
Abstract
Sepsis is a global health care issue that affects millions of people. DNA methyltransferase I (DNMT1)-mediated DNA methylation is involved in a number of human diseases by affecting many types of cellular progression events. However, the role and underlying molecular mechanism of DNMT1 in development of sepsis remain largely unknown. Lipopolysaccharide (LPS) induced lung fibrosis in the sepsis mouse model, and DNMT1 was upregulated in lung tissues of a sepsis mouse model compared with lung tissues from control mice. Then, this study demonstrated that LPS induced the production of interleukin (IL)-7 and tumor necrosis factor (TNF)-α and promoted DNMT1 expression in primary type II alveolar epithelial cells (AECII cells). Knockdown of DNMT1 inhibited IL-7 and TNF-α secretion in AECII cells exposed to LPS. Further study demonstrated that DNMT1 repressed the expression of miR-130a in AECII cells with or without LPS exposure. Next, this study demonstrated that miR-130a inhibited ZEB1 expression in AECII cells exposed to LPS. Ultimately, this study revealed the role of the DNMT1/miR-130a/ZEB1 regulatory pathway in AECII cells exposed to LPS. Overall, our data revealed that LPS induced the secretion of inflammatory factors by modulating the DNMT1/miR-130a/ZEB1 regulatory pathway in AECII cells, thus providing a novel theoretical basis that might be beneficial for establishment of diagnostic and therapeutic strategies for sepsis.
Introduction
In recent years, despite the significant advancements in medical services, sepsis is still an important cause of death in intensive care unit (ICU) patients (Genga and Russell, 2017). Sepsis is an immune disease caused by the immune system's overreaction to severe infections (Cecconi et al., 2018). The most common infection factor of sepsis is bacteria; other factors include parasites, viruses, and fungi in tissues, including in the urinary tract, lung, blood, and skin. Sepsis is considered a continuous process from infection to widespread organ dysfunction syndrome, such as acute liver injury, acute kidney injury, and acute lung injury (Mayer et al., 1995). Although sepsis has serious consequences, the mechanisms of occurrence and development of sepsis are not yet clear.
As an important epigenetic modification, DNA methylation refers to the covalent bonding of a methyl group to the 5th carbon position of the cytosine of the genomic CpG dinucleotide under the action of DNA methyltransferase (DNMT1) (Moore et al., 2013; Diomede et al., 2017; Ren et al., 2018). In mammals, DNA methyltransferases mainly include DNMT1, DNMT3a, and DNMT3b (Pizzicannella et al., 2021).
DNMT1 is mainly responsible for maintaining DNA methylation modification, and DNMT3a and DNMT3b are responsible for reinitializing DNA methylation modification (Parashar et al., 2018). In the state of high DNA methylation modification, gene expression usually manifests as transcriptional inhibition, while in contrast, hypomethylation manifests as increased gene expression (Consortium, 2012; Argentieri et al., 2017).
DNMT1-mediated DNA methylation is involved in a number of human diseases by affecting many types of cellular processes, including cell growth, cell cycle progression, metastasis, apoptosis, development, and tumorigenesis (Bird, 2002; Jones, 2012; Argentieri et al., 2017). Several reports have documented that DNMT1 is increased in sepsis (Robertson, 2005; Dhas et al., 2015). Nevertheless, the role of DNA methylation modification in sepsis and its molecular mechanism require further study.
MicroRNAs (miRNAs) are a class of endogenous small RNAs with lengths of ∼20–24 nucleotides (Hammond, 2015). miRNAs play a variety of important regulatory roles in various cellular processes by regulating the numbers of genes through binding with response elements on the mRNAs of genes, leading to alterations of mRNA stability and translation efficiency (Saliminejad et al., 2018).
A variety of miRNAs are aberrantly expressed and exert an important influence on the physiological and pathological processes of sepsis (Kingsley and Bhat, 2017). Catalin Vasilescu et al. (2009) showed that miR-150 could serve as a plasma prognostic marker in patients undergoing sepsis. Cui et al. (2016) showed decreased expression of miR-130a in severe sepsis patients. Nevertheless, the regulatory and functional mechanisms of miR-130a in sepsis need to be further studied.
This study aimed to investigate the role of DNMT1 and to further elucidate the functional molecular mechanism in sepsis. This study showed that increased DNMT1 elevated the stage of DNA methylation modification of the miR-130a promoter and thus led to decreased expression of miR-130a in lipopolysaccharide (LPS)-induced sepsis mouse and cell models, providing a novel mechanism of DNMT1-mediated DNA methylation in the development of sepsis.
Methods
Murine model of sepsis
Seven- to 8-week-old, male C57BL/6 mice were obtained from Shanghai SLAC Laboratory Animal Co., Ltd. (Shanghai, China), and housed in an SPF room at Shanghai Pulmonary Hospital, and the experimental procedure was conducted under the supervision of the Animal Care Committee of Shanghai Pulmonary Hospital according to the Guide for the Care and Use of Laboratory Animals (Ethic number: K21–242).
Ten mice were randomly divided into two groups: the sham group and LPS group, each with five mice. For the LPS group, mice were intraperitoneally injected with 5 mg/kg LPS (Lianmai Biological Engineering, Shanghai, China) for 12 h; the sham group was intraperitoneally injected with phosphate-buffered saline (PBS). HE staining was performed by Wuhan Servicebio Biological Technology Co., Ltd. (Wuhan, China).
Cell culture
AECII cells were purchased from Suzhou Junxin Biological Technology Co., Ltd. (Junxin Biotech, Suzhou, China), and were routinely maintained in our laboratory. Then, AECII cells were maintained in Dulbecco's modified Eagle's medium (DMEM; Sangon, Shanghai, China) containing 10% fetal bovine serum (Quacell, Zhongshan, Guangzhou), 100 U/mL penicillin, and 100 mg/mL streptomycin in a humidified incubator containing 95% air and 5% CO2 at 37°C.
Characterization of AECII cells by immunofluorescence staining assay
The purity of AECII cells was verified using immunofluorescence staining analysis. After seeding onto collagen-coated coverslips, the AECII cells were fixed with paraformaldehyde, permeabilized with acetone/methanol, and blocked with animal serum. Then, AECII cells were incubated with the anti-CK-18 antibody at a dilution of 1:200, followed by secondary antibodies at a dilution of 1:1000.
After staining of nuclei with Hoechst, the cell images were captured under a fluorescence microscope (Leica, Germany) with at least three random fields
Plasmid construction, miRNAs, and cell transfection
Ectopic and silenced gene expression systems were constructed based on the lentivirus system. The cDNA of DNMT1 was cloned into the pHIV-puro vector, and short hairpin RNAs (shRNAs) against DNMT1 (shRNA-DNMT1) were inserted into the pLKO-puro vectors. Small RNAs, including miR-130a mimics, miR-130a inhibitor, and corresponding control RNAs, were purchased from Shanghai GenePharma Technology Co., Ltd. (Shanghai, China).
All plasmids and small RNAs were transfected into the cells using the highly efficient transfection solution purchased from Suzhou Junxin Biotechnology Co., Ltd.
Gene expression analysis by quantitative real-time PCR
For RNA preparation, the tissues were ground into cell fluid with a glass grinder, and cells were washed with precooled PBS. RNA extraction solution (Suzhou, Junxin, China) was employed to isolate the total RNA, and a Reverse Transcription Kit (Tiangen Biotech, Beijing, China) was used to perform cDNA synthesis based on the manufacturer's protocol.
SYBR Green I was applied to carry out the quantitative real-time PCR (qPCR) analysis on an ABI-Prism 7500 real-time quantitative PCR system (ABI). Gene expression was normalized to the internal reference gene, β-actin. Differences in the genes were calculated with the 2Ct(candidate)/2Ct(control) formula.
The following primers were used: forward primer of DNMT1: TCCGGAAGAGCCATGTGAAC; reverse primer of DNMT1: GTCTCTGTCCTCGGGAGTCT; forward primer of ZEB1: ACAAGCGAGAGGATCATGGC; reverse primer of ZEB1: TGCATCTGAGTCACACTCGT; forward primer of β-actin: GGTACCACCATGTACCCAGG; and reverse primer of β-actin: GGTGTAAAACGCAGCTCAGTAA.
Gene expression analysis by Western blot
For protein preparation, the tissues were ground into cell fluid with a glass grinder, and cells were washed with precooled PBS. Proteins were extracted using RIPA buffer, and the concentrations were measured using the Coomassie brilliant blue method. Protein samples were resolved by 10% sodium dodecyl sulfate–polyacrylamide gel electrophoresis (SDS-PAGE), followed by electrophoretic transfer onto a polyvinylidene fluoride membrane (Millipore).
Protein bands were probed with the anti-DNMT1 antibody (1:1000; Abcam, Cambridge, UK), anti-ZEB1 antibody (1:1000; Abcam), or anti-β-actin antibody (Abcam), followed by corresponding secondary antibodies conjugated with horseradish peroxidase (1:4000; Abcam). Antigenic bands were detected using enhanced chemiluminescence (ECL; Junxin, Suzhou, China). β-Actin served as an internal reference gene.
Cytokine production analysis by enzyme-linked immunosorbent assay
The supernatants were harvested, centrifuged (10 min, 1500 g) at 1500 g/min for 5 min at 4°C, and stored at −80°C. Concentrations of interleukin (IL)-7 and tumor necrosis factor (TNF)-α were measured with ELISA kits (Sigma) following the instructions of the manufacturer.
Statistical analysis
The results of experiments repeated three times are represented as the mean ± standard deviation. Student's t test was used to determine the variance between two samples using Microsoft Excel software. One-way analysis of variance was employed for comparisons among multiple groups using Prism 5.0 (San Diego, CA). p-Values <0.05 were regarded as statistically significant.
Results
Increased expression of DNMT1 in lung tissue of an LPS-induced sepsis mouse model
As shown in results of the HE analysis, LPS induced lung fibrosis in the sepsis mouse model (Fig. 1A). Furthermore, secretion of IL-7 and TNF-α was increased in the LPS-induced sepsis mouse model based on results of the enzyme-linked immunosorbent assay (ELISA) analysis (Fig. 1B).

Increased expression of DNMT1 in lung tissues of an LPS-induced sepsis mouse model.
Further study showed that DNMT1 expression was upregulated in the LPS-induced sepsis mouse model based on the results of qPCR and Western blot analysis (Fig. 1C, D), indicating that DNMT1 expression might be involved in development of sepsis.
Downregulation of DNMT1 inhibited the role of LPS in AECII cells
As shown in results of the immunofluorescence staining analysis, the isolated AECII cells were positive for cytokeratin-18 (Fig. 2A), indicating the high purity of AECII cells used in this study. After exposure to LPS (0, 5, 10, 25, and 50 μg/mL) for 48 h, secretion of IL-7 and TNF-α was increased in AECII cells (Fig. 2B). As shown by the results of qPCR and Western blot analysis, mRNA and protein levels of DNMT1 were increased in AECII cells exposed to different concentrations of LPS (Fig. 2C).
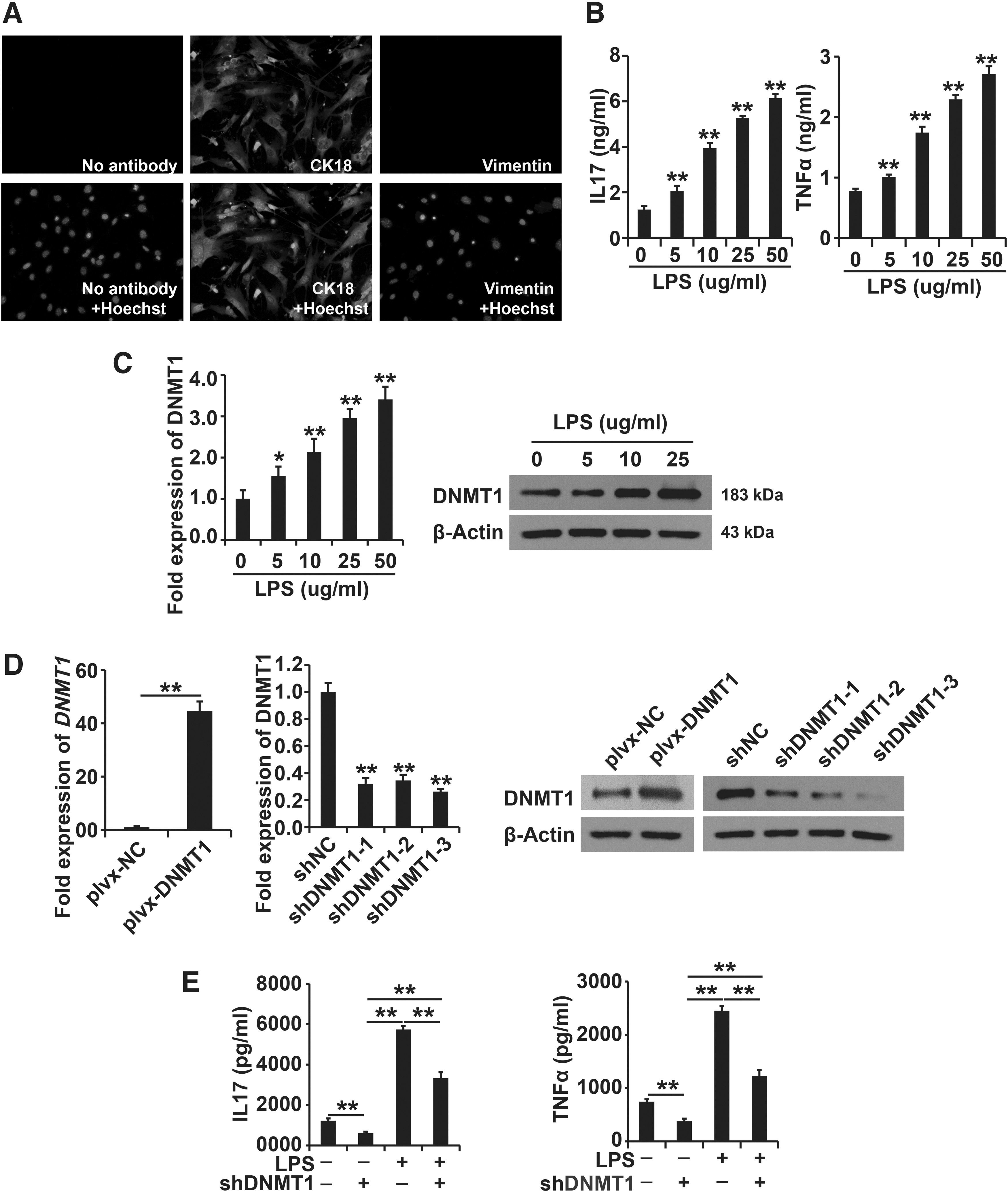
Downregulation of DNMT1 inhibited the role of LPS in AECII cells.
The upregulation and downregulation of the DNMT1 system were successfully constructed (Fig. 2D). After knockdown of DNMT1 expression, AECII cells were treated with LPS (50 μg/mL) for 48 h, and secretion of IL-7 and TNF-α was determined using ELISA. Downregulated expression of DNMT1 decreased IL-7 and TNF-α production in AECII cells with or without LPS exposure (Fig. 2E). In short, our study proved that ectopic expression of DNMT1 repressed the LPS-induced cellular progression of AECII cells.
DNMT1 inhibited miR-130a expression in AECII cells with or without LPS exposure
miR-130a expression was significantly downregulated in sepsis. Based on the qPCR analysis data, miR-130a expression was decreased in the lung tissue of the LPS-induced sepsis mouse model (Fig. 3A). After exposure to LPS (0, 5, 10, 25, and 50 μg/mL) for 48 h, the expression of miR-130a in AECII cells exposed to LPS was determined by qPCR and Western blot analysis.
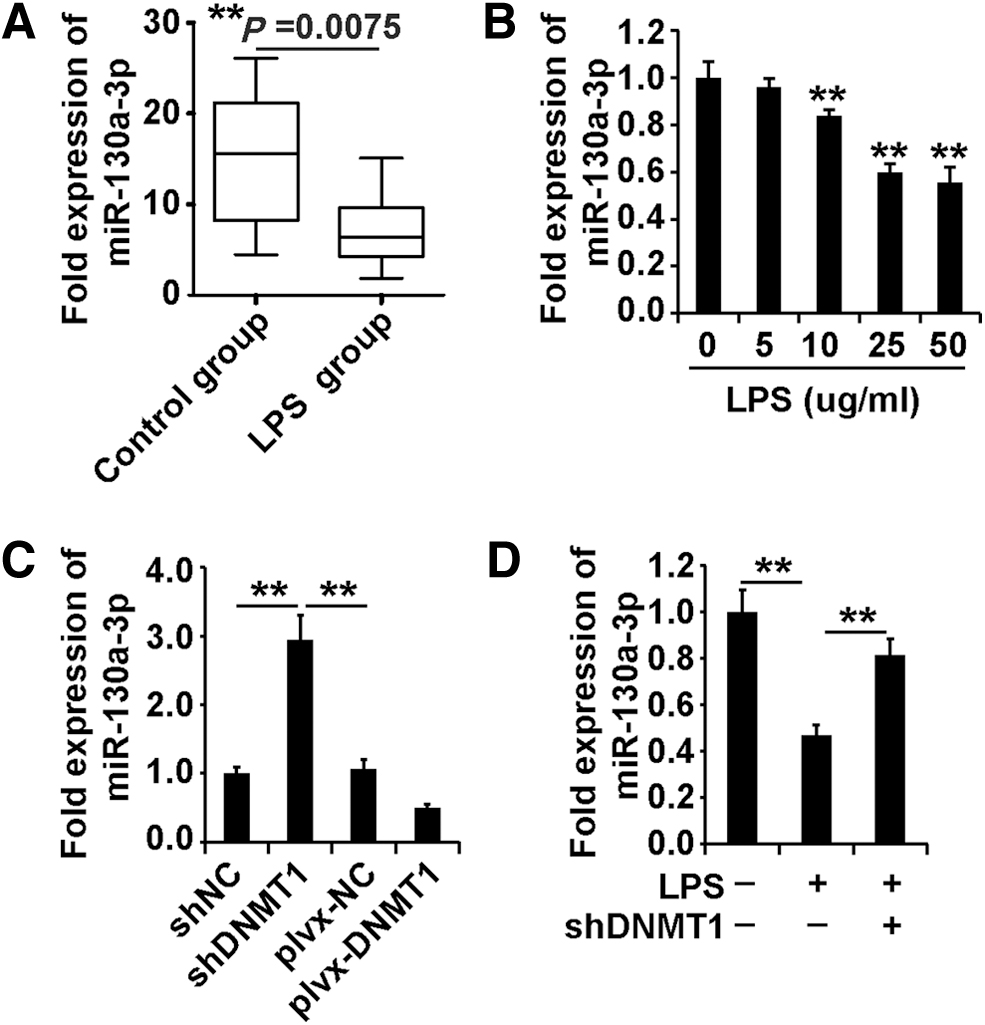
Increased expression of miR-130a in LPS-induced sepsis mouse and cell models.
As shown in Figure 3B, miR-130a expression was decreased in AECII cells exposed to different concentrations of LPS. Taken together, these data indicated that miR-130a expression was decreased in LPS-induced animal and cell models.
We wonder whether miR-130a was regulated by DNMT1. According to the results of qPCR, DNMT1 inhibited miR-130a expression in AECII cells (Fig. 3C). After downregulation of DNMT1, AECII cells were treated with LPS (50 μg/mL) for 48 h, and the expression of miR-130a was determined using qPCR analysis.
As shown in Figure 3D, downregulation of DNMT1 promoted miR-130a expression in AECII cells with or without LPS exposure. In summary, these observations indicated that DNMT1 inhibited miR-130a expression in AECII cells with or without LPS exposure.
miR-130a inhibited ZEB1 expression in AECII cells
To test whether miR-130a could regulate ZEB1 expression in AECII cells, the miR-130a mimics, miR-130a inhibitor, and their control small RNAs were transfected into AECII cells for 48 h. According to the qPCR and Western blot analysis data, miR-130a inhibited the mRNA and protein levels of ZEB1 in AECII cells (Fig. 4A, B). Then, we asked whether miR-130a directly inhibited ZEB1 expression in AECII cells.

miR-130a inhibited ZEB1 expression in AECII cells.
According to data from the luciferase reporter system, miR-130a inhibited the luciferase activity of luciferase-ZEB1 wt, which possesses a binding site for miR-130a, and the luciferase activity of luciferase-ZEB1 mut, which possesses a mutant binding site for miR-130a, in AECII cells (Fig. 4C, D). Collectively, these data proved that miR-130a inhibited ZEB1 expression in AECII cells.
The DNMT1/miR-130a regulatory pathway regulated ZEB1 expression in AECII cells with LPS exposure
According to the qPCR analysis data, ZEB1 expression was upregulated in the LPS-induced sepsis mouse model (Fig. 5A). Based on the results of qPCR and Western blot analysis, ZEB1 expression was upregulated in AECII cells with LPS exposure (Fig. 5B).
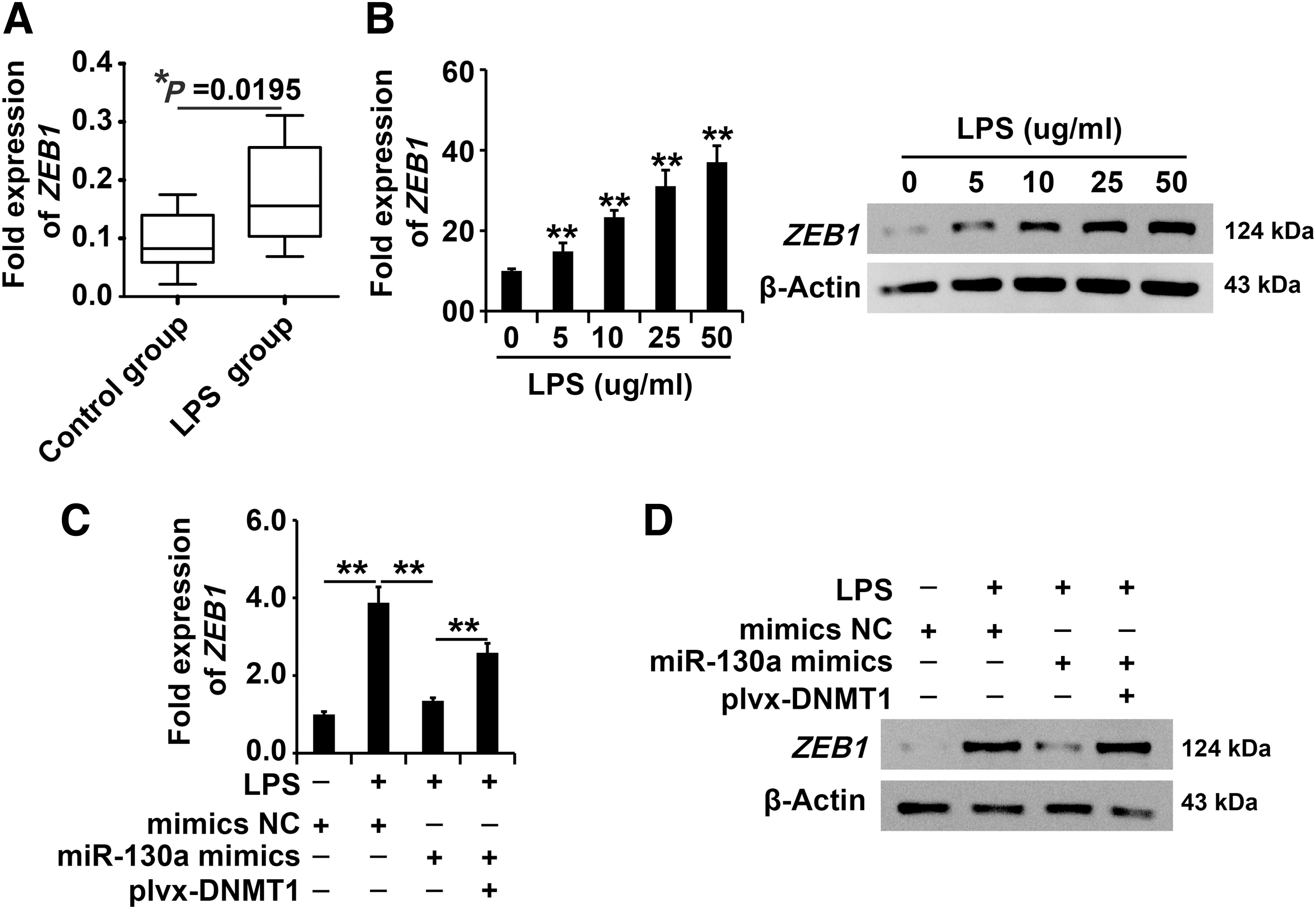
The DNMT1/miR-130a regulatory pathway regulated ZEB1 expression in AECII cells with LPS exposure.
According to the qPCR and Western blot analysis data, upregulation of DNMT1 attenuated miR-130a-induced inhibition of ZEB1 expression in AECII cells with LPS exposure (Fig. 5C, D), indicating that ZEB1 expression was regulated by the DNMT1/miR-130a regulatory pathway in AECII cells with LPS exposure.
The DNMT1/miR-130a/ZEB1 regulatory pathway affected the progress of AECII cells exposed to LPS
Downregulation of the ZEB1 system was successfully constructed (Fig. 6A, B). To systematically study the effect of the DNMT1/miR-130a/ZEB1 regulatory pathway on cellular function in LPS-exposed AECII cells, the cells were treated under the following conditions: control, LPS, LPS plus miR-130a mimics, LPS plus miR-130a mimics plus overexpression of DNMT1, and LPS plus miR-130a mimics plus overexpression of DNMT1 and silencing of ZEB1.
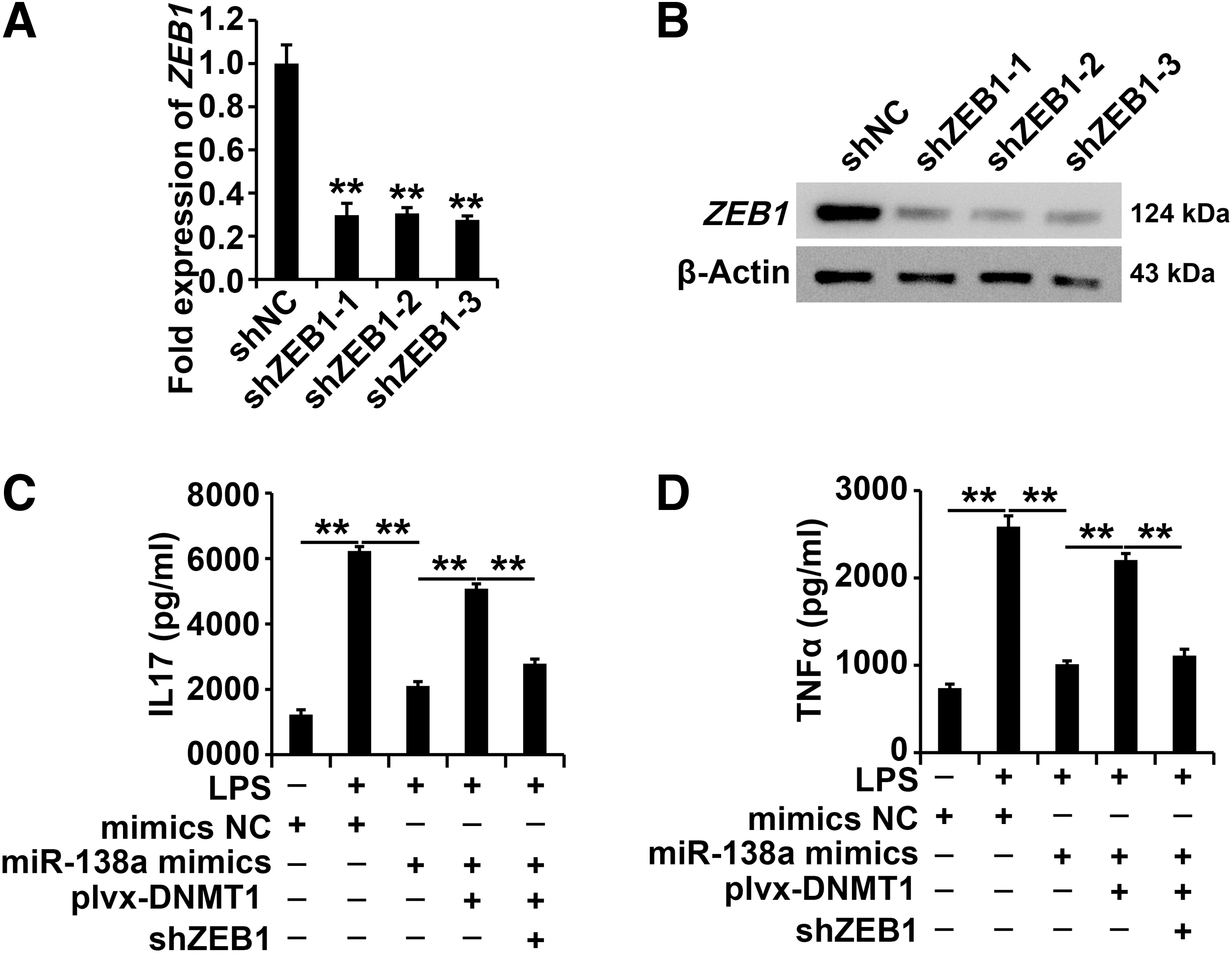
The DNMT1/miR-130a/ZEB1 regulatory pathway affected the progress of AECII cells exposed to LPS.
According to the ELISA data, overexpression of DNMT1 attenuated the role of miR-130a mimics; furthermore, silencing of ZEB1 restored the role of overexpressed DNMT1 in AECII cells under LPS treatment (Fig. 6C, D), indicating the huge role of the DNMT1/miR-130a regulatory pathway in AECII cells exposed to LPS.
Discussion
Sepsis is a global health care issue that affects millions of people (Verdonk et al., 2017). As an autoimmune disease, sepsis is characterized by excessive pro- and anti-inflammatory cytokines in the blood (Hawiger et al., 2015). Therefore, significant reduction of inflammatory factors in the blood of patients with sepsis is an important indicator for the effective remission and treatment of sepsis (van der Poll et al., 2017). This study demonstrated that inhibition of DNMT1 decreased the production of IL-7 and TNF-α in LPS-induced sepsis mice and a cell model.
As an important method of gene expression regulation, DNMT1 is involved in many types of human diseases by affecting a variety of cellular processes (Hamidi et al., 2015). DNMT1-mediated DNA methylation modification exerts a large influence on inflammation-related disease. Wang et al. (2018) demonstrated that prenatal exposure to LPS altered DNMTs by modulating the TNF-α and IL-6/Fli-1 pathways.
Cheng et al. (2014) investigated whether DNMT1-mediated hypermethylation of SOCS1 affected inflammatory cytokine secretion in macrophages exposed to LPS. Shih et al. (2016) revealed that procainamide exerted a therapeutic function by inhibiting DNMT1, thus leading to proinflammatory cytokines in rhabdomyolysis.
Fubing Ma et al. (2019) reported that increased DNMT1-mediated DNA methylation played a crucial role in LPS-induced sepsis by modulating the SMAD2/DNMT1/miR-145 negative regulatory loop. As noted above, silenced DNMT1 inhibited production of IL-7 and TNF-α in LPS-induced sepsis mouse and cell models, resulting in the anti-inflammatory function of DNMT1 in sepsis.
miRNAs exert important roles in various human diseases as well as sepsis (Essandoh et al., 2016; Zhong et al., 2018; Mirna et al., 2019; Szilagyi et al., 2019). Ge et al. (2017) systematically screened the miRNA expression profiles in blood samples from patients undergoing sepsis through microarray analysis. Dagmar Pfeiffer et al. (2017) showed that increased miR-146a, miR-146b, and miR-155 elevated IL-6 and IL-8 expression levels in a sepsis cell model.
Previous studies have shown increased miR-130a expression in sepsis (Cui et al., 2016). However, the regulatory and functional mechanisms of miR-130a in development of sepsis are still largely uncharacterized. In this study, we showed the increased expression of miR-130a in LPS-induced sepsis mice and a cell model. miR-130a was inhibited by DNMT1-mediated DNA methylation, and miR-130a inhibited ZEB1 expression in AECII cells exposed to LPS, indicating that miR-130a plays a crucial role in sepsis.
Conclusions
This study aimed to dissect the role of DNA methylation in the LPS-induced inflammatory response in a sepsis mouse model and AECII cells. The elevated DNMT1 inhibited miR-130a expression and further restored ZEB1 expression in LPS-treated cell models.
Thus, this study is the first to propose a novel functional signaling pathway, DNMT1/miR-130a/ZEB1, in LPS-induced mouse and cell models.
Data Availability
Data are available on request.
Ethics
For this study, no experiments on human subjects were conducted.
Footnotes
Acknowledgment
The authors are thankful for the technical support from the Central Laboratory of Shanghai Pulmonary Hospital.
Disclosure Statement
No competing financial interests exist.
Funding Information
This research did not receive any specific grant from funding agencies in the public, commercial, or not-for-profit sector. Permission for this study was granted by the Department of Emergency, Shanghai Pulmonary Hospital.