
Abstract
Endometriosis is a benign gynecological disease sharing several features with malignant tumor. Cytoplasmic polyadenylation element-binding protein 3 (CPEB3), a potential target of miR-21-5p, is downregulated in endometriotic specimens. However, the function of CPEB3 in endometriosis is elusive. In this study, in cultured primary human endometrial stromal cells (ESCs), the overexpression and inhibition of CPEB3 were achieved by transduction of adenovirus-mediated CPEB3 overexpressed plasmid and shRNA, respectively. Functional analysis uncovered that upregulated CPEB3 reduced cell viability and arrested cell cycle entry. The expression of cyclin D1 and c-Myc was decreased after CPEB3 overexpression. Overexpression of CPEB3 facilitated ESC apoptotic potential, accompanied by increased Bax, cleaved-caspase 3 and cleaved-caspase 9, and reduced Bcl2. Moreover, elevated CPEB3 weakened migration and invasion abilities of ESCs. CPEB3 overexpression also reduced the expression of fibronectin and vimentin and the activities of matrix metalloproteinase (MMP)-9 and MMP-2. Interestingly, these effects were counteracted by CPEB3 inhibition. Furthermore, CPEB3 controlled the protein level of CXCL12, a homeostatic chemokine. CXCL12 elevation partially reversed the effects of CPEB3 on inhibiting ESC proliferation, migration and invasion, and promoting apoptosis. Based on these findings, it seems possible that CPEB3, as a critical player, attenuated the progression of endometriosis through repressing CXCL12 expression.
Introduction
Endometriosis is an estrogen-dependent gynecological disorder, predominantly defined as the presence and growth of endometrial glands and stroma outside the uterine cavity (Burney and Giudice, 2012; Vercellini et al., 2014). This disease is considered to be a chronic system disease that can cause chronic pelvic pain, lesion progression, and infertility, affecting ∼5–10% of reproductive-age women (Giudice and Kao, 2004; Taylor et al., 2021). Endometriosis is associated with a 50% increase in the risk of ovarian cancer (Vercellini et al., 2014).
Of note, it is characterized by infiltration of endometrial tissue and exhibits abnormal cellular proliferation and invasion, similar to malignant disease (Giudice et al., 1998; Chapron et al., 2019). Currently available treatments for endometriosis are hormones and surgery that results in undesirable side effects and are associated with high recurrence rates (Macer and Taylor, 2012; Vercellini et al., 2014). Its pathological mechanism remains poorly understood to date. Therefore, it is essential to unravel the molecular and cellular mechanisms linking the pathogenesis of endometriosis for exploring effective therapies.
A growing body of evidence indicates that dysregulation of mRNA is implicated in the development of endometriosis (Cai et al., 2019; Wu et al., 2020). Cytoplasmic polyadenylation element-binding proteins (CPEB1-4) are all RNA-binding proteins that activate or inhibit target specific mRNAs to modulate various biological behaviors (McGrew et al., 1989; Hake and Richter, 1994; Ford et al., 2019). Abnormal downregulated expression of CPEB3 has been observed in cervical and ovarian cancer, two types of common cancer in women (Hansen et al., 2009).
Previous researches have reported that CPEB3 functions as a suppressor of cell proliferation, migration, and invasion in cervical and ovarian cancer (Liu et al., 2019; Fang et al., 2020; Zhang et al., 2020b). CPEB3 also represses metadherin mRNA translation and reduces epithelial–mesenchymal transition and cell metastasis in hepatocellular carcinoma (Zhang et al., 2020a). Crispi et al.'s transcriptional profiling study uncovered decreased CPEB3 expression in ectopic endometrium (Crispi et al., 2013). However, little is known about the role of CPEB3 in endometriosis.
Chemoattractant cytokine C-X-C motif chemokine ligand 12 (CXCL12), also known as stromal-derived factor 1, is widely expressed in various human tissues, including lymph nodes, lung, stromal and endothelial cells, and plays an important role in a variety of biological processes (Zhou et al., 2019). Endometriosis produces chemokine CXCL12, which attracts bone marrow cells to the lesions (Moridi et al., 2017) and promotes cell proliferation, migration, and invasion (Ruiz et al., 2018). Based on these data, this study intended to investigate the function of CPEB3 in behavior of endometriosis-derived stromal cells and its possible mechanism linking CXCL12.
Materials and Methods
Clinical specimens
All volunteers provided informed consent form before recruitment, and the research protocol was approved by the Ethics Commission of Harbin Medical University (202106). In our study, ectopic endometrial tissues were obtained from five patients with ovarian endometriosis. As a control, normal endometrial samples were acquired from five healthy women undergoing laparoscopy or laparotomy. The clinicopathological characteristics enrolled in our study are shown in Supplementary Table S1. Staging of endometriosis was based on the widely accepted revised American Society for Reproductive Medicine classification (rASRM). All of the samples were histologically characterized and fixed for immunohistochemistry staining.
Immunohistochemistry
The fixed samples were paraffin embedded, dehydrated, and subjected to antigen retrieval. Then, the sections were incubated with 3% H2O2 for 15 min and blocked with 1% BSA for 15 min at room temperature. After overnight incubation at 4°C with primary antibody against CPEB3 (diluted 1:50; Abclonal, China), the sections were incubated with peroxidase-labeled goat anti-rabbit IgG secondary antibody (diluted 1:500; Thermo Fisher Scientific) for 1 h. After that, the reaction was developed with DAB, and they were lightly counterstained with hematoxylin (Solarbio, China). The images were observed under an Olympus microscope at 400 × magnification.
Isolation of endometrial stromal cells and treatment
Human primary endometrial stromal cells (ESCs) were isolated from ectopic endometrial tissues using the method previously described (Samartzis et al., 2019). Tissue samples were minced with scissors and digested enzymatically with collagenase solution at 37°C for 1.5 h. Stromal cells were collected by centrifugation and resuspended in DMEM (Servicebio, China) supplemented with 10% fetal calf serum at 37°C in a humidified atmosphere of 5% CO2. Before cellular experiments, the purity and homogeneity of ESCs were checked by immunofluorescence with antibodies against cytokeratin 7 (CK7, 1:50; Abcam, UK) and vimentin (1:50; Abclonal) as specific markers of epithelial and stromal cells, respectively.
Recombinant adenovirus infection was conducted according to the manufacturer's instructions. The purified ESCs were cultured in six-well plates and infected with adenovirus containing CPEB3 overexpression (oe-CPEB3) plasmid, CPEB3 shRNA, or their negative controls (vector and NC shRNA) at 37°C. After 48 h of infection, the efficiency of transduction was verified by western blotting assay. For evaluation of mRNA stability, the cells were treated with cycloheximide (5 mg/mL; MCE) for 1, 2, and 4 h on ice in the presence of oe-CPEB3.
Quantitative real-time PCR analysis
Total RNA was isolated using TRIpure reagent (BioTeke, China). Single-stranded cDNA was synthesized using the BeyoRT II M-MLV reverse transcriptase (Beyotime, China). Quantitative real-time PCR (qRT-PCR) was performed using SYBR Green, 2 × Taq PCR MasterMix (Solarbio) and specific primers synthesized by GenScript (Nanjing, China). The primer pairs for CXCL12 were as follows: 5′-CCAAACTGTGCCCTTCA-3′ (forward) and 5′-CTCTTCTTCTGTCGCTTCT-3′ (reverse).
Western blotting analysis
Total protein extracted from ESCs was quantified by a BCA protein quantitative analysis kit (Beyotime). The protein samples mixed with 5 × loading buffer were boiled for 5 min and loaded on a 10% sodium dodecyl sulfate-polyacrylamide gel electrophoresis (SDS-PAGE), and then transferred onto polyvinylidene difluoride membranes (Thermo Fisher Scientific). After sealing with 5% (M/V) BSA for 1 h, the membranes were incubated overnight with the corresponding primary antibodies at 4°C. Subsequently, they were subjected to HRP-labeled secondary antibodies at 37°C for 40 min. The signal was visualized using enhanced chemiluminescence (ECL)-western blotting detecting reagent (7 Sea biotech, China). The characteristics of primary antibodies are listed in Table 1.
The Primary Antibodies Used in Western Blotting Analysis
Methylthiazoletetrazolium assay
Cell viability was evaluated using a methylthiazoletetrazolium (MTT) assay kit (KeyGen, China) according to the manufacturer's instructions. In brief, ESCs were seeded in 96-well plates at 3 × 103 cells per well. The cells were incubated with 20 μL MTT solution for 4 h, and then treated with DMSO for 10 min in darkness. Finally, the optical absorbance at 490 nm was recorded.
Cell cycle analysis
The primary ESCs were harvested by centrifugation at 300 g for 5 min. After that, 500 μL staining buffer was added to resuspend the cells, and they were incubated with 25 μL propidium iodide staining solution at 37°C for 30 min in the dark. The distribution of cell cycle was analyzed by flow cytometry.
Terminal deoxynucleotidyl transferase-mediated dUTP nick-end labeling assay
Terminal deoxynucleotidyl transferase-mediated dUTP nick-end labeling (TUNEL) assay was carried out based on an In Situ Cell Death Detection Kit (Roche, Germany). Cultured ESCs were incubated in 0.1% Triton X–100 permeabilization solution (Beyotime) at 37°C for 15 min. After washing with phosphate-buffered saline for three times, the samples were incubated with TUNEL reaction mixture for 1 h. The TUNEL-positive cells were observed under a fluorescence microscope.
Apoptosis analysis
Apoptosis of ESCs was evaluated in accordance with the manufacturer's protocols (KeyGen). When the ESCs reached subconfluence, they were harvested by centrifugation. Then, 7-amino-actinomycin D (7-AAD) was added and reacted in the dark for 10 min. Before being analyzed by flow cytometry, the cells were incubated with 1 μL Annexin V-PE for 10 min at room temperature.
Wound healing assay
A 200 μL pipette tip was used to scrape a line, and loose cells were removed. The cells were then cultured in serum-free DMEM for 24 and 48 h, and photographed under a microscope at 100 × magnification. The percentage of cell migration was calculated according to the method of Lu et al. (2021).
Transwell assay
When cells reached 90% confluency, the medium was removed and the cells were digested with 0.25% trypsin. The lower chamber was filled with media containing 10% fetal calf serum, and 3 × 104 cells per well were inoculated into the upper chamber. After 48 h of incubation, the cells were fixed with 4% paraformaldehyde for 20 min and dyed with 0.5% crystal violet for 5 min. The stained cells were observed under an Olympus microscope at 200 × magnification. The number of invasive cells was counted in five random fields of each chamber.
Gelatin zymography assay
Gelatin zymography assay was performed to assess the activities of matrix metalloproteinase (MMP)-2 and MMP-9. The supernatant of ESCs was collected and dissolved on 10% SDS-PAGE containing gelatin (Sigma). The gels were then washed with TritonX-100 and incubated with reaction buffer at 37°C for 40 h. After Coomassie brilliant blue staining for 3 h, the gels were destained and the intensity of bands was determined by gel documentation system.
RNA immunoprecipitation assay
HEK293 cells were obtained from Shanghai Zhong Qiao Xin Zhou Biotechnology Co., Ltd. RIP assay was performed using EZ-Magna kit (Millipore). After being lysed in specific lysis buffer, the cells were centrifuged for 10 min at 14,000 rpm. The supernatant was collected and incubated at 4°C with anti-CPEB3 antibody for 3 h for coprecipitation. The magnetic bead-antibody complex was resuspended in 0.5 mL RNA immunoprecipitation (RIP) wash buffer. The immuneprecipitated RNA was isolated and reversed, followed by PCR analysis.
Statistical analysis
All experiments were performed in triplicate at least. Data were processed using the GraphPad 8.0 software and presented as mean ± standard deviation. Comparisons between the two groups were evaluated using Student's unpaired two-tailed t-test. Differences between multiple groups were assessed using one-way analysis of variance (ANOVA). Data between groups at different time points were analyzed using two-way ANOVA, followed by Tukey's multiple comparisons test. Statistical significance was set at p < 0.05.
Results
CPEB3 expression is abnormally downregulated in endometriosis
To clarify the expression pattern of CPEB3 and CXCL12 in endometriosis, we first analyzed the transcription profile of selected endometriosis (GSE25628) from open NCBI GEO DataSets (

CPEB3 expression was reduced in endometrium from patients with endometriosis.
Furthermore, primary ESCs were isolated from human ectopic endometrial tissues and confirmed by immunofluorescence staining. It was demonstrated that vimentin was strongly expressed in isolated cells, whereas the specific marker for epithelial cells CK7 was almost unexpressed (Fig. 1D), indicating that primary ESCs were successfully obtained in vitro.
CPEB3 suppresses the proliferation of ESCs
To explore the role of CPEB3 in primary ESCs, we overexpress or suppress it followed by functional assays. As shown in Figure 2A, the transduction efficiency of CPEB3 overexpression or knockdown was confirmed. MTT assay revealed that cell viability was inhibited after CPEB3 overexpression (Fig. 2B). Overexpression of CPEB3 also delayed cell cycle entry (Fig. 2C and Supplementary Fig. S1A). Western blotting analysis showed that CPEB3 overexpression reduced the expression of cyclin D1 and c-Myc (Fig. 2D). Of note, these effects were diminished after CPEB3 knockdown (Fig. 2).
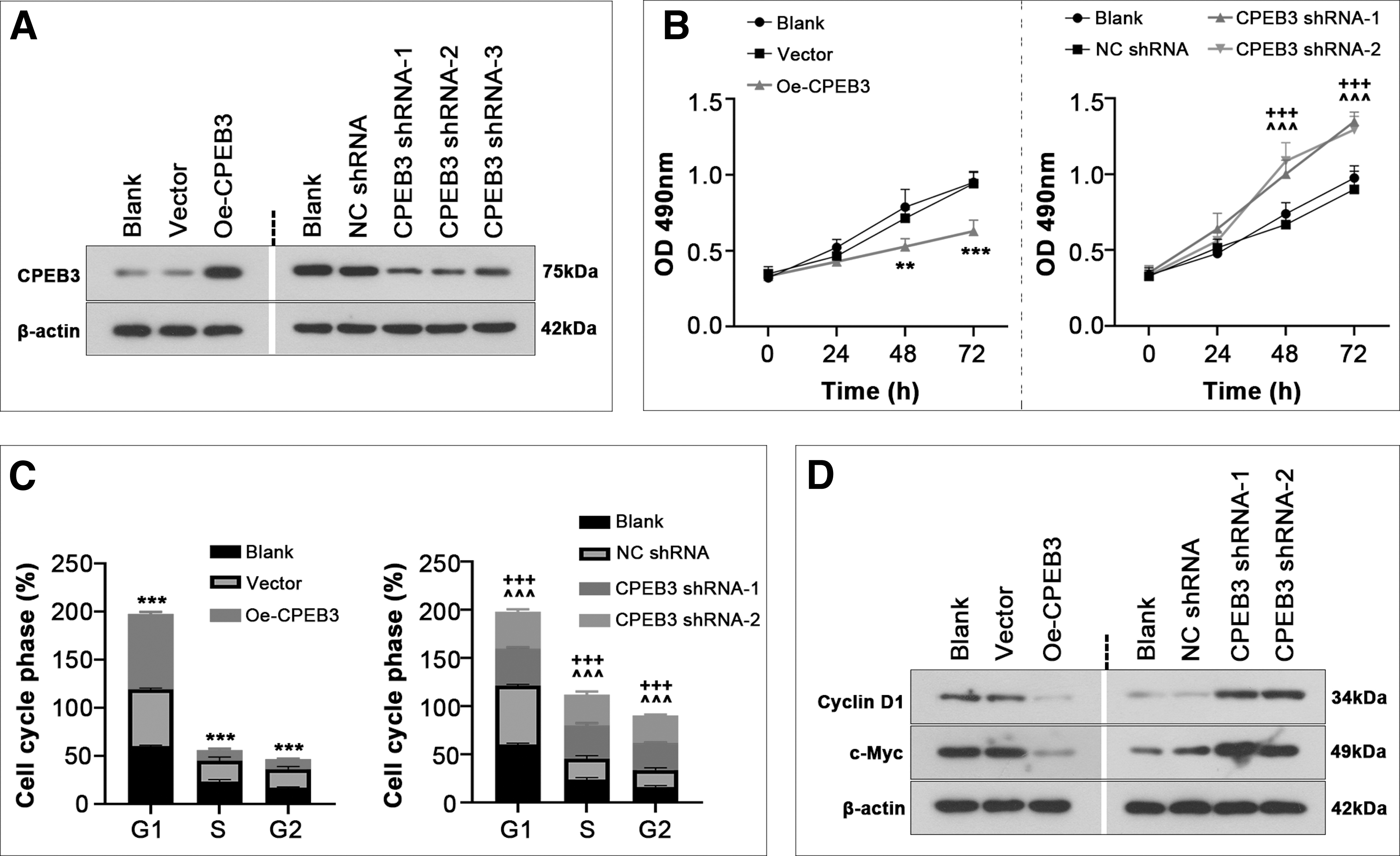
CPEB3 suppressed proliferation of ESCs. Primary human ESCs were infected with adenovirus containing CPEB3 overexpression plasmid or its shRNA for 48 h (n = 3).
CPEB3 potentiates the apoptosis of ESCs
To find out the effect of CPEB3 on apoptosis of ESCs, TUNEL staining and flow cytometry were used. The results demonstrated that CPEB3 overexpression increased apoptosis, which was reduced by CPEB3 inhibition (Fig. 3A, B and Supplementary Fig. S1B). Moreover, CPEB3 overexpression increased the expression levels of apoptosis-related proteins Bax, cleaved-caspase 3 and cleaved-caspase 9, and reduced Bcl2 (Fig. 3C). However, CPEB3 inhibition suppressed the apoptotic potential of ESCs (Fig. 3).
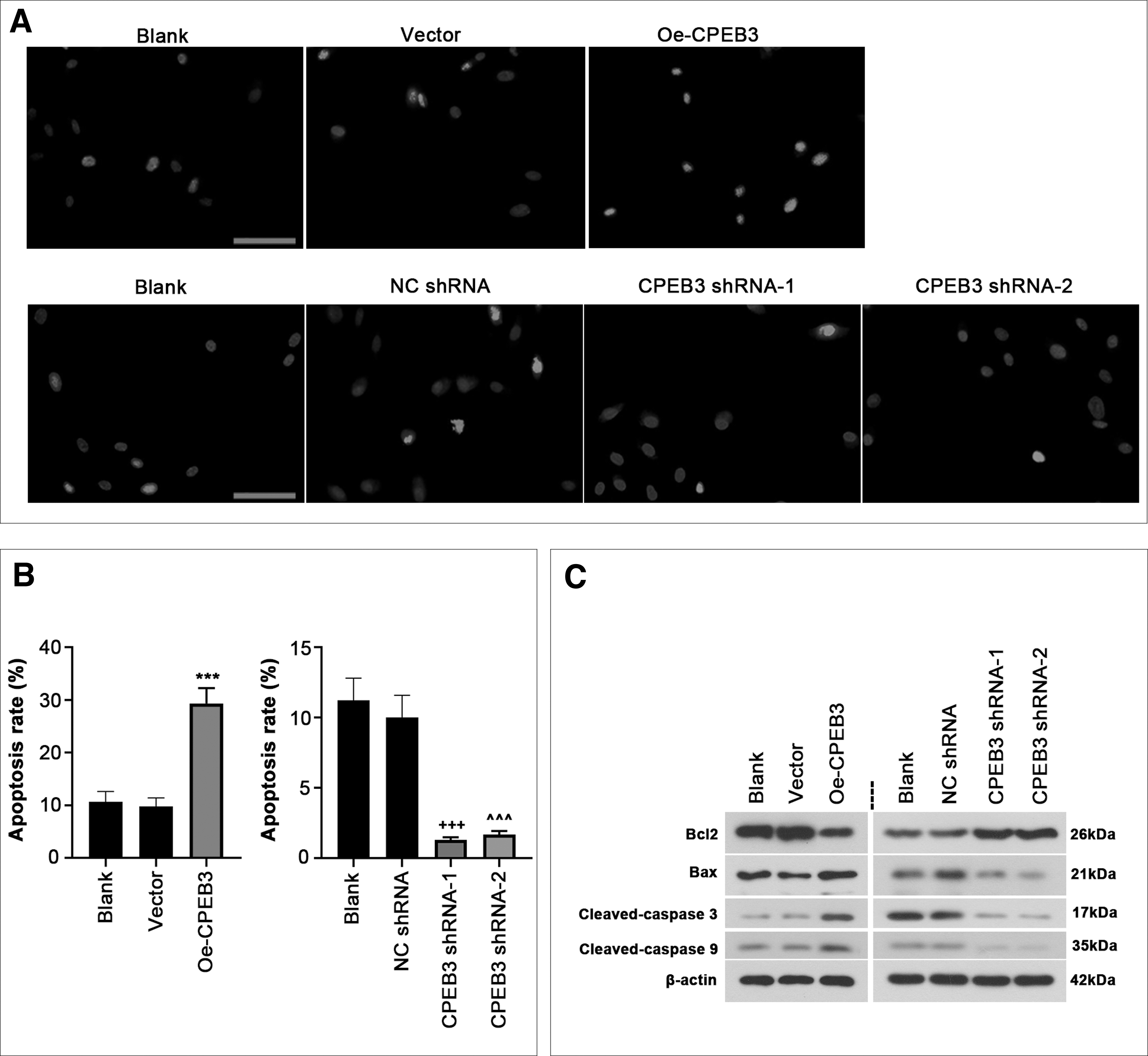
CPEB3 promoted cell apoptosis.
CPEB3 inhibits the migration and invasion of ESCs
To further determine whether CPEB3 affects migration and invasion of ESCs, wound healing and transwell assays were conducted. As exhibited in Figure 4A–D, CPEB3 overexpression suppressed the migration and invasion capabilities of ESCs, whereas the opposite effects were observed after CPEB3 inhibition. Besides, relative expression levels of vimentin and fibronectin were obviously decreased after CPEB3 overexpression, but enhanced after CPEB3 inhibition (Fig. 4E). We also observed that CPEB3 overexpression inhibited the activities of MMP2 and MMP9 by gelatin zymography assay, which was partially abolished by inhibition of CPEB3 (Fig. 4F).
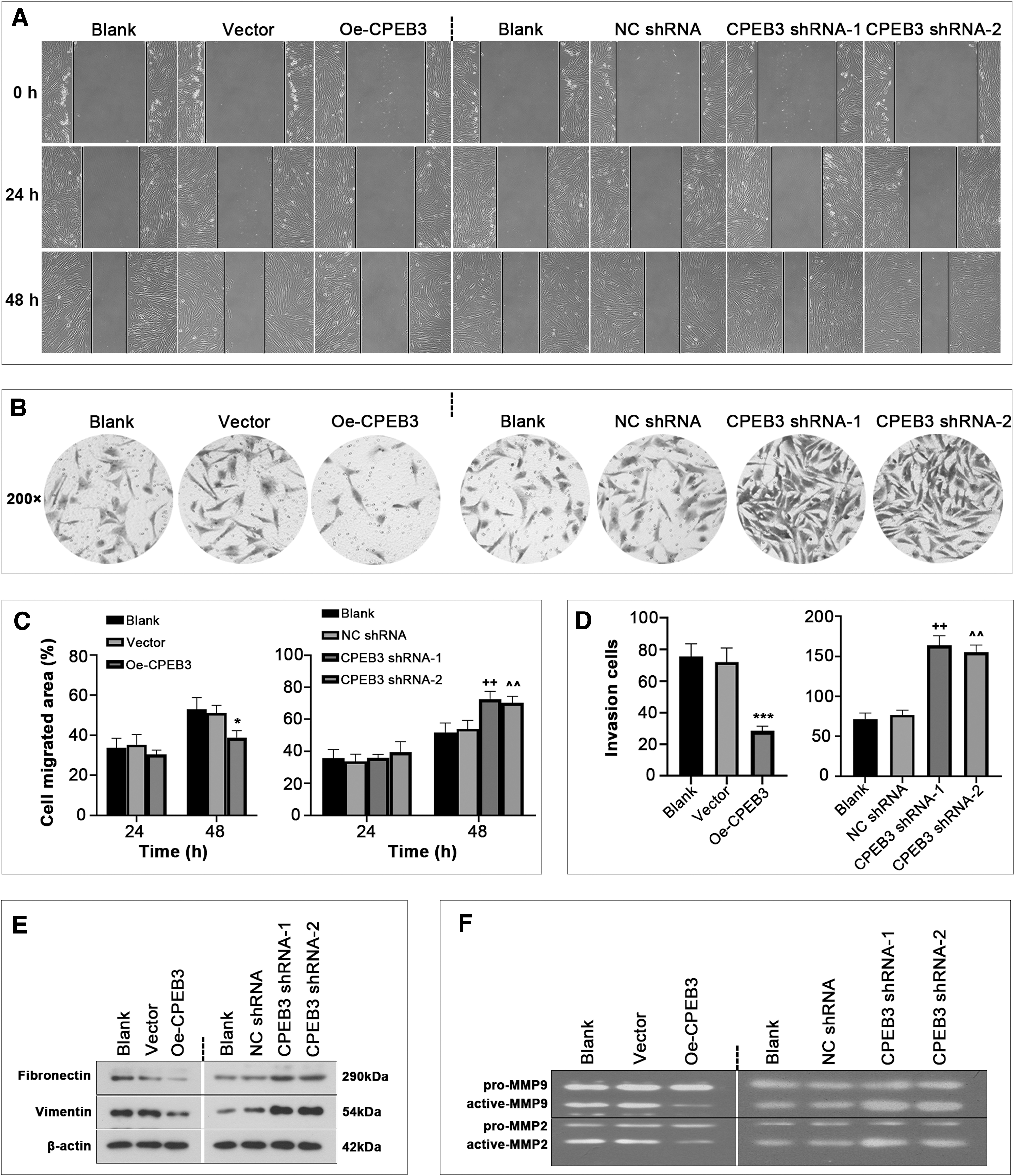
CPEB3 inhibits endometrial cell migration and invasion.
CPEB3 blocks CXCL12 expression
To gain insight into the molecular mechanisms related to CPEB3 in endometriosis, a stromal cell-derived chemokine CXCL12 was focused on. As depicted in Figure 5A–C, CPEB3 overexpression or inhibition did not obviously alter CXCL12 mRNA level and stability, and only affected CXCL12 protein level, suggesting that CPEB3 might modulate the translation of CXCL12 mRNA. Moreover, the result of RIP assay showed that CXCL12 signal was enriched in CPEB3 precipitated RNAs in HEK293 cells (Fig. 5D), further confirming that CXCL12 might be crucial for the function of CPEB3.

CPEB3 impacts CXCL12 expression.
CPEB3 impacts cell behavior of ESCs by regulating CXCL12
To clarify whether CPEB3 regulates cell behaviors by affecting CXCL12, CXCL12 was overexpressed in ESCs (Fig. 6A). Functional analysis showed that overexpression of CXCL12 rescued the decrease in cell viability caused by CPEB3 overexpression. The apoptosis-promoting effect of CPEB3 was partially offset by overexpression of CXCL12 (Fig. 6C). Overexpression of CXCL12 also reversed the inhibitory effect of CPEB3 on cell migration and invasion (Fig. 6D, E). In addition, CPEB3 expression was decreased after treatment of micoRNA (miR)-21-5p, a noncoding small RNA molecule (Supplementary Fig. S2A). The CPEB3's effects on ESC viability, apoptosis, and invasion were partially counteracted after miR-21-5p treatment (Supplementary Fig. S2B–D).

CXCL12 overexpression reversed the effect of CPEB3 on endometrial cell behavior.
Discussion
Although endometriosis is a gynecological disorder, not a malignant disease, it possesses malignant biological behavior, similar to tumor (Tosti et al., 2015). CPEB3, an RNA-binding protein, plays a key role in regulating cellular aggressive behavior (Richter, 2007; D'Ambrogio et al., 2013). It is found to be abnormally downregulated in endometriosis; however, its function and regulatory mechanisms in endometriosis remain largely unknown. This study demonstrated that CPEB3 could alter several cellular characteristics of endometriosis by modulating CXCL12. To our knowledge, this report is the first study to highlight the importance of CPEB3-CXCL12 signaling in endometriosis.
Initial evidence linking CPEB3 strongly proves its regulatory role in tumorigenesis (D'Ambrogio et al., 2013). CPEB3 expression is downregulated in ovarian, cervical, and colorectal tumors (Liu et al., 2019; Fang et al., 2020; Zhang et al., 2020b). Overexpressing CPEB3 can depress the migration and invasion of high-grade ovarian serous tumor (Liu et al., 2019). Inhibition of CPEB3 facilitates cell cycle entry and reduces the apoptosis rate of cumulus cells (Wang et al., 2020). Given that endometriosis shares striking characteristics with human tumors (He et al., 2018), we investigated the function of CPEB3 in several behaviors of endometrial cells.
As expected, aberrant downregulation of CPEB3 expression in ectopic endometrium was identified, indicating that CPEB3 may be a critical factor in the pathogenesis of endometriosis. Similar to previous studies, we found that upregulation of CPEB3 suppressed the proliferation, migration, and invasion of ectopic endometrial cells, and accelerated cell apoptosis. On the contrary, knockdown of CPEB3 resulted in opposite cellular behaviors. These data support the importance of CPEB3 in endometriosis. MiR-21-5p, a small regulatory oncogenic miRNA, has been reported to be obviously upregulated in the endometrium of patients with endometriosis and impact ESC apoptosis, which may be a therapeutic biomarker for endometriosis (Liu et al., 2018; Park et al., 2018; Moustafa et al., 2020).
Interestingly, CPEB3 has been identified as a putative downstream target of miR-21-5p (Stadnik et al., 2021). This prompted us to speculate that the function of CPEB3 in endometriosis may be mediated by miR-21-5p. In this study, we found that miR-21-5p treatment diminished the changes caused by CPEB3 overexpression in ESCs, which was consistent with our speculation. However, the regulatory mechanisms involving miR-21-5p and CPEB3 in endometriosis require further investigation.
RNA-binding proteins have recently been recognized to play vital roles in regulating gene expression and cellular physiology (Hentze et al., 2018). It is reported earlier that CPEB3 can bind to the U-rich regions to regulate the translation of indentified mRNA targets (Huang et al., 2006). Ford et al. (2019) reported that CPEB3 can be recruited and inhibits the translation of its target mRNAs, regulating the persistence of memory. Zhang et al. (2020a) uncovered that CPEB3 acts as a suppressor in liver tumor through suppressing MTDH mRNA translation.
However, the regulatory mechanism whereby CPEB3 affects ectopic endometrial cell behaviors remains unclear. CXCL12 is a chemokine that is commonly expressed at sites of tissue damage (Dotan et al., 2010). Ruiz et al. (2018) found that CXCL12 can impact normal reproductive biology and immune surveillance. CXCL12 also facilitates the proliferation, migration, and invasion of endometriotic cells (Leconte et al., 2014; Ruiz et al., 2018). Our present data demonstrated that CXCL12 was highly expressed in endometriosis lesions compared with eutopic endometrium, which was in agreement with a previous study (Leconte et al., 2014).
Moreover, we demonstrated that CPEB3 overexpression reduced CXCL12 protein level, whereas CXCL12 mRNA level and stability were unchanged. Further studies are required to validate whether CPEB3 affects CXCL12 RNA stability or production. A profound and persistent residues-dependent CPEB3 binding to CXCL12 mRNA was observed using RIP assay, further confirming the binding of CPEB3 and CXCL12 mRNA. Furthermore, CXCL12 overexpression promoted ESC proliferation, migration, and invasion, but inhibited apoptosis under CPEB3 overexpression. These results support that CPEB3 may act as a key regulator of CXCL12 and that CPEB3 may attenuate endometriotic lesions potentiality through inhibiting CXCL12 expression.
However, we have only evaluated and clarified the biological function of CPEB3 in primary ESCs. Obviously, an animal endometriosis model is clearly needed to further verify the role of CPEB3 in endometriosis. Besides, there are limited data in the literature on the function of the CPEB family in endometriosis, and further studies are required to elucidate the unknown mechanism of cross talk between CXCL12 and the CPEB protein families. In addition, CXCL12 can bind to its specific receptor CXCR4, which impacts immune evasion in endometriosis (Tal et al., 2021). CXCL12 has been reported to be involved in immune surveillance of endometriosis (Chen et al., 2021). Whether the CPEB3/CXCL12 axis affects immune evasion and surveillance needs to be studied further.
In conclusion, our study identifies the function of CPEB3 in ESCs. Overexpression of CPEB3 represses ESC proliferation, migration, and invasion, and facilitates apoptosis, at least in part through negative regulation of CXCL12, as illustrated in Figure 7. It is highlighted that targeting CPEB3 may be an attractive strategy to attenuate ectopic endometrial lesions.
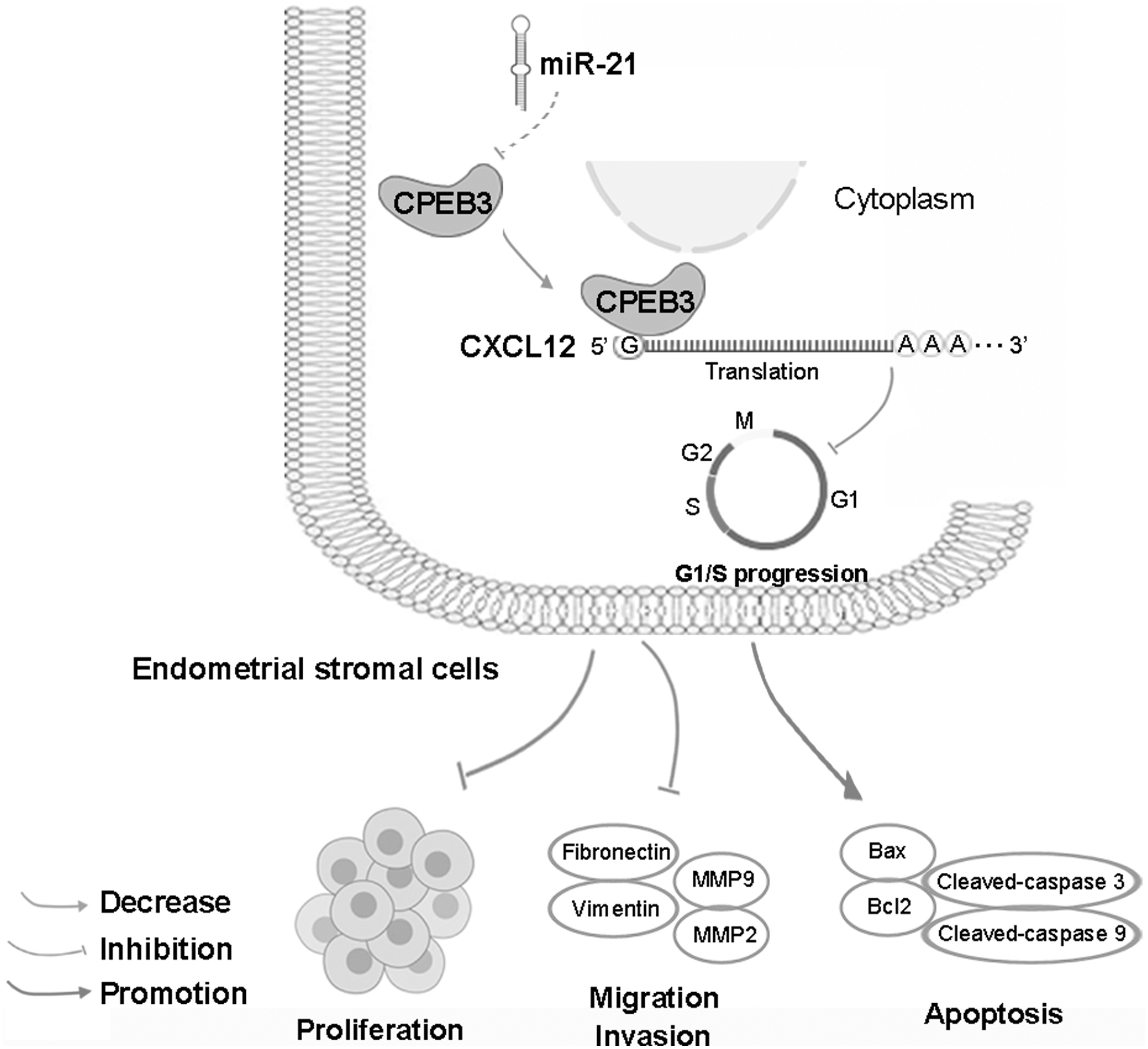
Graphical abstract. Overexpression of CPEB3 repressed ESC proliferation, migration, and invasion, and facilitates apoptosis that acted, at least in part, by negatively regulating CXCL12.
Data Availability Statement
The data supporting the findings of this study are available in this article.
Footnotes
Authors' Contributions
G.Z. and J.W. conceptualized the study and wrote the article. J.W., H.W., X.W., X.Z., and L.S. performed the experiments. Y.C., X.J., and J.L. analyzed the data. All authors have read and approved the final article.
Disclosure Statement
No competing financial interests exist.
Funding Information
This study was supported by the National Natural Science Foundation of China (81971359 and 82101725); the China Postdoctoral Science Foundation (2018M640308); the Key Research and Development Program of Heilongjiang (GA21C008); the Research and Development of Applied Technology of Harbin (2017AB9BS039).
Supplementary Material
Supplementary Table S1
Supplementary Figure S1
Supplementary Figure S2
References
Supplementary Material
Please find the following supplemental material available below.
For Open Access articles published under a Creative Commons License, all supplemental material carries the same license as the article it is associated with.
For non-Open Access articles published, all supplemental material carries a non-exclusive license, and permission requests for re-use of supplemental material or any part of supplemental material shall be sent directly to the copyright owner as specified in the copyright notice associated with the article.