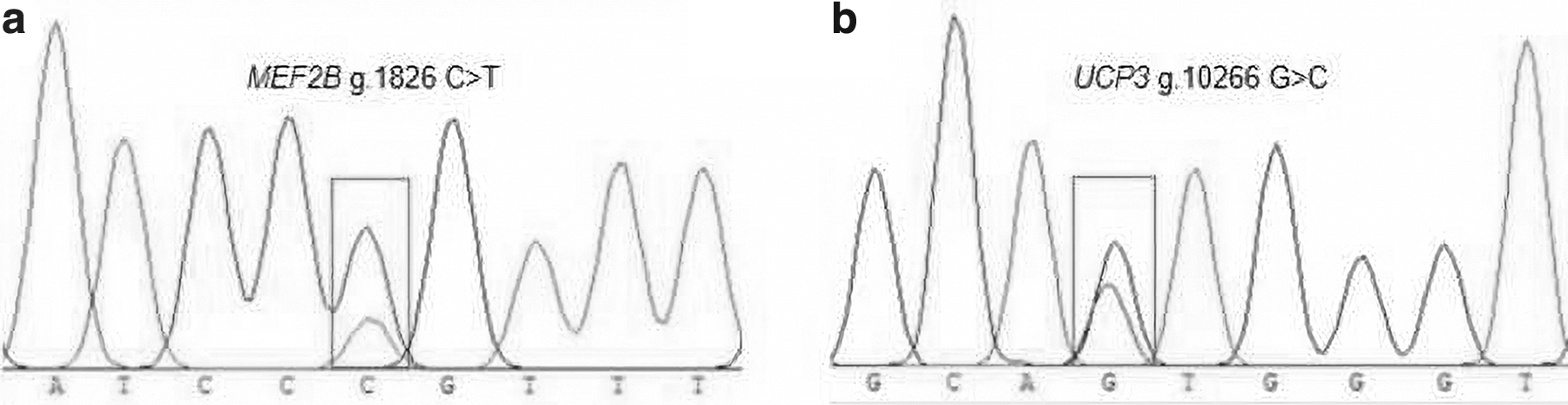
Sequencing peak map of MEF2B
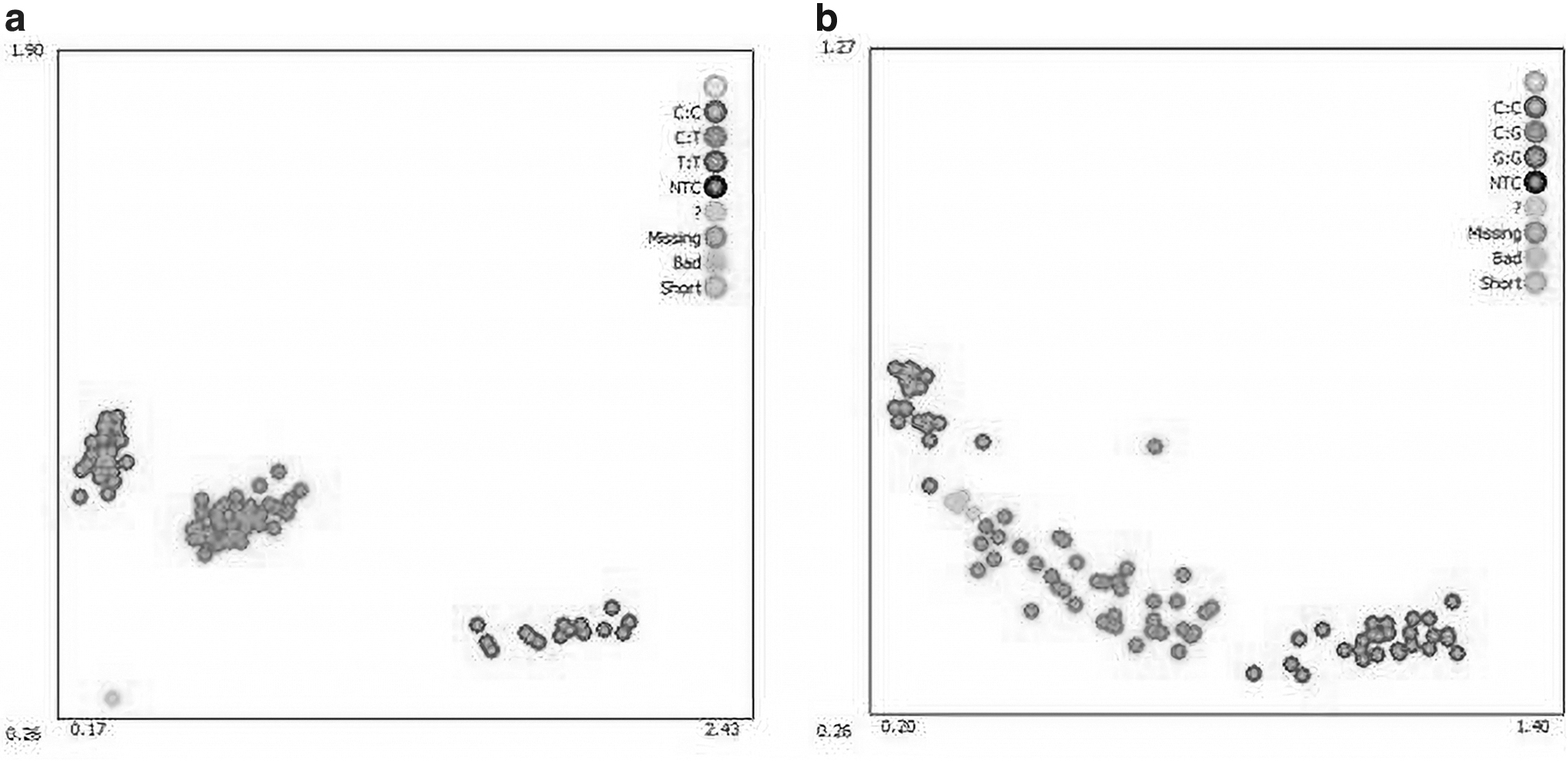
Genotyping map of MEF2B
Primer Sequences and PCR Conditions
Primers Used for the KASPar Technology
Genetic parameters of the identified SNPs
Table 4 shows the genetic parameters of the SNP g.1826 C > T in MEF2B and g.10266 G > C in UCP3. For MEF2B g.1826 C > T, the genotype frequencies of CC, CT, and TT were 0.58, 0.37, and 0.05, respectively. CC was identified to be the dominant genotype, and C was the dominant allele (0.76). Furthermore, for UCP3 g.10266 G > C, the genotype frequencies of GG, GC, and CC were 0.29, 0.39, and 0.32, respectively. C was the major allele (0.51). The He, Ho, Ne, and PIC for MEF2B were 0.37, 0.63, 1.59, and 0.30, respectively, and those for UCP3 were 0.50, 0.50, 2.00, and 0.38, respectively, showing moderate polymorphism for both.
Genetic Parameters of SNPs at the MEF2B and UCP3 Loci
SNP, single nucleotide polymorphism.
Association between SNPs in MEF2B and UCP3 and growth traits
To explore the association between SNPs in MEF2B and UCP3 and growth traits, we used the general linear model (Table 5). MEF2B g.1826 C > T was significantly correlated with body weight and chest circumference at 80, 100, 120, 140, 160, and 180 days and with cannon circumference at 80, 100, 120, 140, and 180 days (p < 0.05). In addition, the body size of sheep with the CC genotype was higher than that for sheep with the CT and TT genotypes, indicating that CC was the dominant genotype related to sheep growth traits.
Correlation Between Growth Traits and SNPs in MEF2B and UCP3
Statistical data are expressed as mean ± standard deviation. p < 0.05 indicates statistical significance.
Furthermore, UCP3 g.10266 G > C was significantly correlated with body weight, chest circumference, and cannon circumference at 80, 100, 120, 140, 160, and 180 days (p < 0.05). The body size of sheep with the GG genotype was higher than that for sheep with the CC and GC genotypes, indicating that GG was the dominant genotype related to sheep growth traits.
Association between MEF2B and UCP3 genotype combinations and sheep body weight
As shown in Table 6, we analyzed the complete effects of the two SNPs on sheep body weight. At 80, 100, 120, 140, 160, and 180 days, the body weight of animals with the CC MEF2B /GG UCP3 genotype was higher compared with those with other genotypes; moreover, at 80, 120, 140, 160, and 180 days, the body weight of animals with the CC MEF2B /GG UCP3 genotype was significantly higher compared with those with the CT MEF2B /GG UCP3 , CT MEF2B /GC UCP3 , CT MEF2B /CC UCP3 , and TT MEF2B /GG UCP3 genotypes (p < 0.05).
Association Between MEF2B and UCP3 Genotypes and Sheep Body Weight
Statistical data are expressed as mean ± standard deviation. p < 0.05 indicates statistical significance.
MEF2B and UCP3 expression levels in different tissues
The cDNA of MEF2B, UCP3, and β-actin was amplified by qRT-PCR. Gel electrophoresis revealed that the size of the amplified products was as anticipated.
Real-time quantitative PCR, which was performed to detect MEF2B and UCP3 expression levels in the heart, liver, spleen, lung, kidney, rumen, duodenum, muscle, lymph, and tail fat of six experimental sheep, revealed that MEF2B and UCP3 were expressed in the aforementioned tissues (Fig. 5). The relative expression level of MEF2B was higher in the lung and kidney than in the other tissues and that of UCP3 was high in the lung but low in the remaining tissues.
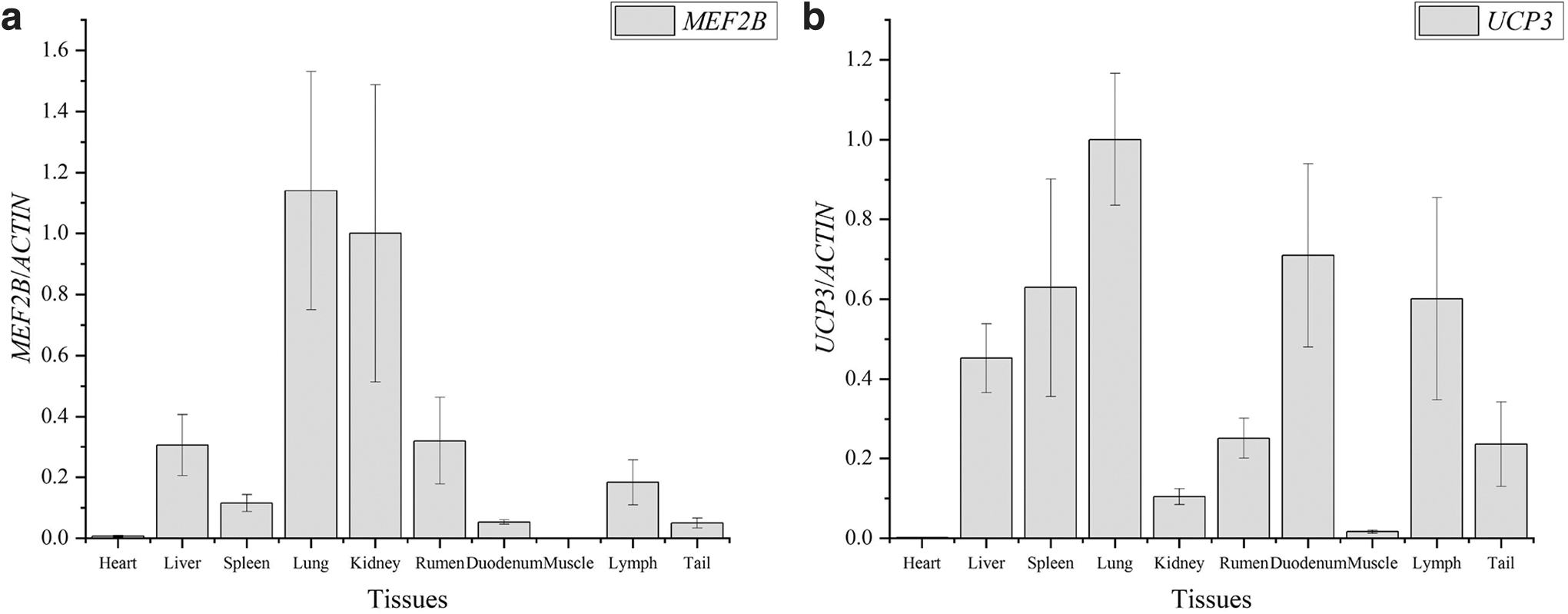
Relative expression levels of MEF2B
Discussion
Breeding efficiency can be substantially improved by identifying SNPs in genes, consequently leading to an improvement in the production performance of sheep; missense mutations in coding exons are the main target. However, progressively more evidence reveals that synonymous mutations also influence gene functional expression profiles.
In this study, we investigated the association between SNPs in MEF2B and UCP3 and sheep growth traits. Two synonymous mutations were found in MEF2B and UCP3 in total. In previous studies, synonymous mutations have been reported to affect gene functional expression by changing the protein folding mechanism, affecting mRNA splicing as well as RNA stability and folding (Zhu et al., 2018; Synonymous codon substitutions perturb cotranslational protein folding in vivo and impair cell fitness, 2020; Sharma et al., 2019 ). Therefore, we herein speculated that the two synonymous mutations detected in MEF2B and UCP3 affect sheep growth traits by affecting the transcription and translation of genes that regulate these traits.
MEF2B chiefly exists in muscle and nerve tissues. MEF2B and other genes belonging to the same family play a key role in myocyte development and also participate in regulating various gene functions besides muscle cell differentiation (Xiao-Meng et al., 2015). At present, the research on MEF2B mainly focuses on its effects on lymphoma. For example, Liu et al. reported that MEF2B expression was related to the pathological and structural subtypes of mantle cell lymphoma (Liu et al., 2020), and Julia et al. (2015) found that MEF2B mutations in non-Hodgkin lymphoma dysregulated cell migration by reducing MEF2B target gene activation.
Herein, to study the effects of MEF2B on sheep growth traits, we analyzed the correlation between different genotypes and these traits. Our findings revealed that MEF2B g.1826 C > T was significantly correlated with sheep growth traits at different stages of age (p < 0.05). Molkentin et al. showed that MEF2B binds to specific sites of muscle development-related genes (Molkentin and Olson, 1996), implying that in the absence of MEF2B or related family members, the expression of muscle development-related genes is partially interrupted. Hence, we believe that mutations in MEF2B can affect muscle development and growth traits.
UCP3 is mainly distributed in the inner membrane of mitochondria and mediates the uncoupling of oxidation and ADP phosphorylation processes. The main physiological role of UCP3 is to regulate fatty acid metabolism (Harper et al., 2001). The current research on UCP3 mainly focuses on cattle and swine. For example, Sherman et al. found A/G mutation in the third intron of UCP3 in cattle and reported that it was significantly associated with average daily gain (Sherman et al., 2008). In addition, Fabris et al. suggested that the in vivo gene expression of UCP2 and UCP3 in the skeletal muscle of rats is differently regulated by circulating lipids and that selective insulin resistance seems to somewhat favor the prevention of fat accumulation in tissues not primarily destined for fat storage, contributing to increased adiposity (Fabris et al., 2001).
Cristiana et al. reported that UCP3 expression was positively correlated with carbohydrate oxidation and negatively correlated with lipid oxidation (Cristiana et al., 2017), and Zhang et al. found a significant association between UCP3 gene polymorphisms and growth traits at different growth stages in Nanyang cattle (Zhang and Li, 2011). In this study, on assessing the effects of an SNP in UCP3 on sheep growth traits, we found that UCP3 g.10266 G > C was significantly correlated with body weight, chest circumference, and cannon circumference at each stage of growth (p < 0.05). This effect was consistent with that of UCP3 on the growth traits of cattle.
Conclusions
On analyzing the combined effects of MEF2B g.1826 C > T and UCP3 g.10266 G > C on sheep growth traits, we observed that the growth traits of sheep with the CC MEF2B /GG UCP3 genotype were much better than those of sheep with other genotypes, with body weight showing a significant difference (p < 0.05). Correlation analyses revealed that there was a positive correlation between body weight and other growth traits. To summarize, our data indicated that g.1826 C > T and g.10266 G > C in MEF2B and UCP3, respectively, affected the growth traits of sheep, suggesting that these SNPs can serve as candidate molecular markers for sheep growth traits. Further studies are nevertheless warranted.
Footnotes
Disclosure Statement
No competing financial interests exist.
Funding Information
This work was supported by the National Natural Science Foundation of China (31960653), the National for Joint Research on Improved Breeds of Livestock and Poultry (19200140), and the Gansu Province Modern Silk Road Cold and Arid Agriculture Special.