
Abstract
Accumulating evidence implies that long noncoding RNAs participate in non-small cell lung cancer (NSCLC) tumorigenesis. Our current study synthetically analyzed RNA sequencing data downloaded from The Cancer Genome Atlas (TCGA) and Gene Expression Omnibus (GEO) databases. We identified LINC00961 significantly downregulated in NSCLC tissues. We explored the LINC00961 expression in NSCLC tumor tissues and cell lines with reverse transcription-quantitative polymerase chain reaction analysis. Lentivirus-mediated infection upregulated the expression of LINC00961 in A549 cells. The proliferation and migration capability were also measured in A549 cells. In addition, we performed luciferase reporter gene assay to investigate whether LINC00961 directly interacts with miR-19a-3p/miR-19b-3p/miR-125b-5p. A nude mice model was used to detect the potential biological process of LINC00961 on tumor growth in vivo. The results showed that LINC00961 was significantly down-egulated in NSCLC tissues and cell lines. LV-LINC00961 effectively increased the expression of LINC00961 and decreased the expression of miR-19a-3p/miR-19b-3p/miR-125b-5p. LINC00961 upregulation remarkably inhibited cell proliferation, migration, and invasion while promoting cell apoptosis in A549 cells. Luciferase reporter gene assay revealed that LINC00961 could directly sponge miR-19a-3p/miR-19b-3p/miR-125b-5p. Moreover, overexpressed miR-19a-3p/miR-19b-3p/miR-125b-5p reversed the effect of LINC00961 on cell function of A549 cells. Western blot assays revealed that LINC00961 could partially act as a tumor suppressor via affecting PI3K-AKT/MAPK/mTOR signaling pathway. In addition, overexpressed LINC00961-inhibited tumor growth was demonstrated in vivo. Overexpression of LINC00961 inhibited cell viability, invasion, and induced apoptosis in NSCLC, potentially via suppressing the expression of miR-19a-3p/miR-19b-3p/miR-125b-5p by targeting PI3K-AKT/MAPK/mTOR signaling pathways, which might provide the potential biomarker for NSCLC diagnosis and therapies.
Introduction
The latest findings disclose that lung cancer is still the most common cancer in both incidence and mortality for both men and women in 2020 (Brenner et al., 2020; Jung et al., 2020; Siegel et al., 2020). It is identified several types for lung cancers, including non-small cell lung cancer (NSCLC) (such as adenocarcinomas, squamous carcinomas, large cell carcinomas, and several other types) and small cell lung cancer (SCLC), (de Sousa and Carvalho, 2018).
SCLC accounts for about 15–17% of all diagnosed lung cancers characterized by early metastatic spread, rapid growth, and initial response to treatment, with a less than 7% 5-year survival rate (Tsoukalas et al., 2018). As the largest subgroup of lung cancer, NSCLC has become a key issue of global research due to its large proportion (∼85%), easy misdiagnosis, and unobvious symptoms (Jonna and Subramaniam, 2019).
The conventional diagnosis methods of NSCLC include X-ray, computed tomography (CT) scan, magnetic resonance imaging (MRI), positron emission tomography (PET) scan, bronchoscopy, and tissue biopsy, which have relatively low rates of early diagnosis (Goebel et al., 2019). Dozens of biomarkers for the early diagnosis of NSCLC have been identified, such as carcinoembryonic antigen, cytokeratin 19 fragments antigen (CYFRA21-1), squamous cell carcinoma antigen (SCC-Ag), carbohydrate antigen 125 (CA125), CA15-3, CA19-9, and CA72-4, which have improved the early diagnosis rate of NSCLC, and improved patients quality of life (Chen et al., 2018b). In the past decade, the increasing number of clinically available inhibitors of targeted signal transduction pathways have provided enormous opportunities to improve NSCLC patient prognosis.
Compared with standard chemotherapy, treatment of mutations or fusions in epidermal growth factor receptor, anaplastic lymphoma kinase, proto-oncogene receptor tyrosine kinase (ROS1), and serine/threonine-protein kinase b-raf (BRAF) with targeted inhibitors significantly improves the prognosis of patients with NSCLC (Li et al., 2021; Liu et al., 2020; Roviello et al., 2021). However, due to the targeted oncogenes (on-target resistance) and changes in other related signaling pathways (off-target resistance) in NCSLC cells, great majority patients develop resistance to these targeted drugs after 1 year (Liu et al., 2020).
Despite the development of diagnosis, current therapies, and new targeted treatments, the 5-year overall survival rate of NSCLC cases remains less than 15% (Hirsch et al., 2017; Nasim et al., 2019). Therefore, it is vital to identify the novel molecules of pathophysiological mechanism to advance treatment strategies for NSCLC cases.
Recent studies reveal that noncoding RNAs (ncRNAs) play essential biological roles in the carcinogenesis and development of cancers, including cell proliferation, differentiation, and apoptosis (Anastasiadou et al., 2018; Chan and Tay, 2018; Romano et al., 2017).
Long noncoding RNAs (lncRNAs) constitute a significant class of ncRNAs, more than 200 nucleotides (nt) in length without the protein-coding ability (Kapranov et al., 2007). Because of the more substantial tissue specificity, lncRNAs have been more effective as biomarkers of diagnosis and prognosis of diseases (Hung and Chang, 2010). An increasing number of studies suggest that differentially expressed lncRNAs have participated in the occurrence and development of multiple diseases, including NSCLC (He et al., 2017; Yang et al., 2019; Yu et al., 2019).
For instance, LncRNA-XIST was overexpressed and identified as an oncogene by suppressing the expression level of KLF2 in NSCLC (Fang et al., 2016). MicroRNA (miRNA) has been proved to be an important ncRNA in the development of NSCLC (Sui, 2020). Recent studies show that lncRNA might function as molecular sponges for miRNA to regulate progression of cancer (Salmena et al., 2011). For example, a high lncRNA DLEU2 expression level predicted shorter overall survival and promoted tumor growth and metastasis by regulating the miR-30c-5p/SOX9 axis in NSCLC (Zhou et al., 2019). Although various underlying functions and mechanisms of lncRNAs in NSCLC have been characterized, the majority of lncRNAs contributing to the biological properties of NSCLC remain inconclusive.
Long intergenic noncoding RNA 00961 (LINC00961) is 1546 nt located in chromosome 9 with the ability to encode the polypeptide-small regulatory polypeptide of amino acid response (Matsumoto et al., 2017). Importantly, recent studies have demonstrated that LINC00961 has been a novel biomarker in multiple types of cancers, such as renal cell carcinoma (Chen et al., 2019a), melanoma (Mu et al., 2019), and glioma (Lu et al., 2018). In addition, LINC00961 has also been reported to be correlated with the survival and progression of NSCLC (Huang et al., 2018b; Jiang et al., 2018). However, the potential mechanism of LINC00961 in NSCLC has never well been investigated.
In the present study, we first explored the LINC00961 expression of NSCLC in the Cancer Genome Atlas (TCGA) database, Gene Expression Omnibus (GEO) database, patients' tissues, and cells. Furthermore, LV-LINC00961 was designed to investigate effects on cell proliferation, apoptosis, and invasion in NSCLC cells and the underlying mechanisms in NSCLC progression.
Materials and Methods
Identification of the abnormally expressed lncRNA LINC00961 in TCGA dataset and microarray gene profiling data
We downloaded lncRNA sequencing data from 521 lung adenocarcinoma (LUAC) and 504 lung squamous cell carcinoma from the TCGA data portal (
Patients and tissue samples
This study obtained 83 Chinese Han population NSCLC tissue specimens (lung tumor tissues and paired adjacent noncancerous lung tissues) from the Nanjing Chest Hospital, and the detailed clinical data are shown in Supplementary Data S1. After surgery, lung tissues were immediately cleaned with saline, frozen with RNAlater (Ambion, Foster), and stored at −80°C until further analysis. We also collected detailed paper pathology reports and quality assessment reports. Informed consent forms were obtained from the patients. The Ethics Committee approved the current study of Zhongda Hospital, Southeast University (No.: 2014ZDSYLL031.0).
Cell culture
We purchased 16HBE cells and A549 cells from the Chinese Academy of Sciences Type Culture Collection Cell Library Committee (Shanghai, China). Benzo(a)pyrene-transformed 35th-generation 16HBE cells [B(a)P 16HBE] were cultured by our group. We separately cultured 16HBE cells and B(a)P, 16HBE, and A549 cells in Dulbecco's modified Eagle's medium and 1640 cell culture medium (HyClone Laboratories, Inc., Logan), supplemented with 10% fetal bovine serum (HyClone, Thermo Fisher Scientific), and 100 U/mL penicillin and 100 mg/mL streptomycin (Thermo Fisher Scientific), in a humidified 5% CO2 incubator at 37°C.
Detection of the expression level of LINC00961
According to the manufacturer's instructions, the total RNAs were extracted from lung tissues and cells using TRIzol LS (Genstar, Beijing, China). RNAs reverse-transcribed to cDNA using StarScript II M-MLV (Genstar) and through real-time PCR to detect the expression level of LINC00961 using the Step One Plus™ PCR System (Applied Biosystems, Thermo Fisher Scientific), and RealStar Green MIX (Genstar) was used for quantitative real-time polymerase chain reaction (qRT-PCR) detection. The LINC00961 sense primer was 5′-GATTTGCTGTGCTGATGG-3′, and the reverse primer was 5′-GTTACTGGAGGGGAAGGA-3′. The housekeeping gene GAPDH was used as a control, the GAPDH sense primer was 5′-GGGAGCCAAAAGGGTCATCA-3′, and the reverse primer was 5′-TGATGGCATGGACTGTGGTC-3′. The qRT-PCR data were analyzed and calculated by the 2−ΔΔCt method.
Plasmid transfection
The LINC00961 vectors were constructed at Genechem Co., LTD Shanghai, China. In the present study, LINC00961 upregulated cell transfection through the Ubi-MCS-SV40-EGFP-IRES-puromycin-LINC00961 lentivirus vector (LV-LINC00961). A549 cells were cultured in 12-well plates and transfected with LV-LINC00961, according to the manufacturer's protocol. After culturing 48 h, stably transfected A549 cell lines were screened with puromycin. Finally, the stable transfection A549 cell line expression level of LINC00961 was detected by qRT-PCR.
Cell proliferation detection
The Cell Counting Kit-8 (CCK-8) assay was used to assess cell proliferation. Five thousand LV-LINC00961 and Negative Control A549 cells were separately seeded into each well of a 96-well plate. Individually, cells were cultured in a complete culture medium for 12, 24, 48, and 72 h. The culture supernatants were removed, and 100 μL solution (CCK-8:1640 = 1:10) was added to each well and incubated for 1 h at 37°C. The optical density value was measured at 450 nm in a microtiter plate reader.
Flow cytometric analysis
LV-LINC00961 and Negative Control A549 cells were seeded into a six-well plate. Cells were cultured in a complete culture medium for 48 h. Cells were harvested by trypsinization without EDTA and were washed twice in ice-cold phosphate-buffered saline (PBS). According to the manufacturer's protocol, after staining with Annexin V-APC and propidium iodide (PI) (KeyGEN BioTECH, China), the cells were analyzed for apoptosis using a flow cytometer (BD Biosciences).
Cell migration and invasion assays
Cells were separately seeded into six-well plates for cell migration assays and cultured normally until reaching 95% confluency, and the scratch was made in the container using a sterile 200 μL pipette tip. Subsequently, detached cells were washed with PBS, and the cells were incubated with 5% CO2 at 37°C. Finally, microscope photographs were taken at a different time node, photographed using a digital camera system (FSX100 Biological Image System, Olympus).
For invasion assay, 5 × 104 cells were seeded into the 24-well Transwell plate upper chamber, which insert coated with Matrigel. After culturing for 24 h, cells on the bottom side of the upper chambers' membrane were fixed by methanol for 10 min, then stained by crystal violet dye for 5 min, and finally imaged using six random visions (Olympus, Tokyo, Japan).
Luciferase assays
LncBase Experimental v.2 (
Western blotting
According to the article, the total proteins were prepared in cell lysis buffer containing 1% protease inhibitor (Thermo Fisher) and quantified by bicinchoninic acid protein assay kit (KeyGEN BioTECH). The sodium dodecyl sulfate-polyacrylamide gel electrophoresis (SDS-PAGE) was performed to load the denatured proteins and transferred to a polyvinylidene difluoride (PVDF) membrane. After blocking in 5% skim milk for 2 h at 37°C, PVDF membranes were incubated for 12 h at 4°C with primary antibodies. After washing for 30 min, PVDF membranes were incubated with the corresponding secondary antibodies for 2 h at 37°C. The chemiluminescence chromogenic substrate (Millipore, Billerica) was used to visualize protein bands, and densitometry analysis was performed with Tannon 5200. GAPDH was used as a housekeeping protein.
In vivo experiment
The Animal Experimental Ethical Inspection Form approved all procedures of animal experiments of Southeast University (No.: 20210221002). BALB/c nude mice aged 4 weeks and weighing ∼20 g (GemPharmatech, Nanjing, China) were divided into two groups and kept in an aseptic, constant temperature, and humidity condition. Tumors were generated by subcutaneous injection with 1 × 108 A549 cells. Tumor volume and weight were determined at the end of 28 days after sacrificing. The tumor volume was calculated with the formula: 1/2 (length × width)2. The tumors were immediately excised, imaged, and weighted. Finally, the expression levels of LINC00961, possibly targeted miRNAs, and critical proteins were detected in the tumor by qRT-PCR and western blotting.
Statistical analysis
All statistical analyses were performed with IBM SPSS 21.0 software (Chicago). The results are expressed as mean ± standard deviation. Statistical significance was determined using the Student's t-test and one-way analysis of variance as appropriate. p-Value <0.05 was considered statistically significant.
Results
Expression of LINC00961 in TCGA, GEO, lung tumor tissues, and cells
To investigate the potential role of LINC00961 in NSCLC, the expression of LINC00961 was first detected in TCGA and GEO datasets.
LINC00961 expressions were calculated by TCGA RNA sequencing data, which showed that LINC00961 was indeed downregulated in LUAD and LUSC (Fig. 1A). Moreover, LINC00961 expression was significantly downregulated in the GSE19188 dataset (−1.39-fold-change), GSE30219 dataset (−1.43-fold-change), GSE27262 dataset (−1.72-fold-change), GSE18842 dataset (−1.72-fold-change), GSE19804 dataset (−1.49-fold-change), and GSE31210 dataset (−2.86-fold-change) (Fig. 1A). As shown in Figure 1B, the LINC00961 level was significantly decreased in lung tumor tissues compared with matched adjacent noncancerous tissues. Similarly, LINC00961 expression levels were also downregulated in B(a)P, 16HBE, and A549 cells compared with 16HBE cells (Fig. 1C).
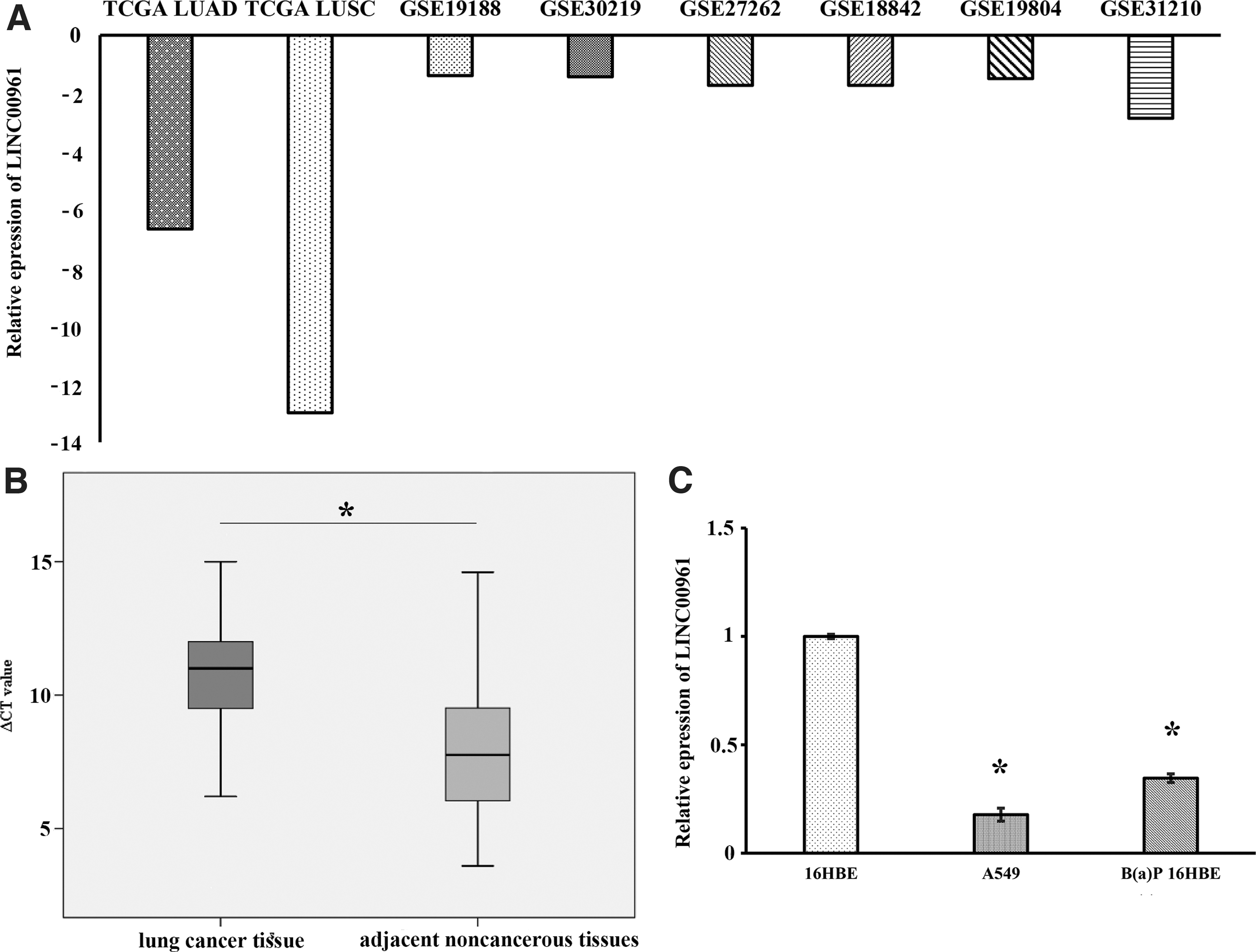
Expression profile of LINC00961 in NSCLC tissues and cell line.
Lentivirus-mediated infection altered the in vitro expression of LINC00961
First, lentivirus vectors were constructed and infected into A549 cells. Subsequently, the lentivirus stably transfected A549 cells were screened by puromycin (Fig. 2A). Finally, LINC00961 expression level detection results showed that the LINC00961 expression was significantly upregulated in LV-LINC00961, A549 cells compared with negative control cells (Fig. 2B).

Lentivirus-mediated infection altered the in vitro expression of LINC00961.
LINC00961 affected A549 cell proliferation and apoptosis
The CCK-8 assay and flow cytometric assay were performed to detect the affection of overexpressed LINC00961 on A549 cell proliferation and apoptosis. CCK-8 assay results showed that A549 cells growth was significantly decreased in the LV-LINC00961 group after 48 and 72 h compared with the negative control and blank group A549 cells (p < 0.05, Fig. 3A). Moreover, there was no significant difference in the proliferative capacity in negative control and blank group A549 cells (p > 0.05). Apoptosis of the stably transfected A549 cells was determined by flow cytometric assay, and the results revealed that apoptosis of LV-LINC00961 A549 cells was significantly increased compared with the negative control group (p < 0.05; Fig. 3B, C).
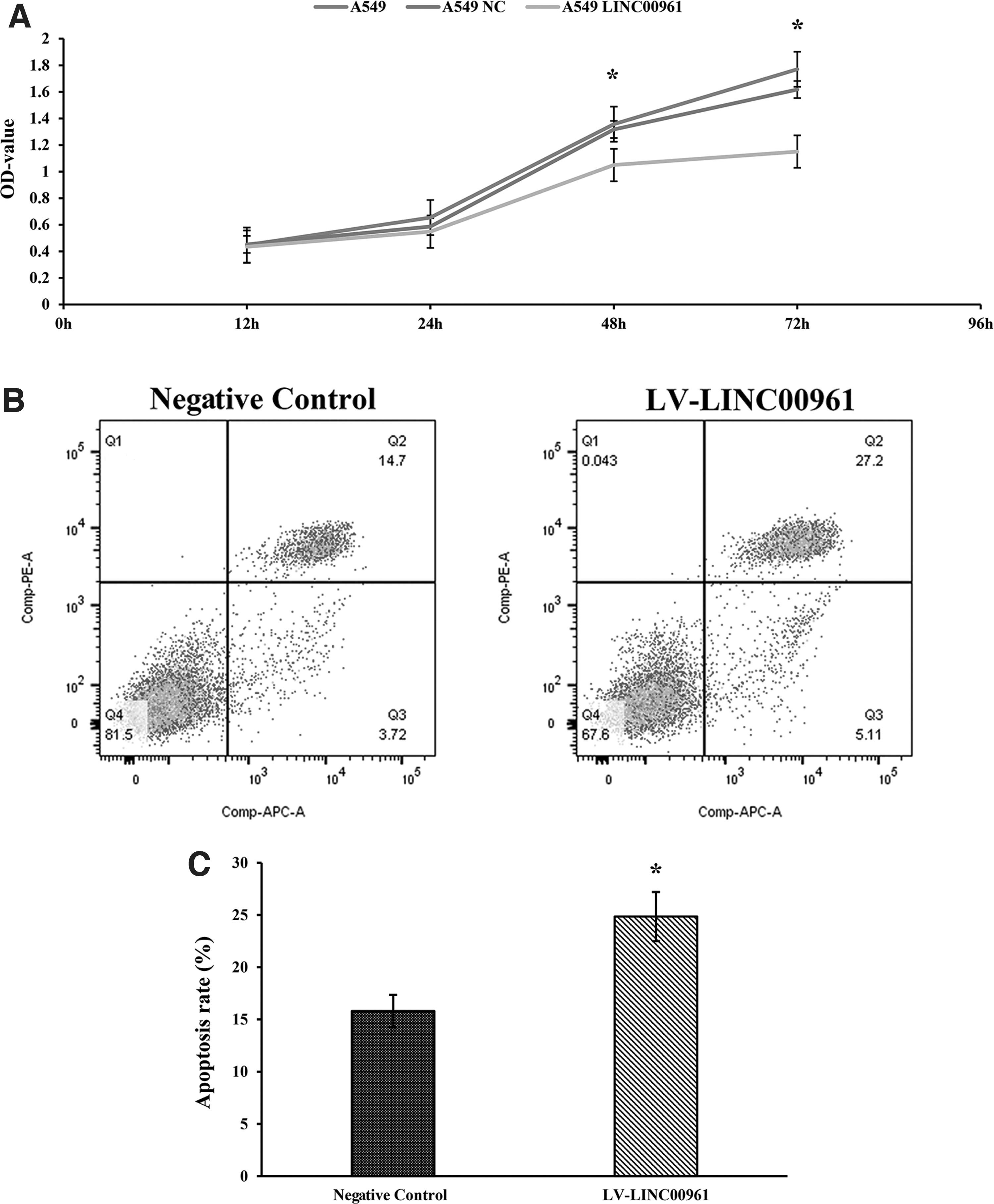
Overexpression of LINC00961 significantly suppressed A549 cell proliferation and promoted apoptosis.
LINC00961-overexpression decreased the migration and invasion of A549 cells
Cell migration and invasion are also the major malignant phenotypes of NSCLC cells and play essential roles in NSCLC progression. To detect the potential function of LINC00961, we also investigate the migration and invasion of A549 cells. The results of the migration assay revealed that the migration of LV-LINC00961 A549 cells decreased significantly compared with negative control cells (p < 0.05) after 24 and 48 h (Fig. 4A, B). In addition, the invasive assays indicated similar results when stable infection by LV-LINC00961. Overexpressed LINC00961 significantly inhibited the invasive ability of A549 cells compared with negative control cells (Fig. 4C, D). The above results suggested that LINC00961 overexpression can inhibit NSCLC cell migration and invasion.
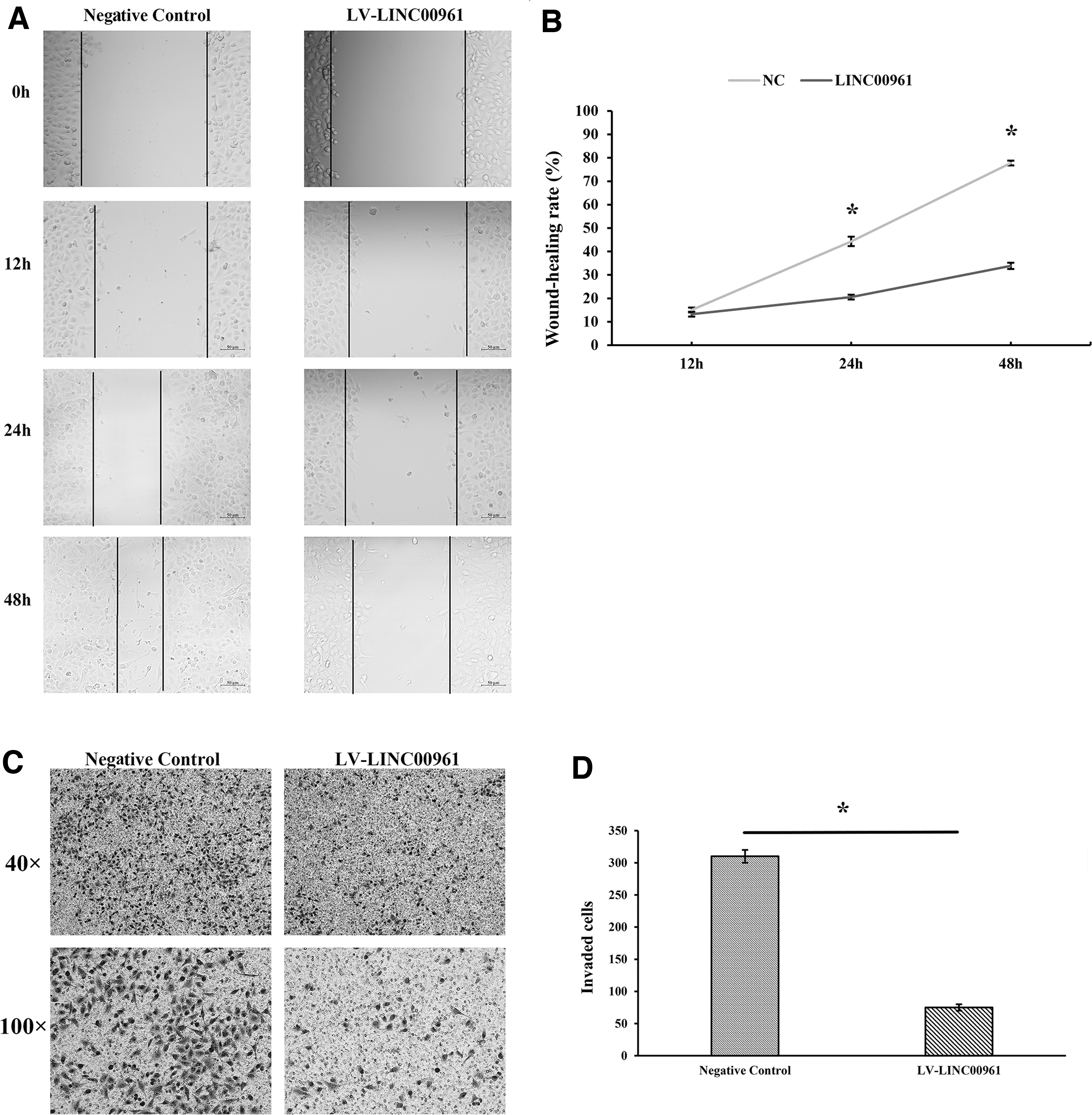
Overexpression of LINC00961 significantly suppressed A549 cell migration and invasion.
LINC00961 directly interacts with miR-19a-3p/miR-19b-3p/miR-125b-5p and rescue experiment
Previous studies have verified that lncRNAs may directly interact with the miRNAs sponge to regulate mRNAs (Sui et al., 2016). Based on LncBase Experimental v.2, LINC00961 was predicted to contain potential binding sites of miR-19a-3p, miR-19b-3p, and miR-125b-5p (Fig. 5A). To confirm the directly regulated relationship between miR-19a-3p/miR-19b-3p/miR-125b-5p and LINC00961, the luciferase reporter inserted with the whole LINC00961 sequence was constructed and transfected with miR-19a-3p/miR-19b-3p/miR-125b-5p/miR-NC into A549 cells. The results showed that miR-19a-3p/miR-19b-3p/miR-125b-5p significantly decreased luciferase activities of pmiR-LINC00961 (Fig. 5B). The results of qRT-PCR indicated that LINC00961 overexpression significantly inhibited the expression of miR-19a-3p/miR-19b-3p/miR-125b-5p (Fig. 5C).
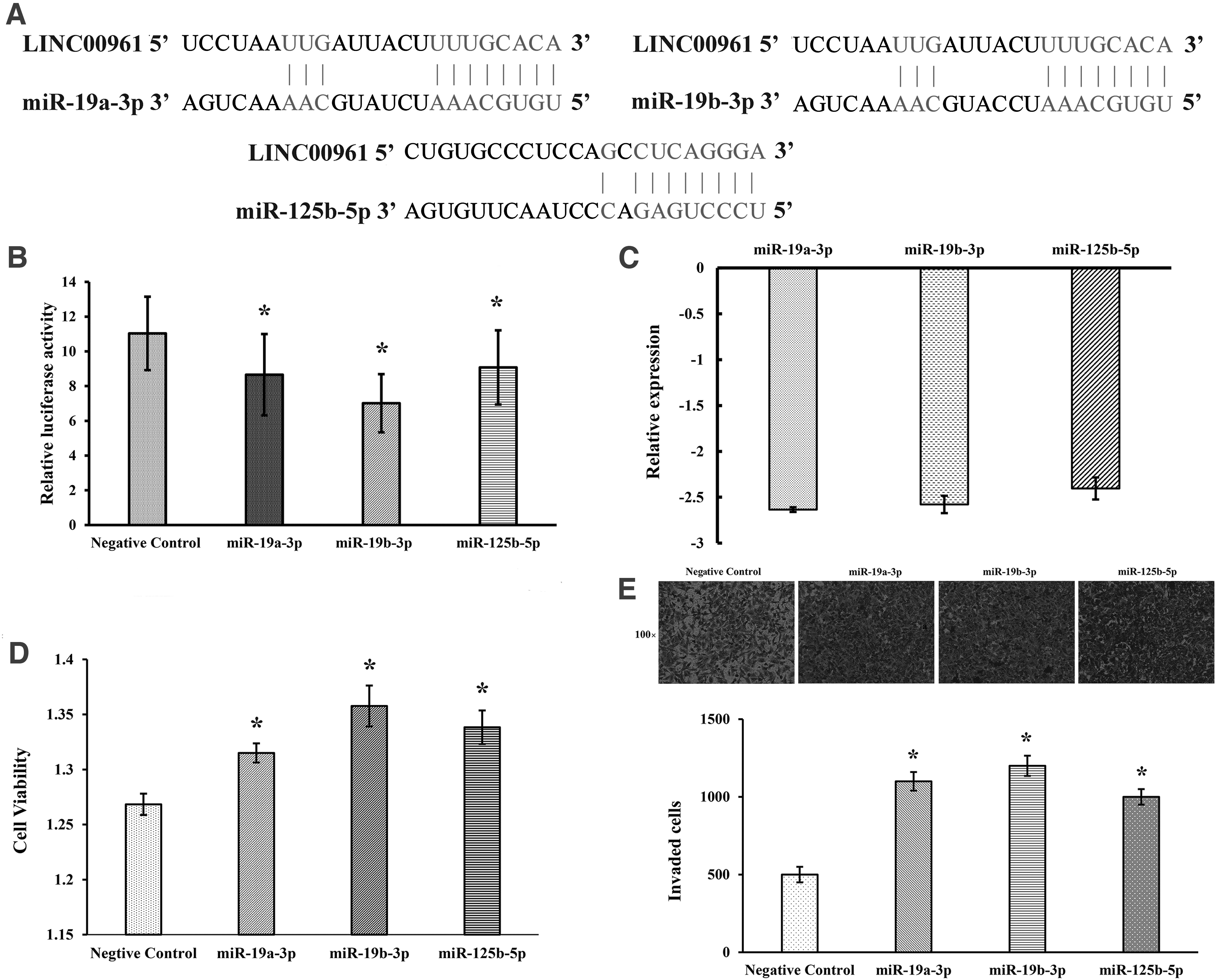
LINC00961 directly interacted with miR-19a-3p/miR-19b-3p/miR-125b-5p.
The results of rescue experiment assay showed that cotransfection of LV-LINC00961 A549 cells with miR-19a-3p/miR-19b-3p/miR-125b-5p promoted the ability of proliferation and invasion, which were inhabited by LINC00961 (Fig. 5D, E). The results indicated that LINC00961 could directly interact with miR-19a-3p/miR-19b-3p/miR-125b-5p to influence the A549 cell function.
LINC00961-overexpression influences PI3K-AKT/MAPK/mTOR signaling pathways
PI3K-AKT, MAPK, and mTOR signaling pathways are vital pathways that play essential roles in the development of lung cancer and the cellular functionality of lung cancer cell proliferation, apoptosis, invasion, and migration. After western blotting detection, the protein expression results showed that compared with the Negative control group, LV-LINC00961 group, the expression levels of essential proteins of PI3K-AKT signaling pathway such as PI3K, AKT, P-AKT, P-GSK-3β, BCL-XL, and BCL-2 were significantly decreased, the expression levels of essential proteins of MAPK signaling pathway such as JNK, P-JNK, ERK, P-ERK, P38, and P-P38 were significantly reduced; the expression levels of critical proteins in mTOR signaling pathways such as mTOR and P-mTOR were significantly decreased (Fig. 6). Molecular hypothesis pathway diagram is shown in Figure 7.

The key protein expression in PI3K-AKT/MAPK/mTOR signaling pathways were detected after transfection with LV-LINC00961 effect was verified at the protein levels.
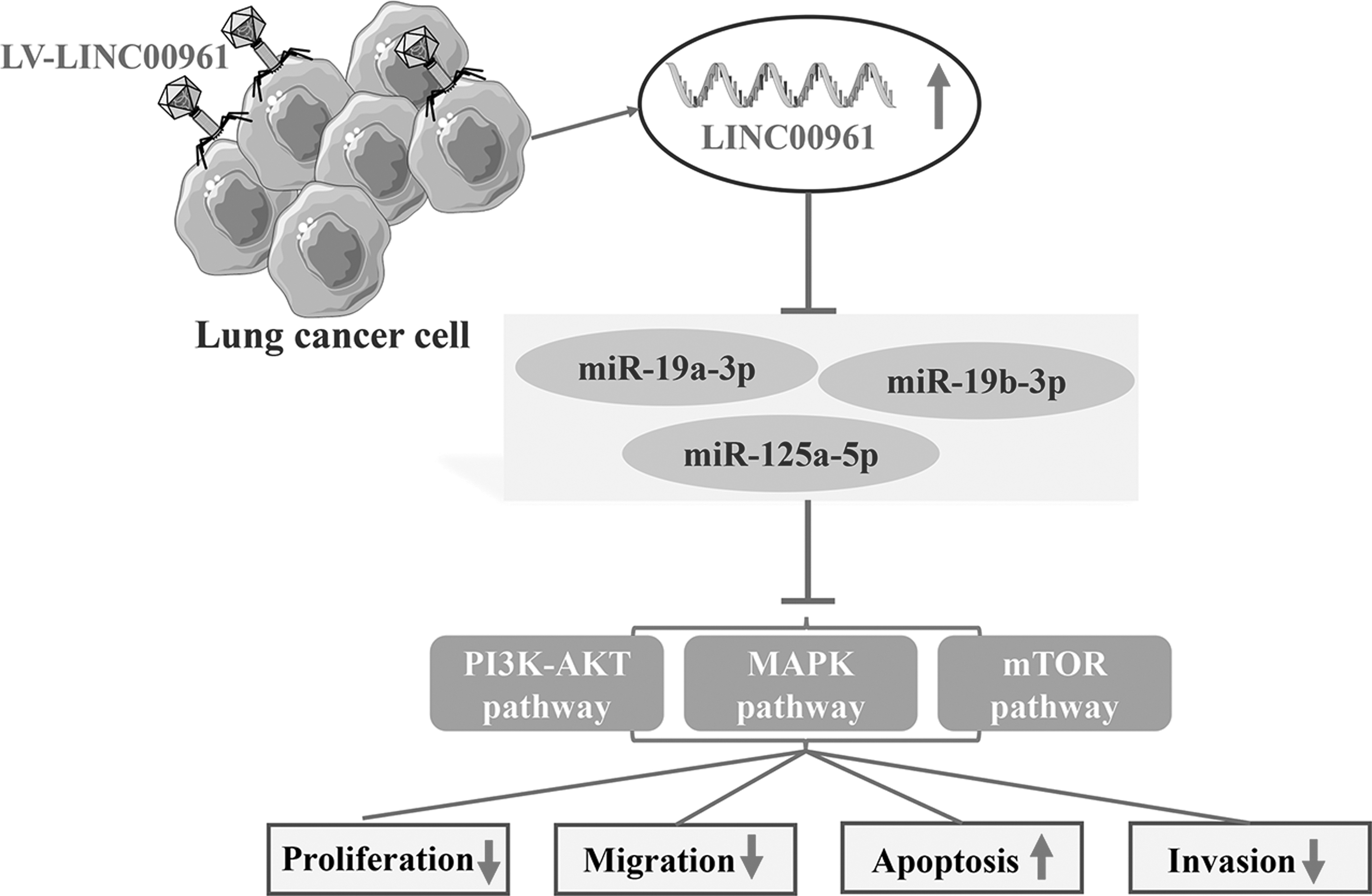
Molecular hypothesis pathway diagram in vitro.
LINC00961 overexpression inhibited tumor growth in vivo
A nude mice model was used to detect the potential biological process on tumor growth in vivo, the LINC00961 stably expressed A549 cells were subcutaneously injected into nude mice. The results demonstrated that, compared with the Negative control group, overexpressed LINC00961 significantly suppressed the average tumor volume (Fig. 8A) and weight (Fig. 8B) in vivo. The expression levels of miR-19a-3p/miR-19b-3p/miR-125b-5p significantly decreased (Fig. 8C). The expression levels of key proteins of PI3K-AKT/MAPK/mTOR signaling pathway in xenografted tissues were detected by western blotting, and the results exhibited that the expression levels of PI3K, P-ERK, BCL-XL, and BCL-2 were significantly reduced in vivo compared with the Negative control group (Fig. 8D).

LINC00961 suppresses NSCLC cell growth in vivo.
In summary, these results revealed that LINC00961-overexpression inhibited tumor growth through the PI3K-AKT/MAPK/mTOR signaling pathway in vivo.
Discussion
NSCLC accounted for 85–90% of lung cancer diagnoses, and despite advanced therapy, the median survival of 5-year survival rates for metastatic NSCLC patients remains dismal (Kanaan et al., 2015). In recent years, the incidence and mortality of NSCLC are steadily rising in both males and females (Dubey et al., 2016). Therefore, the identification of effective biomarkers and the novel regulatory mechanism for NSCLC should keep more attention. Accumulating evidence implies that lncRNA participated in the tumorigenesis of various malignant human cancers, including NSCLC (Dai et al., 2018; Lei et al., 2018), as exemplified by H19 and HOTAIR, which were reported to regulate the progression and development of NSCLC (Chen et al., 2019c; Huang et al., 2018a; Yang et al., 2018; Zheng et al., 2019).
In the current study, LINC00961 was downregulated in NSCLC tissues and cells, consistent with previous reports (Huang et al., 2018b; Jiang et al., 2018). Moreover, overexpression of LINC00961 inhibited NSCLC cell proliferation, migration, and invasion but promoted cell apoptosis in vitro. LINC00961-overexpression dramatically suppressed tumor volume and weight in vivo. However, the exact role of LINC00961 and its underlying mechanism is still unclear. This study first investigated PI3K-AKT/MAPK/mTOR signaling pathways involved in NSCLC development regulated by LINC00961 via miR-19a-3p/miR-19b-3p/miR-125b-5p.
Recent studies have also reported that LINC00961 was downregulated as tumor suppressor lncRNA in various tumors. Wu et al. verified that LINC00961 was significantly downregulated in colon tissues and cultured colon cell lines and promoted migration and invasion in colon cancer cells (Wu et al., 2020). Pan et al. reported that upregulated LINC00961 significantly suppressed cell proliferation and stimulated cell apoptosis in oral squamous cell carcinoma (OSCC), suggesting tumor suppressor of LINC00961 in OSCC (Pan and Sun, 2019). In glioma, downexpressed LINC00961 was found to be a worse predictor for overall survival, while upregulated LINC00961 markedly inhibited cell proliferation, migration, and invasion in vitro (Lu et al., 2018).
In the present study, we found LINC00961 was significantly downregulated in NSCLC tissues and cell lines. Moreover, overexpressed LINC00961 markedly suppressed the progression of NSCLC in vivo and in vitro, which was consistent with the previous studies (Huang et al., 2018b; Jiang et al., 2018).
Few studies on the function of LINC00961 in NSCLC have been reported, there is still no elaboration on the potential mechanism of LINC00961 involved in the occurrence of NSCLC. A competing endogenous RNA (ceRNA) network functioned as a novel regulating mechanism has been identified, in which lncRNA functions as a molecular sponge to adsorb miRNA, consequently suppressing miRNA target genes (Salmena et al., 2011). Accumulating evidence implies that a lncRNA could modulate more than one miRNA in various cancers. PVT1 negatively regulated the expression of miR-497 in NSCLC cells (Guo et al., 2018), miR-424 in ovarian cancer cells (Chen et al., 2018a), miR-128-3p in glioma cells (Fu et al., 2018), and miR-143 in gallbladder cancer cells (Chen et al., 2019b).
In addition, H19 might function as a ceRNA for miR-29b-3p and relieve the suppression of DNMT3B, which increased cell proliferation, invasion, and migration and promoted metastasis and EMT in bladder cancer (Lv et al., 2017). Moreover, previous studies have reported that LINC00961 inhibits proliferation, migration, and invasion via sponging miR-367 in melanoma (Mu et al., 2019), miR-223-3p in colon cancer (Wu et al., 2020), and miR-5581-3p in hepatocellular carcinoma (Yin et al., 2019).
In the present study, we first provided that LINC00961 sponge miR-19a-3p/miR-19b-3p/miR-125b-5p by a luciferase reporter. Furthermore, previous studies have suggested that suppressing miR-19b-3p exhibited inhibitory effects on cell proliferation, migration, and invasion in NSCLC cell lines (Wang et al., 2019). miR-125b-5p was upregulated in LUAC tissues/cells, inhibiting miR-125b-5p might suppress LUAC development (Zhang et al., 2020).
Our study confirmed that overexpressed LINC00961 suppressed the expression level of miR-19a-3p/miR-19b-3p/miR-125b-5p and upregulated miR-19a-3p/miR-19b-3p/miR-125b-5p overexpression reversed cell viability, invasion in A549 cells. Moreover, we found that upregulation of LINC00961 inhibited cell proliferation, migration, and invasion and promoted cell apoptosis by targeting PI3K-AKT/MAPK/mTOR signaling pathways. It indicated that LINC00961 might function as a ceRNA for miR-19a-3p/miR-19b-3p/miR-125b-5p to regulate PI3K-AKT/MAPK/mTOR signaling pathway to affect the progression of NSCLC.
Conclusions
In conclusion, our present study first investigated the underlying mechanism of LINC00961 in regulating the progression of NSCLC. The results indicated upregulated LINC00961 inhibited NSCLC cell proliferation, migration, and invasion and promoted cell apoptosis, potentially via suppressing the expression of miR-19a-3p/miR-19b-3p/miR-125b-5p by targeting PI3K-AKT/MAPK/mTOR signaling pathways. Taken together, our study provides a new perspective that LINC00961 acts as a tumor suppressor in NSCLC progression and a novel diagnostic and prognostic biomarker and therapeutic target for NSCLC. However, there are still many underlying mechanisms of LINC00961 that should be further investigated in NSCLC.
Footnotes
Authors' Contributions
Conceptualization, J.S., G.L.; Methodology, J.S.; Software, J.S.; Writing—original draft, J.S.; Writing—review and editing, Q.Z. and Y.Z. All authors have read and agreed to the published version of the article.
Acknowledgments
Part of this research is based on data generated by the TCGA and GEO database, these data are also available in TCGA and GEO.
Disclosure Statement
The authors declare no financial interest in the findings described in this study.
Funding Information
This study was supported by the National Natural Science Foundation of China (No. 81803201, 81673132, 81472939), the National Natural Science Foundation of China Youth Science Fund Project (81803201), the Startup Foundation for Introducing Talent of NUIST (No. 2020r088), and the Innovation and Entrepreneurship Program of Jiangsu Province (JSSCBS20210472)
Supplementary Material
Supplementary Data S1
References
Supplementary Material
Please find the following supplemental material available below.
For Open Access articles published under a Creative Commons License, all supplemental material carries the same license as the article it is associated with.
For non-Open Access articles published, all supplemental material carries a non-exclusive license, and permission requests for re-use of supplemental material or any part of supplemental material shall be sent directly to the copyright owner as specified in the copyright notice associated with the article.