
Abstract
Breast cancer (BC) is one of the most frequently occurring malignant tumors in female adults. The long intergenic nonprotein coding RNA 00982 (LINC00982) has been regarded as a cancer suppressor in several human cancers. However, the function and the underlying mechanisms of LINC00982 have not been studied in BC. The present study found that LINC00982 was significantly downregulated in BC tumor tissues, and the low LINC00982 level predicts a poor prognosis of BC. Through the overexpression and suppression of LINC00982 in two BC cell lines, we found that LINC00982 could inhibit cell proliferation, migration, and invasion by suppressing the activity of the signal transducer and activator of transcription 3 (STAT3)/nuclear factor kappa B (NF-κB) signal pathway. Furthermore, luciferase reporter assay has been used to verify that LINC00982 functions as a molecular sponge for miR-765, which could target DPF3. The relative expression of miR-765 decreased with LINC00982 overexpressing, and DPF3 increased at the same time.
In addition, the suppression of cell malignant phenotype caused by overexpression of LINC00982 can be reversed by inhibition of DPF3. To verify the function of LINC00982 in vivo, the BC cells were implanted in nude mice and the results suggested the tumor growth and malignant phenotype were suppressed by LINC00982. In this study, we prove that LINC00982 regulates the growth and development of BC through STAT3/NF-κB signal pathway, mediated by the miR-765/DPF3 axis. LINC00982 may function as a target molecule to take part in the prognosis and therapy of BC.
Introduction
Breast cancer (BC) is one of the most common malignancies (Siegel et al., 2018), which is still a major cause of cancer-related death among women worldwide (Lu et al., 2009). It accounts for nearly one-third of women diagnosed with cancer in the United States (DeSantis et al., 2011). Despite significant progress in detection and treatment, cancer metastasis remains the primary cause of death in BC patients. About 90% of cancer-related deaths are caused by tumor metastasis (Network, 2012). Currently, the 5-year survival rate in patients with BC metastasis is less than 30% (Peart, 2017). Compared with distant metastatic BC patients, the 5-year overall survival rate of patients without metastasis has risen to 55% approximately (Valastyan and Weinberg, 2011; Allemani et al., 2018).
BC presents a tendency to distinct organs, which leads to terrible consequences to treatment and patient prognosis. Therefore, it is crucial to explore the potential molecular mechanisms underlying BC pathology in finding novel therapeutic targets.
In recent years, accumulating evidence confirmed that long noncoding RNAs (lncRNAs) played crucial roles in different kinds of cancer. According to the human genome program, the genes that encode proteins account for less than 2% of the whole genome, whereas many of the remaining sequences without coding proteins still can transcribe, and the transcript is noncoding RNA (Djebali et al., 2012; Lee, 2012).
lncRNA is a class of transcribed RNA molecules greater than 200 nucleotides in length and lacks protein-coding potential (Renganathan and Felley-Bosco, 2017). According to numerous studies, lncRNAs have been found to tightly correlate with various biological processes, including cell proliferation, migration, and invasion (Li et al., 2016; Ransohoff et al., 2018). The long intergenic nonprotein coding RNA 00982 (LINC00982) plays an important role in various human cancers. Recently, there are some researchers who demonstrated that LINC00982 promotes the expression of cathepsin F (CTSF) by blocking the interaction between transcription factor YRPW motif 1 (HEY1) and CTSF promoter, and this lncRNA acts as a tumor suppressor in gastric cancer and prevents the development of gastric cancer cells (Zheng et al., 2021).
In 2019, it was found that LINC00982 downregulates in renal cancer samples in contrast with the adjacent specimens, and the upregulation of LINC00982 suppresses cell proliferation and potentiated cell apoptosis by affecting the activity of phosphatidylinositol 3-kinase (PI3K)/AKT signaling pathway in renal cancer (Zhang et al., 2019). In addition, it was found that the low expression of LINC00982 and PR domain-containing 16 (PRDM16) has a close relation with poor prognosis in lung adenocarcinoma (Lv et al., 2018). Based on these researches, LINC00982 is assumed to be a tumor suppressor in different human cancers. However, the function and the underlying mechanisms of LINC00982 in the development of BC have not been fully explored.
In the present study, we aimed to explore the functions and clinical significance of LINC00982 in BC. We discovered that LINC00982 expression was decreased in tumor tissue, and this lncRNA could inhibit the proliferation, migration, and invasion in BC by suppressing the activity of the signal transducer and activator of transcription 3 (STAT3)/NF-κB signal pathway. Moreover, LINC00982 was confirmed to be a molecular sponge of miR-765, which targets DPF3 and affected the expression of miR-765 and DPF3 at the same time. Our findings clarified that LINC00982 played a significant role to suppress the development of BC cells and could serve as a potential therapeutic target for cancer prognosis and therapy.
Materials and Methods
Bioinformatic analysis
The expression of LINC00982 in BC tumor and normal tissues, and its expression levels at different stages of BC from the Cancer Genome Atlas (TCGA) database was analyzed by the Gene Expression Profiling Interactive Analysis (GEPIA) website (
Tissue collection
The 40 samples of BC and adjacent tissues were collected from Shengjing Hospital of China Medical University. The patients, who provided samples for this study, have known the application of these issues. All patients signed the written informed consent. This research was approved by the Ethics Committee of Shengjing Hospital of China Medical University (2017PS308K).
Cell culture and transfection
BC cell lines (MDA-MB-231, MCF-7, MDA-MB-468, and MDA-MB-453) and human normal mammary cell line (MCF-10A) were purchased from iCell Bioscience Inc., (Shanghai, China). BC cell line T-47D was purchased from Procell Life Science & Technology Co., Ltd. (Wuhan, China). MDA-MB-231, MDA-MB-468, and MDA-MB-453 cells were cultured in Leibovitz's L-15 Medium (Procell, Wuhan, China) containing 10% fetal bovine serum (FBS) (Sijiqing, Hangzhou, China). MCF-7 cells were grown in Minimum Essential Medium (Solarbio, Beijing, China) containing 10% FBS. T-47D cells were grown in Roswell Park Memorial Institute (RPMI)-1640 medium (Solarbio, Beijing, China) containing 10% FBS. MCF-10A is cultured with the dedicated medium. All of these cells were cultured in an incubator at 37°C, 5% CO2.
We constructed LINC00982 overexpressed vector and knockdown vector using adenovirus, and sh-DPF3 and sh-NC were constructed in adenovirus. miR-765 mimic, NC-mimic, miR-765 inhibitor, and NC-inhibitor were purchased from JTS scientific company (Wuhan, China). Cell transfection was performed by the Lipofectamine 3000 Kit (Invitrogen, CA).
RNA extraction and quantitative real-time PCR analysis
Total RNAs from tissue samples or cultured cells were extracted using TRIpure Reagent (Bioteke, Beijing, China). miRNAs were isolated using the Total RNA Extraction Kit (Tiangen Biotech, Beijing, China). Both of them were quantified using an ultraviolet spectrophotometer NANO 2000 (Thermo Scientific, PA). Then, total RNAs were used for reverse transcription reaction and cDNA synthesis using BeyoRT II M-MLV reverse transcriptase (Beyotime Biotech, Shanghai, China).
The miRNAs were reversed by using the miRNA First-Chain cDNA Synthesis (plus tail method) Kit (Sangon Biotech, Shanghai, China). SYBR Green Real-time Master Mix (Solarbio, Beijing, China) and 2 × Taq PCR Master Mix (Solarbio, Beijing, China) were used for quantitative real-time PCR (RT-qPCR). All primers were synthesized from GenScript Biotechnology (Nanjing, China) and presented in Table 1. Fluorescent quantizer Exicycler™ 96 (Bioneer, Daejeon, Korea) was used for RT-qPCR and data collection. The relative mRNA expression was analyzed by using the 2−ΔΔCT method.
The Primer Sequences of Different Genes in Quantitative Real-Time PCR
Western blot analysis
Total proteins were isolated by using RIPA buffer (Beyotime Biotech, Shanghai, China) and their concentration was detected by using the BCA Kit (Beyotime Biotech, Shanghai, China). Proteins were separated by 10% sodium dodecyl sulfate–polyacrylamide gel electrophoresis (SDS-PAGE) and then transferred to polyvinylidene fluoride membranes (Millipore, MA).
Subsequently, the membrane was incubated with primary antibodies, including anti-cyclin D1 (A19038, ABclonal), anti-CDK4 (A0366, ABclonal), anti-MMP-2 (10373-2-AP, Proteintech), anti-Vimentin (A19607, ABclonal), anti-STAT3 (AF6294, Affinity), anti-p-STAT3 (AF3293, Affinity), anti-p65 (AF5006, Affinity), anti-p-p65 (AF2006, Affinity), anti-DPF3 (bs-13866R, Bioss), and anti-GAPDH (AC-002, ABclonal) at 4°C overnight. The dilution ratio of these primary antibodies is 1:1000. Next, the membranes were incubated with goat anti-mouse IgG-HRP (1:5000, A0216, Beyotime) or goat anti-rabbit IgG-HRP (1:5000, A0208, Beyotime) at 37°C for 45 min.
Finally, the ECL (Beyotime Biotech, Shanghai, China) was utilized to detect the protein expression levels. The original blot images are shown in Supplementary Data.
MTT assay
MDA-MB-231 or MCF-7 cells (3 × 103 per well) were plated in 96-well plates and cultured for 12, 24, 36, 48, 60, and 72 h, respectively. At the indicated time, cells in each well were treated with 20 μL MTT reagent (KeyGen Biotech., Nanjing, China) for 4 h in an incubator. After the culture medium was removed, 150 μL of dimethyl sulfoxide was added into each well, and absorbance was evaluated at 490 nm.
Flow cytometry
Cell cycle distribution was examined using the Cell Cycle Detection Kit (Beyotime Biotech, Shanghai, China) following the instruction. For cell cycle distribution, the infected MDA-MB-231 and MCF-7 cells were stained with Propidium Iodide solution for 30 min and then detected by a NovoCyte flow cytometer (ACEA Biosciences, CA).
Immunofluorescence staining
After cell infection and culture, cell samples were fixed with 4% paraformaldehyde for 15 min, and after 0.1% Triton X-100 infiltration for 30 min, goat serum (Solarbio, Beijing, China) was utilized for the blocking step at an ambient temperature for 15 min. Next, the cells were incubated with E-cadherin antibody (1:100, A3044, ABclonal) and p65 antibody (1:100, AF5006, Affinity) at 4°C overnight, followed by incubation with FITC-marked goat anti-rabbit IgG (1:200, A0562, Beyotime) and Cy3-marked goat anti-rabbit IgG (1:200, A0516, Beyotime) at room temperature in the dark for 1 h, separately. Then samples were stained with DAPI (Aladdin, Shanghai, China) to redye the nucleus. Finally, Olympus confocal microscope (Olympus, Tokyo, Japan) was used to observe and photograph the results.
Luciferase reporter assay
The fragments of LINC00982 and DPF3 3′UTR wild type (WT) or mutant-type (MT) containing the binding sites of miR-765 were cloned into pmirGLO vectors. The constructed vectors were cotransfected into HEK-293T cells (Zhong Qiao Xin Zhou Biotechnology Co., Ltd., Shanghai, China) along with miR-765 mimic or NC mimic through Lipofectamine 3000 (Invitrogen, CA). Forty-eight hours after transfection, the luciferase activity was detected by running a multifunctional microplate reader (TECAN, Männedorf, Switzerland) using the Luciferase Assay Kit (KeyGEN, Nanjing, China).
Transwell migration and invasion assay
For transwell migration assay, MDA-MB-231 and MCF-7 cells were planted in the upper transwell chamber (Corning, NY) in a 37°C incubator. For invasion assay, cells were planted in the Matrigel-covered upper chamber. Meanwhile, the upper chamber and the lower chamber were both added with a corresponding medium containing 10% FBS. After incubation for 24 h, the migrated and invaded cells were fixed with 4% polyformaldehyde for 20 min and stained with 0.5% Crystal Violet for 5 min. Finally, the migrated or invaded cells were observed and counted under a microscope.
Immunohistochemistry
Isolated tumor tissues were fixed and dehydrated, followed by paraffin embedding, and serial sectioning. After drying, a paraffin-embedded section (5 μm in thickness) was dewaxed, rehydrated, and incubated with hydrogen peroxide. After blocking with goat serum, sections were incubated with DPF3 antibody (1:100, Bioss, Beijing, China) at 4°C overnight. Then, was incubated in HRP-marked goat anti-rabbit IgG (1:500; Thermo Scientific, PA) for 1 h at 37°C. After DAB coloration, Hematoxylin redyes, and dehydration, sections were visualized using a microscope.
In vivo tumorigenesis assay
A total of 24 BALB/c female nude mice 6 weeks of age were obtained from Wanlei Biotechnology (Shenyang, China) and were fed in standard conditions. One week after adaptive feeding, they were randomly divided into four groups: Ad-NC in MDA-MB-231, Ad-LINC00982 in MDA-MB-231, Ad-NC in MCF-7, and Ad-LINC00982 in MCF-7. BC models were established by injecting 1 × 107 Ad-infected cells into the subcutaneous flanks of nude mice. Tumor volumes were measured every 5 days. About 30 days after implantation, the mice were executed and the tumors were obtained for further analysis. Replicate experiments were performed six times. The animal research was approved by the Ethics Committee of Shengjing Hospital of China Medical University (2019PS339K).
Statistical analysis
All experiments were conducted at least three times independently. Data were analyzed using Student's t-test, one-way analysis of variance, or chi-square test and presented as the mean ± standard deviation. The p-values <0.05 were considered statistically significant.
Results
LINC00982 is downregulated in BC tissues and the low LINC00982 level predicts poor prognosis of BC
To explore the functions of LINC00982 in BC, we analyzed the expression of LINC00982 in normal and tumor tissues from the TCGA database through the GEPIA website. As shown in Figure 1a, LINC00982 was significantly suppressed in tumor tissues (n = 1085) in contrast to normal tissues (n = 291). Its expression levels were also analyzed at different stages of tumor development. Results showed that LINC00982 expression declined with the tumor stage (Fig. 1b). Furthermore, the mean value of LINC00982 was used as the threshold to analyze the relapse-free survival of BC, and the Kaplan–Meier analysis results suggested that the patients with low-LINC00982 levels exhibited lower relapse-free survival rate (Fig. 1c).
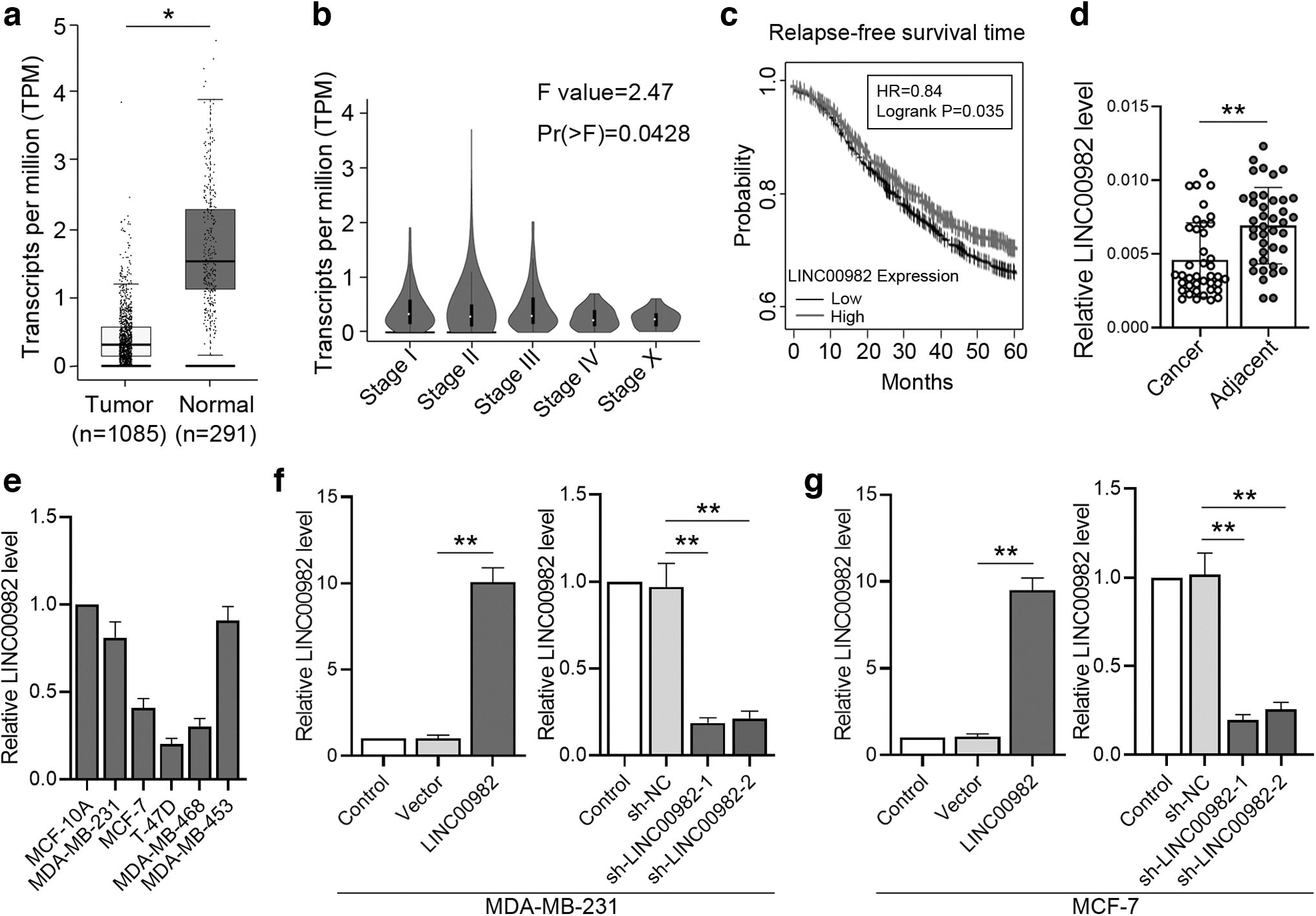
Expression of LINC00982 in the tissues and cells of BC.
Additionally, 40 pairs of cancer tissues and adjacent tissues were collected to determine LINC00982 expression by RT-qPCR. The results showed that LINC00982 expression significantly decreased in BC compared with the adjacent tissues (Fig. 1d). The correlation between clinicopathologic features and LINC00982 expression of the 40 BC patients was also analyzed. As shown in Table 2, the LINC00982 level was closely related to the tumor diameter (p-value = 0.018) and TNM stage (p-value = 0.017), whereas the level was not correlated with the other features of BC.
The Correlation Between Clinicopathologic Features and LINC00982 Level of Breast Cancers
Bold indicates significant difference at p < 0.05.
Meanwhile, we measured the expressions of LINC00982 in different cell lines (MDA-MB-231, MCF-7, T-47D, MDA-MB-468, and MDA-MB-453) and immortalized breast epithelial cell line MCF-10A using RT-qPCR. Compared with the normal breast epithelial cell line MCF-10A, the expression of LINC00982 in BC cell lines was declined to various degrees (Fig. 1e). Next, to explore the function of LINC00982, we designed different adenoviruses to overexpress or knockdown LINC00982, respectively. RT-qPCR results presented that the expression levels of overexpressing LINC00982 markedly increased in MDA-MB-231 and MCF-7 cell lines, and two pieces of LINC00982 shRNAs significantly repressed the expression of LINC00982 (Fig. 1f, g). These findings illustrated that LINC00982 was poorly expressed in BC tissue and was closely related to the development period of BC.
LINC00982 inhibits BC cell proliferation
To assess the underlying function of LINC00982 in BC cells, we first investigated the effect of overexpression or interference of LINC00982 on the proliferation of MDA-MB-231 and MCF-7 cells by MTT assay. When LINC00982 was overexpressed, the cell proliferation was dramatically suppressed in contrast to control cells at 48, 60, and 72 h (Fig. 2a). When LINC00982 was downregulated, the proliferation was significantly promoted at 48, 60, and 72 h (Fig. 2b). Furthermore, flow cytometry was used to detect cell cycle in different treatments. After LINC00982 was overexpressed, the number of cells in the G1 phase was significantly more than that in control cells, and in the S phase, the number declined (Fig. 2c). After LINC00982 was inhibited, the results were opposite to the above (Fig. 2d).

Effects of LINC00982 on BC cell proliferation.
Moreover, the western blot was used to test the changes in the expression of proliferation marker proteins. The expression of cyclin D1 and CDK4 were both suppressed when LINC00982 was upregulated (Fig. 2e), and those were increased when LINC00982 was suppressed (Fig. 2f). Thus, our results indicated that LINC00982 could affect the cell cycle and suppress cell proliferation in vitro.
LINC00982 suppresses the migration and invasion of BC cells
To investigate the effect of LINC00982 on BC cell migration and invasion, the transwell migration and invasion assays were performed in MDA-MB-231 and MCF-7 cell lines. The results of the transwell migration assay showed that the migratory abilities were significantly suppressed by overexpressing LINC00982 (Fig. 3a). When LINC00982 was silenced by shRNAs, the number of migrated cells was notably increased (Fig. 3b). The transwell invasion assay demonstrated that invasive ability of BC cells was inhibited by overexpressing LINC00982 and promoted by sh-LINC00982 obviously (Fig. 3c, d). Therefore, our results indicated that LINC00982 could restrain BC cell migration and invasion.

Effect of LINC00982 on BC cell migration and invasion.
In addition, the expression of marker proteins for cancer cell migration and invasion was detected by immunofluorescent staining and western blot. E-cadherin, an invasion suppressor, plays a critical role in maintaining intercellular adhesion (Takeichi, 1993). The results of immunofluorescent staining showed that the expression of E-cadherin was increased when LINC00982 was overexpressed, and the expression of E-cadherin declined when LINC00982 was silenced.
Matrix metalloproteinase-2 (MMP-2) belongs to the zinc-dependent endopeptidase family and participates in the degradation of the tumor basal membrane (Bäck et al., 2010). MMP-2 contributes to cancer cell invasion by extracellular matrix degradation through matrix proteolysis (Mendes et al., 2007). Numerous studies have demonstrated that vimentin was a metastasis-associated factor in some human cancers (Li et al., 2010), including BC (Zhou et al., 2008). Results of western blot suggested that the expression of MMP-2 and vimentin reduced when LINC00982 was upregulated (Fig. 4c) and increased after downregulating LINC00982 (Fig. 4d). All the above results confirmed that LINC00982 obstructed the migration and invasion of BC cells.

Effect of LINC00982 on invasion-related proteins.
LINC00982 affects cell behavior through STAT3/NF-κB signaling pathway in BC cells
STAT3 is one of the most important members of the STAT family, which regulates a diverse array of cellular processes (Banerjee and Resat, 2016). Recent evidence suggests that activated STAT3 plays a crucial role in promoting tumor cell proliferation, survival, and invasion in BC. STAT3 also participated in tumor-promoting inflammation by influencing pro-oncogenic inflammatory pathways, including NF-κB (Yu et al., 2009).
NF-κB is a significant pathway that contributes to immune and inflammatory responses. It relies mainly on the activation of the p65 to control the transcription of disease-related key genes (Giridharan and Srinivasan, 2018). Therefore, western blot was used to detect the alteration of activated STAT3 and p65 when LINC0098 was overexpressed or inhibited. As shown in Figure 5a, compared with blank vector treatment, the expression of activated STAT3 and p65 are both decreased when LINC0098 was overexpressed.
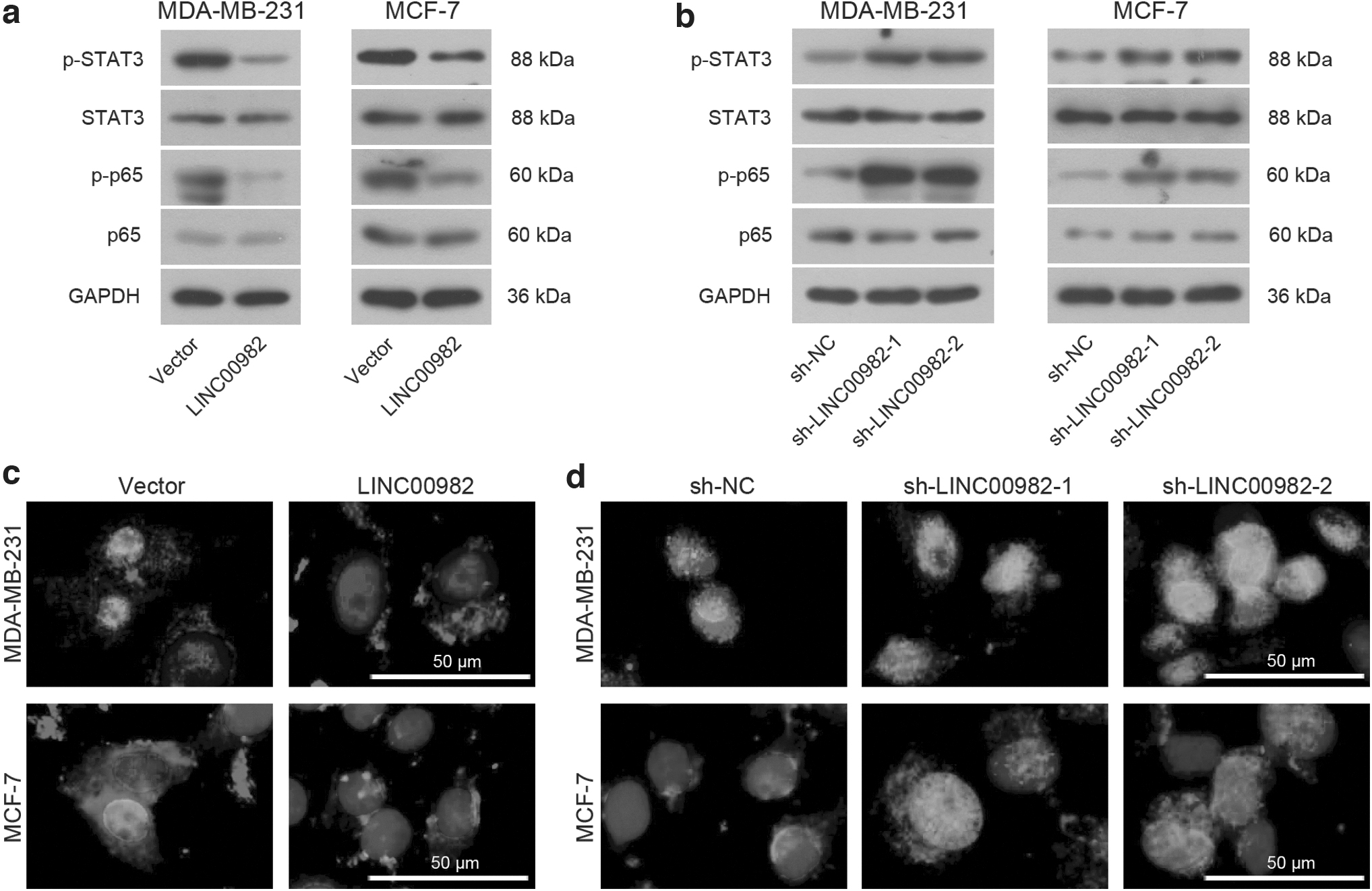
Effects of LINC00982 on the STAT3/NF-κB pathway in BC cells.
In Figure 5b, activated STAT3 and p65 were facilitated by sh-LINC00982. In addition, previous studies present that p65 will be phosphorylated into the nucleus and regulate transcription upon stimulation, so that the location of activated p65 was detected by immunofluorescent staining. As shown in Figure 5c, the p65 protein stained with red fluorescence in the nucleus was significantly reduced when LINC0098 was overexpressed, and it was advanced by sh-LINC0098 treatment (Fig. 5d). These findings demonstrated that LINC00982 could regulate STAT3/NF-κB pathway.
Verification of the regulation of LINC00982 on miR-765/DPF3 axis
Previous studies have verified that lncRNAs can be miRNA sponges to regulate cell behaviors. A binding site of miR-765 was found on LINC00982, then, the WT or mutated type (MT) binding sequence (Fig. 6a) was inserted into the luciferase reporter vector. Through the luciferase reporter assays, we found that relative luciferase activity in miR-765 mimic and LINC00982 WT cotransfected cells was significantly decreased in contrast to other cells (Fig. 6b). After overexpression of LINC00982, miR-765 expression was significantly downregulated. When LINC00982 was knockdown, the expression level of miR-765 was markedly upregulated (Fig. 6c). Therefore, we verified LINC00982 could target miR-765. According to previous studies, miRNAs usually are assumed to exert functions through directly regulating their target genes (Zhang et al., 2021).
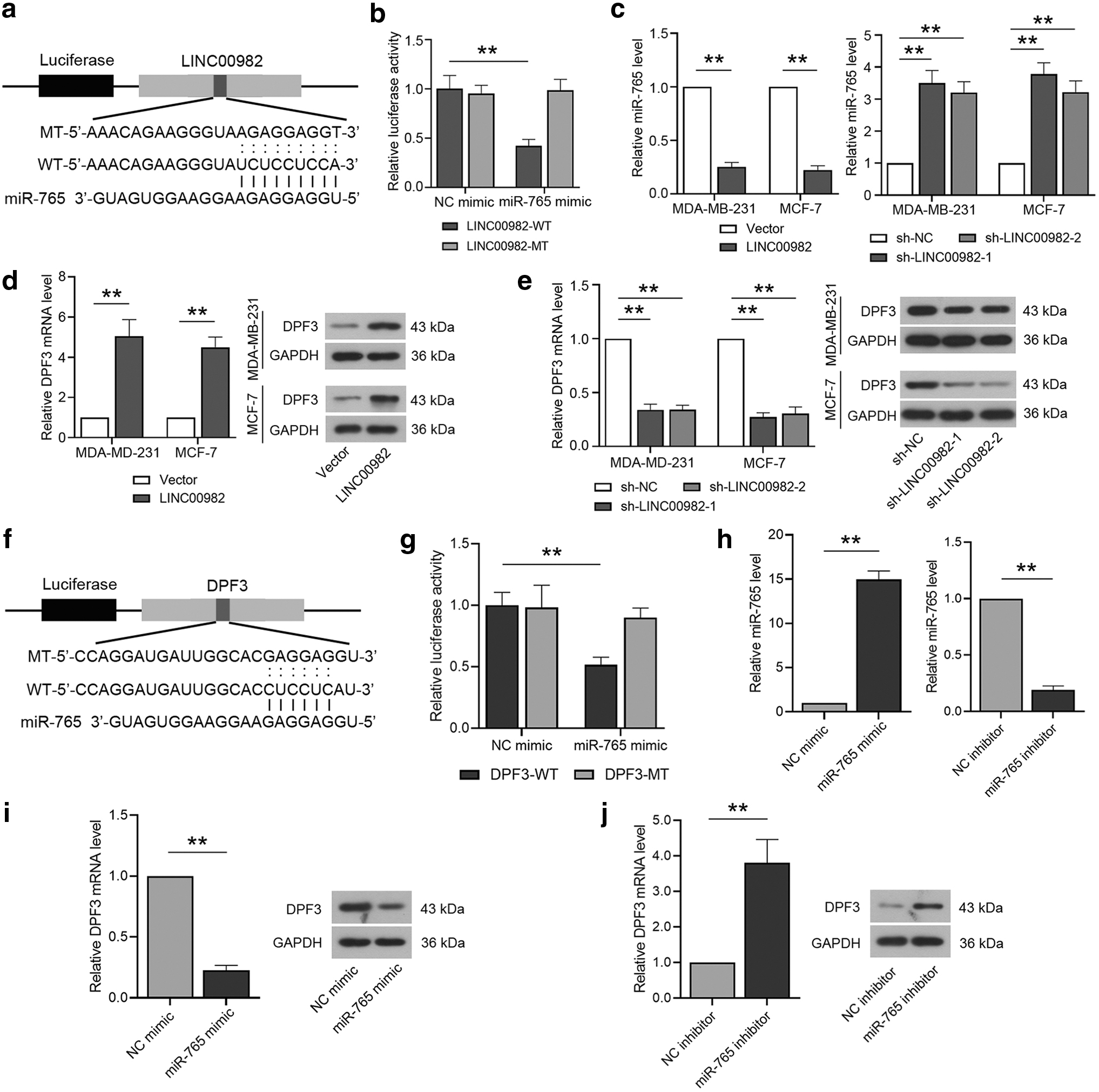
Verification the regulation of LINC00982 on miR-765/DPF3 axis.
Then a target of miR-765, DPF3 was detected, the results explained that the expression of DPF3 mRNA and protein was dramatically increased when LINC00982 was overexpressed (Fig. 6d). Subsequently, the expression of DPF3 mRNA and protein was significantly downregulated with sh-LINC00982 adenovirus infecting (Fig. 6e). These findings suggested that LINC00982 could regulate the expression of DPF3 as a molecular sponge of miR-765.
Thus, a target of miR-765 was found and the WT or mutated type (MT) binding sequence (Fig. 6f) was inserted into the luciferase reporter vector. It is found that the luciferase activity was decreased when the cells were cotransfected with miR-765 mimic and DPF3 WT (Fig. 6g). The efficiency of miR-765 mimic and inhibitor was confirmed by RT-qPCR in the MDA-MB-231 cell line (Fig. 6h). Subsequently, the RT-qPCR and western blot results showed that DPF3 was negatively regulated by miR-765 (Fig. 6i, j). All these findings indicated that miR-765 could target DPF3 directly and affect its expression.
The effect of LINC00982 on the malignant phenotype of BC cells is rescued by the interference of DPF3
Adenovirus-mediated downregulation of DPF3 was conducted to explore the effect of DPF3 on the malignant phenotype of BC, and the knockdown efficiency was confirmed by western blot (Fig. 7a). The results of the MTT assay showed that LINC00982 caused suppression of cell proliferation and was reversed by the downregulation of DPF3 (Fig. 7b). Similarly, LINC00982 induced inhibition on the migration and invasion of MDA-MB-231 cells, and all were disturbed by knockdown of DPF3 (Fig. 7c, d). Furthermore, the western blot results showed that repression on the STAT3/NF-κB signaling pathway induced by LINC00982 overexpression was also blocked by the knockdown of DPF3 (Fig. 7e). These data suggested that interference of DPF3 could rescue the effect of LINC00982 on the malignant phenotype of BC cells.

Effects of the LINC00982/miR-765/DPF3 axis on the development of BC.
Overexpression of LINC00982 inhibits tumorigenesis by regulating the miR-765/DPF3 axis in BC cells in vivo
The LINC00982-overexpressed MDA-MB-231 and MCF-7 cell lines were injected into the subcutaneous flanks of nude mice, and the tumor volume was measured every 5 days. As depicted in Figure 8a and b, upregulation of LINC00982 significantly slowed down the tumor growth. Then, RT-qPCR was used to test the expression of LINC00982 and miR-765 of the xenografts. The results showed that the expression of LINC00982 was increased and miR-765 was reduced in LINC00982-overexpressed xenografts (Fig. 8c, d). At the same, the mRNA and protein expression of DPF3 has significantly increased in LINC00982-overexpressed xenografts (Fig. 8e, f), and these were further verified by immunohistochemistry (Fig. 8g). The above findings illustrated that overexpression of LINC00982 inhibits tumorigenesis through the miR-765/DPF3 axis in BC in vivo.

Effects of LINC00982 on development of tumor growth for BC in vivo.
Discussion
BC is one of the most common cancer that has been gradually increased in women all over the world (DeSantis et al., 2014; Donepudi et al., 2014). BC has seriously threatened women's health and is a medical problem that needs to be solved urgently. Therefore, the exploration of molecular mechanisms related to the growth and progression of BC tumors and the research and development of reliable biomarkers should be effective ways to break through this problem.
In this study, we identified that LINC00982 is downregulated in BC tissues and the low LINC00982 level predicted a poor prognosis of BC. Upregulation of LINC00982 significantly suppressed the proliferation, migration, and invasion of BC cells. Our findings demonstrate that LINC00982 is an essential role in BC cell progress, and it may function as a potential target for diagnosis, prognosis, or even therapy in BC patients.
During the past researches, lncRNAs are assumed to play an important role in multiple physiological processes of cells and have attracted widespread attention. There are some researchers who found that lncRNA TINCR promoted cell proliferation, invasion, and metastasis of BC cells and inhibited cancer cell apoptosis.
Meanwhile, they demonstrate that miR-589-3p is a target molecule for TINCR and interacts with gene IGF1R to affect cancer cell development (Guo et al., 2020). In addition, lnc-408 is highly expressed in BC cells, which promotes cell invasion and metastasis. Enhanced lnc-408 is tightly related to BC progression, clinical stages, and pathological grades in patients with BC (Qiao et al., 2021). In the two previous studies, lncRNAs act as cancer promoters in the process of BC. The present study has confirmed LINC00982 could be a cancer suppressor to inhibit BC cell proliferation, migration, and invasion, and obstruct BC tumor growth in vivo.
Increasing evidence support that plenty of lncRNAs affects human disease development through competitive endogenous RNA (ceRNA) regulation (Wang et al., 2010; Lu et al., 2020; Zhang et al., 2021). This hypothesis was proposed by Salmena et al. (2011), and its main concept is that ceRNAs interact with targeting miRNAs to modulate downstream target genes and to further regulate cell procedure (Zhou et al., 2019).
The previous report demonstrated that lncRNA TINCR acted as a ceRNA to sponge miR-589-3p, promoting the Akt pathway through the IGF1R gene, thus advancing the malignant phenotype of BC and suppressing cell apoptosis (Guo et al., 2020). Recent reports confirmed that TMPO-AS1 functioned as a molecular sponge, modulating miR-577/RAB14 expression to enhance cancer progression (Hou et al., 2021). To further explore the mechanism of LINC00982 to regulate BC, we speculated from these existing researches and found that lncRNAs usually functioned as a ceRNA to interact with miRNAs to participate in cell procedure of human diseases.
Through luciferase report assay, RT-qPCR, and western blot, we confirmed that LINC00982 was a molecular sponge to regulate the miR-765/DPF3 axis. In addition, it was identified that DPF3 could rescue the effect of overexpressing LINC00982 to cancer cells. In previous studies, LINC00982 inhibits cell proliferation in renal cancer and papillary thyroid carcinoma, respectively, through regulating the PI3K/AKT signaling pathway (Xu et al., 2019; Zhang et al., 2019), and it also downregulated in gastric cancer and lung adenocarcinoma (Fei et al., 2016; Lv et al., 2018). Our study demonstrated that LINC00982 could function as a molecular sponge to modulate miR-765/DPF3 axis to inhibit the development of BC.
Targeting signaling pathways, which are related to human cancer, have always been a potentially effective method for cancer treatment (Dancey et al., 2012). STAT is a unique protein family that could bind to DNA directly (Darnell, 1997). STAT3, a crucial nuclear transcription factor, is one of the most important members of the STAT family (Landy et al., 2016; Zhang et al., 2016). NF-κB is a significant pathway that participated in tumor cell progression (Zhang et al., 2017). CyclinD1, c-myc, and plenty of cell proliferation-related proteins are downstream molecules of NF-κB signaling (Yang et al., 2010, 2018). The p65 plays an important role in controlling the transcription of disease-related key genes (Giridharan and Srinivasan, 2018).
Many studies have identified that the activity of STAT3 and NF-κB signaling have a close relationship with cancer development, and the inhibition of the STAT3 pathway could suppress different kinds of human cancers (Chai et al., 2016; Nunez Lopez et al., 2016). In our study, the protein expression of STAT3, p-STAT3, p65, and p-p65 were detected by western blot when LINC00982 overexpressed or knocked down.
The results told us that the activity of STAT3 and p65 were suppressed by the upregulation of LINC00982, and the results showed the opposite when LINC00982 was downregulated. Moreover, the expression changes of DPF3 also affected the activity of STAT3 and p65. p-STAT3 and p65 were increased with the downregulation of DPF3, in that, our research illustrated that LINC00982 suppressed cell proliferation, migration, and invasion through STAT3/NF-κB pathway in BC.
Conclusions
In summary, LINC00982 could suppress the malignant behavior of BC by the repression of the STAT3/NF-kB pathway, and these effects seem to be mediated by the miR-765/DPF3 axis. Overexpression of LINC00982 may serve as a potential therapeutic strategy for BC in the future.
Footnotes
Disclosure Statement
No competing financial interests exist.
Funding Information
This research was funded by the Hospital Project of Shengjing Hospital of China Medical University.
Supplementary Material
Supplementary Data
References
Supplementary Material
Please find the following supplemental material available below.
For Open Access articles published under a Creative Commons License, all supplemental material carries the same license as the article it is associated with.
For non-Open Access articles published, all supplemental material carries a non-exclusive license, and permission requests for re-use of supplemental material or any part of supplemental material shall be sent directly to the copyright owner as specified in the copyright notice associated with the article.