
Abstract
Lung cancer is a common life-threatening tumor with high malignancy and high invasiveness. Long non-coding RNAs (lncRNAs) are involved in almost every stage of tumor initiation and progression. Here, we identified an antisense lncRNA, MetaLnc9 antisense (Metalnc9-AS), which arises from the antisense strand of Metalnc9, located on chr9q34.11, while its biological function and mechanism are not clear in lung cancer. In this study, we demonstrated that the expression of Metalnc9-AS was upregulated in non-small cell lung cancer (NSCLC) tissues compared with corresponding non-tumorous tissues. The gain of MetaLnc9-AS was highly associated with the malignant features of NSCLC. Overexpression of MetaLnc9-AS enhanced tumor metastasis in vitro and in vivo. Mechanically, MetaLnc9-AS could form an RNA–RNA hybrid with its cognate sense counterpart, MetaLnc9, to regulate its expression in NSCLC cells, and that such complexes were protected from ribonuclease degradation. Thus, Metalnc9-AS might be a potential and effective treatment for NSCLC.
Introduction
Lung cancer continues to be the leading cause of cancer-related deaths for many years in the world (Barta et al., 2019). It is broadly divided into small cell lung cancer (SCLC, ∼15% cases) and non-small cell lung cancer (NSCLC, ∼85% cases) (Blandin Knight et al., 2017). Despite the emergence of new adjuvant chemotherapy regimens and targeted biologic agents, and significant progress in understanding the pathophysiological mechanisms in NSCLC, the 5-year survival rates and recurrence rate remain poor (Bade and Dela Cruz, 2020). Thus, there is a compelling need to reveal the underlying pathological mechanisms contributing to NSCLC for identifying novel diagnostic and therapeutic targets to improve the efficacy of anti-NSCLC management (Siegel et al., 2014).
Recently, much research has been focused on the function of long non-coding RNAs (lncRNAs) and their mechanisms of action are even more diverse for lung cancer (Nie et al., 2016; Seiler et al., 2017; Park et al., 2018; Tan et al., 2019). Furthermore, evidence has emerged that lncRNAs are known to regulate diverse tumor hallmarks (Goodall and Wickramasinghe, 2021), such as proliferation, viability, immortality, genome instability, motility, and angiogenesis (Gupta et al., 2010; Tsai et al., 2010; Quinodoz and Guttman, 2014; Engreitz et al., 2016; Ge et al., 2017). Thus, studying lncRNAs is a promising way to identify novel molecular markers of lung cancer prognosis and novel therapeutic targets.
Antisense transcripts, defined as a class of lncRNAs that are reverse complements of their endogenous sense counterparts, frequently do not encode proteins (Li et al., 2021). In recent years. Antisense transcript through genic regions, which was initially considered as transcriptional noise, is increasingly being recognized as a key regulator of gene expression (Pelechano and Steinmetz, 2013). Antisense transcript can regulate the expression of their target genes at one or multiple steps of the gene expression process, from transcriptional regulation by gene promoter activation to post-transcriptional regulation by controlling messenger RNA stability and translatability (Villegas and Zaphiropoulos, 2015; Okuyama et al., 2020; Ren et al., 2020).
Here, we describe an antisense RNA, MetaLnc9 antisense (MetaLnc9-AS), located on chromosome 9q34.11, which arises from the antisense strand of the MetaLnc9. There is considerable interest in understanding the mechanism through which antisense transcript function in gene regulation. First, we determined whether MetaLnc9-AS was concordantly or discordantly upregulated with its cognate coding RNA, MetaLnc9, in NSCLC cell lines and tissues. The results revealed that MetaLnc9-AS expression was correlated with the expression of MetaLnc9 both in NSCLC cells and tissues. MetaLnc9-AS exerted its biological functions through the oncogenic targeted gene in NSCLC cells.
Further analyses showed that MetaLnc9-AS could dramatically increase the expression levels of MetaLnc9 by forming protective RNA duplex via their overlapping (OL) region. The results provide a deeper molecular understanding of NSCLC and implicate MetaLnc9-AS as a potential novel therapeutic target in NSCLC.
Materials and Methods
Cell lines and cell culture
The human NSCLC cell lines A549, H1299, H292, H838, PC-9, and H358 were obtained from the American Type Culture Collection (ATCC) in 2010. Cells were maintained at 37°C in a humidified air atmosphere containing 5% carbon dioxide in Dulbecco's modified Eagle's media or RPMI1640. Theses cell lines were Mycoplasma-free and authenticated by quality examinations of morphology and growth profile.
Human NSCLC tissues
The 73 paired samples of human non-small lung cancer and their matched adjacent noncancerous tissues were collected at the time of surgery between 2011 and 2012 at the Department of Lung Cancer, Shanghai Chest Hospital. The use of human clinical specimens in the present study was approved by the Ethics Committee of Fudan University (approval no.050432-4-1911D). The detailed clinical data for each patient is in the Supplementary Table S1. Patients were informed, and they signed consent forms acknowledging the use of their resected tissues for research purposes.
Reverse transcription PCR and quantitative real-time PCR
RNA from the NSCLC tissue specimens and cell lines in this study was extracted using TRIzol reagent (Invitrogen, CA) according to the manufacturer's protocol and quantified with Nanodrop 2000 (Thermo, Japan). First-strand cDNA was synthesized by PrimeScript RT Reagent Kit (TaKaRa). Real-time polymerase chain reaction was performed with SYBR Green Premix (TaKaRa, Tokyo, Japan). Quantitative primers used in this study are shown in Supplementary Table S2.
Vector construction
Full-length MetaLnc9-AS cDNA was amplified using primers, and genomic DNA was used as plate. The primers used for vector are shown in Supplementary Tables S3–S4, 3. The cDNA was cloned into the expression vector pcDNA 3.0 and the lentivirus expression vector pWPXL. The lentiviral vector, pWPXL (which used as lentiviral transfer plasmid was a gift from Didier Trono), and envelope plasmids and the packaging, psPAX2 and pMD2.G, were used to produce lentiviruses in HEK293T.
Oligonucleotide transfection
The sequences of small interfering RNA (siRNA) oligonucleotide and the negative control siRNA are provided in Supplementary Table S5, and they were purchased form RiboBio (RiboBio Biotechnology, Guangzhou, China). Cell were transfected with siRNAs using Lipofectamine 2000 Reagents (Invitrogen).
Cell migration and invasion assays
Cell migration and invasion assays were performed in a 24-well plate with 8-μm pore size chamber inserts (Corning). For migration assays, 5 × 104 cells were placed into the upper chamber per well with the non-coated membrane. For invasion assays, 1 × 105 cells were placed into the upper chamber per well with the Matrigel-coated membrane that was diluted with serum-free culture medium. Cells that moved to the bottom surface of the chamber were fixed with 100% methanol, stained with 0.1% crystal violet for 30 min. Then, the cells were imaged and counted under a CKX41 inverted microscope (Olympus, Japan).
Animal model
Twenty-four BALB/c nude mice about 5 weeks of age were purchased from Shanghai Cancer Institution (Shanghai, China) and were randomly divided into two groups: Control group and MetaLnc9-AS group (12 mice/group). All mice were maintained in the specific pathogen-free conditions, and animal experiments were only performed once. For tail vein injection mouse model, 2 × 106 A549 cells, of the stably overexpressing MetaLnc9-AS or control group, were injected intravenously through tail vein into the nude mice (12 mice/group).
All mice were euthanized after 6 weeks. The lungs were fixed in 4% paraformaldehyde and embedded in paraffin. The number of metastatic foci in the lung were determined using the hematoxylin eosin (H&E) staining in tissue sections under a binocular microscope. All animal experiments were conducted in accordance with relevant guidelines and regulations and were approved by the Institutional Animal Care and Use Committee of Fudan University (approval. No. FUSCC-IACUC-S20210310).
Immunoblotting analysis
The cell lysates were extracted from cultured cells with a mixture of Protein Extraction Reagent (Thermo), PhosSTOP (Roche), and Complete Mini (Roche). Protein samples were separated by 6% and 8% sodium dodecyl sulfate-polyacrylamide gel electrophoresis and transferred to nitrocellulose filter membranes (Millipore). After blocking in phosphate buffered saline/Tween-20 containing 5% nonfat milk, the membranes were incubated with the primary antibodies. Information on the antibodies are provided in Supplementary Table S6. Unprocessed original scans of blots are shown in Supplementary Figure S4.
Statistical analysis
The data were presented as the mean ± standard error of the mean from one representative experiment out of three independent experiments. General paired or unpaired t-tests and Chi-square tests were conducted using SPSS (IBM, NY) or R (
Results
Identification of an OL antisense transcript at the MetaLnc9 locus
In our previous study, we demonstrated MetaLnc9 was a novel oncogenic lncRNA in NSCLC and facilitated lung cancer metastasis through PGK1-activated AKT/mTOR pathway (Yu et al., 2017). From the University of California Santa Cruz (UCSC) genome browser, we found a single antisense RNA (Tcons_0016491) that was transcribed from the negative strand of the MetaLnc9 locus, located at the chromosomal band 9q34.11, and consist of two exons (Fig. 1A). We therefore refer to Tcons_0016491 as MetaLnc9-AS transcript. MetaLnc9-AS shares a 1279 nucleotide-long region with the third and fourth exons of MetaLnc9 (from RefSeq NR_038955.1). We refer to this region as the OL region (Fig. 1A).
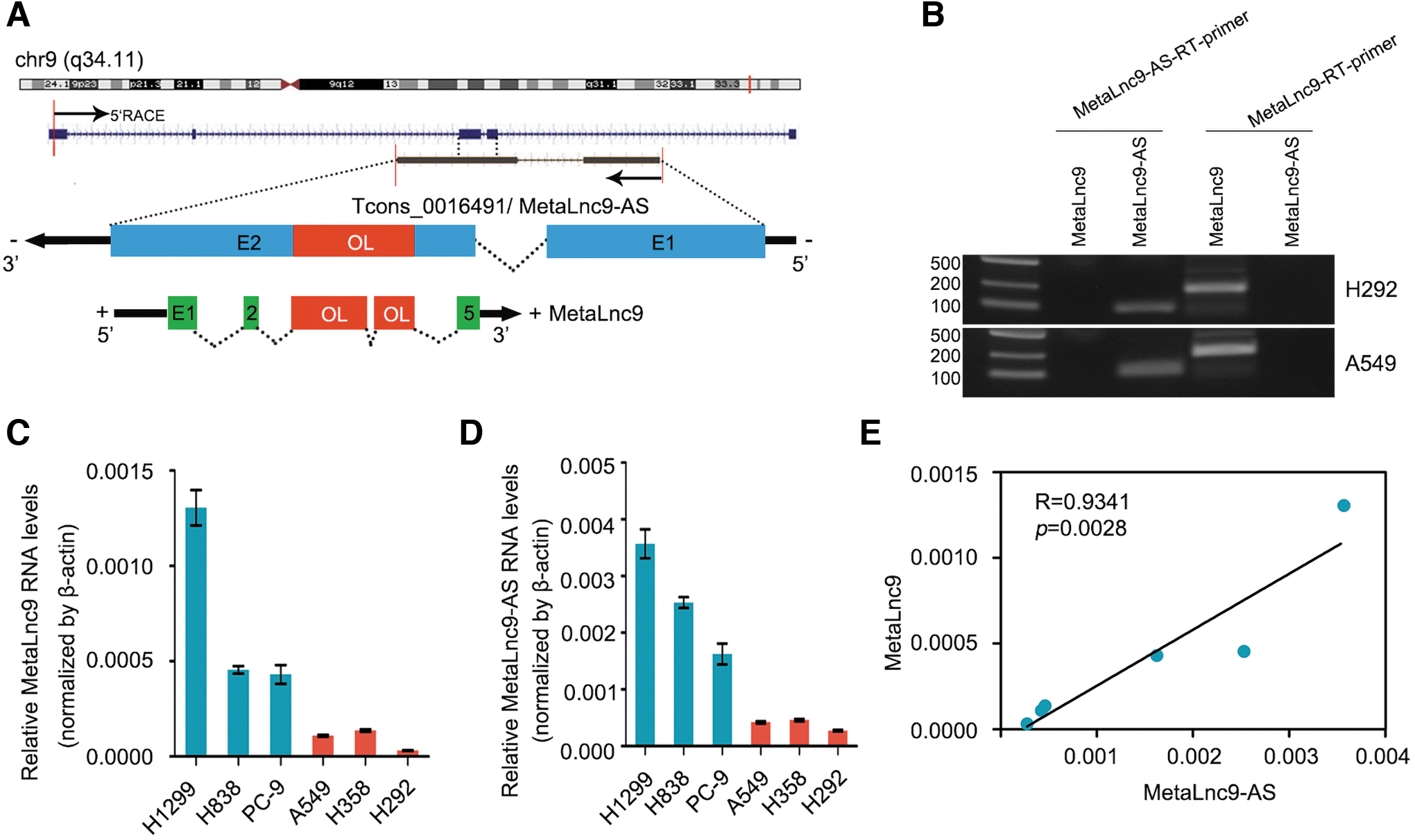
Identification of an OL antisense transcript at the MetaLnc9 locus.
We next analyzed the coding potential of MetaLnc9-AS transcript by using three independent mathematical methods, including PhyloCSF codon substitution frequency analysis (Lin et al., 2011), Coding Potential Calculator (Kong et al., 2007), and Coding Potential Assessment Tool (Wang et al., 2013), which showed that MetaLnc9-AS did not have protein-coding potential (Supplementary Figs. S1A and S2A, B). In addition, we performed single strand-specific RT-PCR to identify MetaLnc9-AS as originating from the reverse strand of the MetaLnc9 locus in H292 and A549 cells. Antisense strand-specific complementary DNA was acquired via first-strand RT using an antisense-specific RT primer. As shown in Figure 1B, MetaLnc9-AS was successfully amplified from antisense-specific RT complementary DNA.
According to recent research, antisense LncRNA appear to exert trans or cis effects on other genes that include suppression and activation, and their mechanism include transcriptional regulation by gene promoter activation and post-transcriptional regulation by affecting mRNA stability (Jadaliha et al., 2018). First, we investigated the endogenous levels of MetaLnc9 and MetaLnc9-AS in six human NSCLC cell lines (Fig. 1C, D). Identification of the MetaLnc9-AS and MetaLnc9 expression levels in these cell lines showed that MetaLnc9-AS and MetaLnc9 were significantly positively correlated in NSCLC cells (Fig. 1E).
MetaLnc9-AS is overexpressed in NSCLC and associated with advanced clinical stage
To explore the clinical significance of MetaLnc9-AS, we detected the expression of MetaLnc9-AS in 73 pairs of human NSCLC and their corresponding noncancerous lung tissues using qRT-PCR. The results showed that MetaLnc9-AS expression was significantly upregulated in NSCLC tissues compared with corresponding non-tumorous tissues (Fig. 2A, B).

MetaLnc9-AS is overexpressed in NSCLC and associated with advanced clinical stage.
In addition, we also observed a statistically significant increase in the expression of MetaLnc9-AS in lung cancer with advanced (stage III and IV) to early stages (stage I and II) (Fig. 2C), Interestingly, when 73 NSCLC samples were stratified on the basis of the status of lymph node and distal metastasis, we observed that MetaLnc9-AS expression was further significantly increased in NSCLC tissues that had lymph node or distal metastasis compared with those that had not spread (Fig. 2D). In addition, there was a positive relationship between MetaLnc9-AS and MetaLnc9 expression in NSCLC tissues (Fig. 2E). Collectively, these data strongly show that we identify an OL antisense transcript at the MetaLnc9 locus, and it potentially has a critical role in the aggressiveness of NSCLC.
MetaLnc9-AS increases MetaLnc9 expression at RNA levels
To identify the direct link between the RNA expression of MetaLnc9-AS and MetaLnc9, we found that knockdown or overexpression of Metalnc9 did not affect the expression levels of MetaLnc9-AS (Fig. 3A, B). However, we found that overexpression of MetaLnc9-AS in A549 and H292 cells could dramatically increase the expression levels of MetaLnc9, and the upregulation of MetaLnc9 induced by MetaLnc9-AS could inhibit by MetaLnc9-siRNA (Fig. 3C–F).
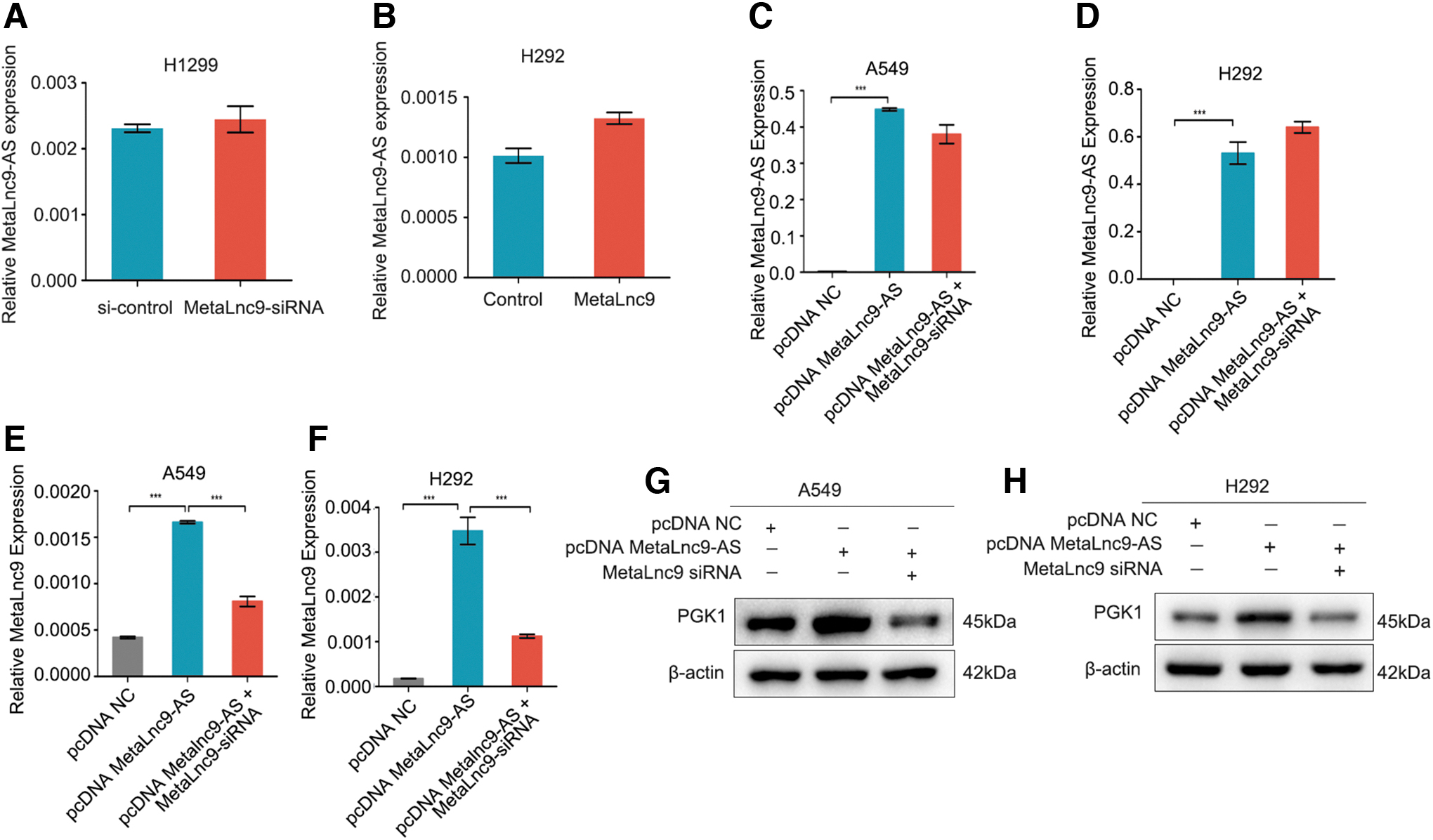
MetaLnc9-AS increases MetaLnc9 RNA expression in NSCLC cells.
As MetaLnc9 played its role in NSCLC cells via interacting with PGK1 (Yu et al., 2017), we next investigated whether MetaLnc9-AS also had effect on PGK1 protein expression. Interestingly, overexpression of MetaLnc9-AS significantly increased the expression levels of PGK1 protein expression, and the promotion could be suppressed by MetaLnc9-siRNA in A549 and H292 cells (Fig. 3G, H). Collectively, these finding suggest that regulation of MetaLnc9 expression by MetaLnc9-AS occurs at the RNA levels.
MetaLnc9-AS acts as an oncogenic antisense transcript in vitro and in vivo in NSCLC
To investigate the biological function of MetaLnc9-AS in NSCLC, A549 and H292 cells were infected with pcDNA vector harboring MetaLnc9-AS sequence, to generate overexpressing cell lines. Migration and invasion assays showed that MetaLnc9-AS overexpression significantly promoted the migration and invasion abilities in A549 and H292 cells, and the promotion abilities could be suppressed by MetaLnc9-siRNA (Fig. 4A, B).
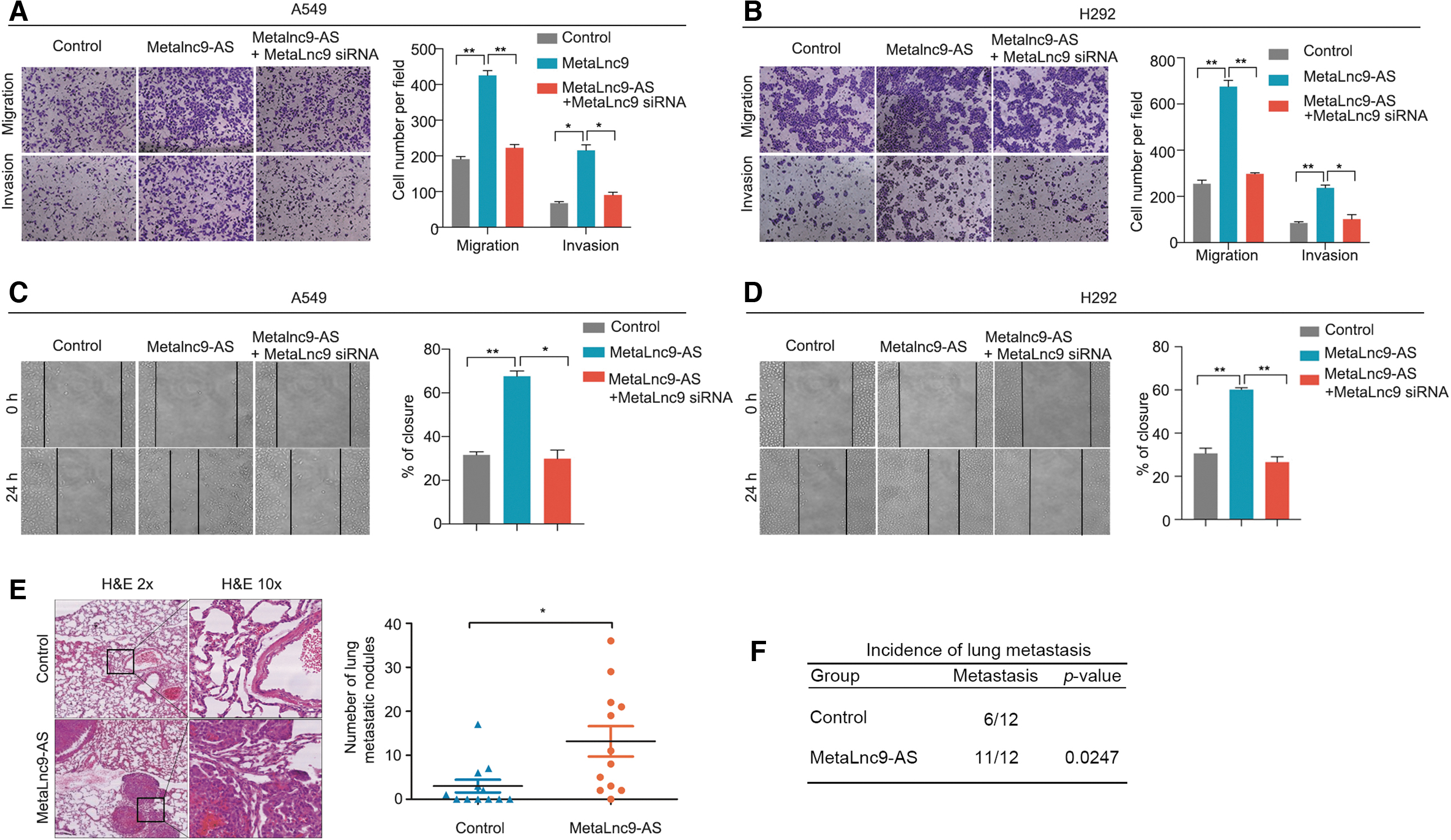
Metalnc9-AS as the oncogenic antisense transcript in vitro and in vivo.
Subsequently, we also found that MetaLnc9-AS overexpression could enhance the migration rates compared with the control group, and the promotion abilities also could decrease by MetaLnc9-siRNA (Fig. 4C, D). Additionally, overexpressed MetaLnc9-AS accelerated cell growth in H292 cells (Supplementary Fig. S5A), and knockdown of MetaLnc9-AS led to a significant decrease in H1299 cells (Supplementary Fig. S5B).
Next, to validate the oncogenicity of MetaLnc9-AS in vivo, we used tail vein injection mouse model by injecting the pWPXL-MetaLnc9-AS A549 cells and the control group into the tail vein of nude mice, and we compared the rates of lung colonization as measured by H&E staining. Six weeks later, the mice were sacrificed and the number of metastatic lesions of each mouse was detected and counted under microscope. As expected, MetaLn9-AS promoted lung metastasis formation. The number of lung metastatic nodules were significantly increased in the MetaLnc9-AS group (Fig. 4E). Conversely, the metastasis rates in the MetaLnc9-AS overexpression group were higher than the control group (Fig. 4F). Taken together, these results suggest that MetaLnc9-AS acts as an oncogenic antisense transcript in NSCLC.
The overlap region of MetaLnc9-AS has a critical role in NSCLC
After verifying the regulation by antisense RNA, we constructed the MetaLnc9-AS- OL (overlap) and –NOL (non-OL) region vector for transfection of A549 and H292 cells (F ig. 5A). We found that the OL region of MetaLnc9-AS vector promoted the MetaLnc9 expression levels, whereas the NOL vector failed to increase MetaLnc9 expression levels (Fig. 5B, C). Consistently, it was the OL region of the MetaLnc9-AS that promoted the expression levels of PGK1 protein expression, and the NOL vector had no effect on PGK1 protein expression in A549 and H292 cells (Fig. 5D, E).
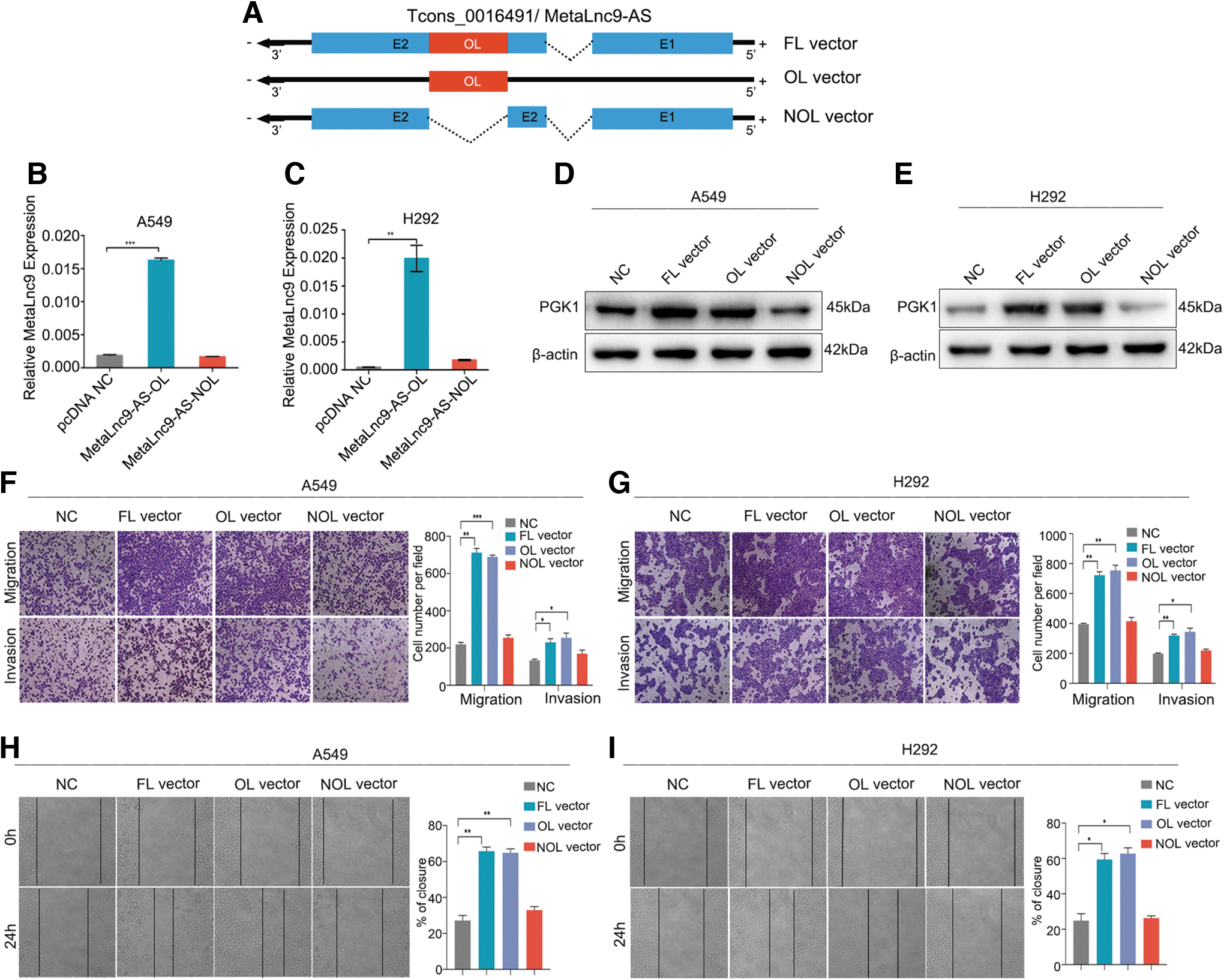
The OL region of MetaLnc9-AS has a critical role.
Moreover, both OL and full length (FL) of MetaLnc9-AS vector promoted the migration and invasion abilities of A549 and H292 cells, whereas the NOL vector did not affect the migration and invasion abilities of A549 and H292 cells (Fig. 5F, G). The OL region of MetaLnc9-AS could enhance the migration rates of A549 and H292 cells and the NOL vector had no effect on NSCLCL cells (Fig. 5H, I). These experiments established that the OL portion of MetaLnc9-AS alone exerted the same regulatory effect and function as FL-MetaLnc9-AS did.
MetaLnc9-AS increases MetaLnc9 RNA stability by forming RNA duplex
Knowledge of subcellular localization of lncRNA can provide a clue about the regulation mechanism of MetaLnc9-AS. For the fractionation assay, we extracted the cytoplasmic and nuclear fractions from A549 cells to detect the distribution of MetaLnc9 and MetaLnc9-AS. As showed in Supplementary Figure S3A, MetaLnc9 was almost equally distributed between cytoplasmic and nuclear fractions, but MetaLnc9-AS was markedly enriched in cytoplasmic fractions. It has been studied that antisense RNA can form duplexes with their cognate sense mRNAs, and that such complexed are protected from ribonuclease degradation. LncRNA KRT7-AS increases its cognate sense counterpart KRT7 by forming an RNA duplex (Huang et al., 2016).
We therefore performed RNase protection assay to verify the hypothesis that MetaLnc9-AS and MetaLnc9 form a protective duplex, specifically at their OL region. Total RNA from A549 cells were used to test this hypothesis. We found that the OL region of MetaLnc9-AS was at least partially protected from RNase degradation (Fig. 6A, B).
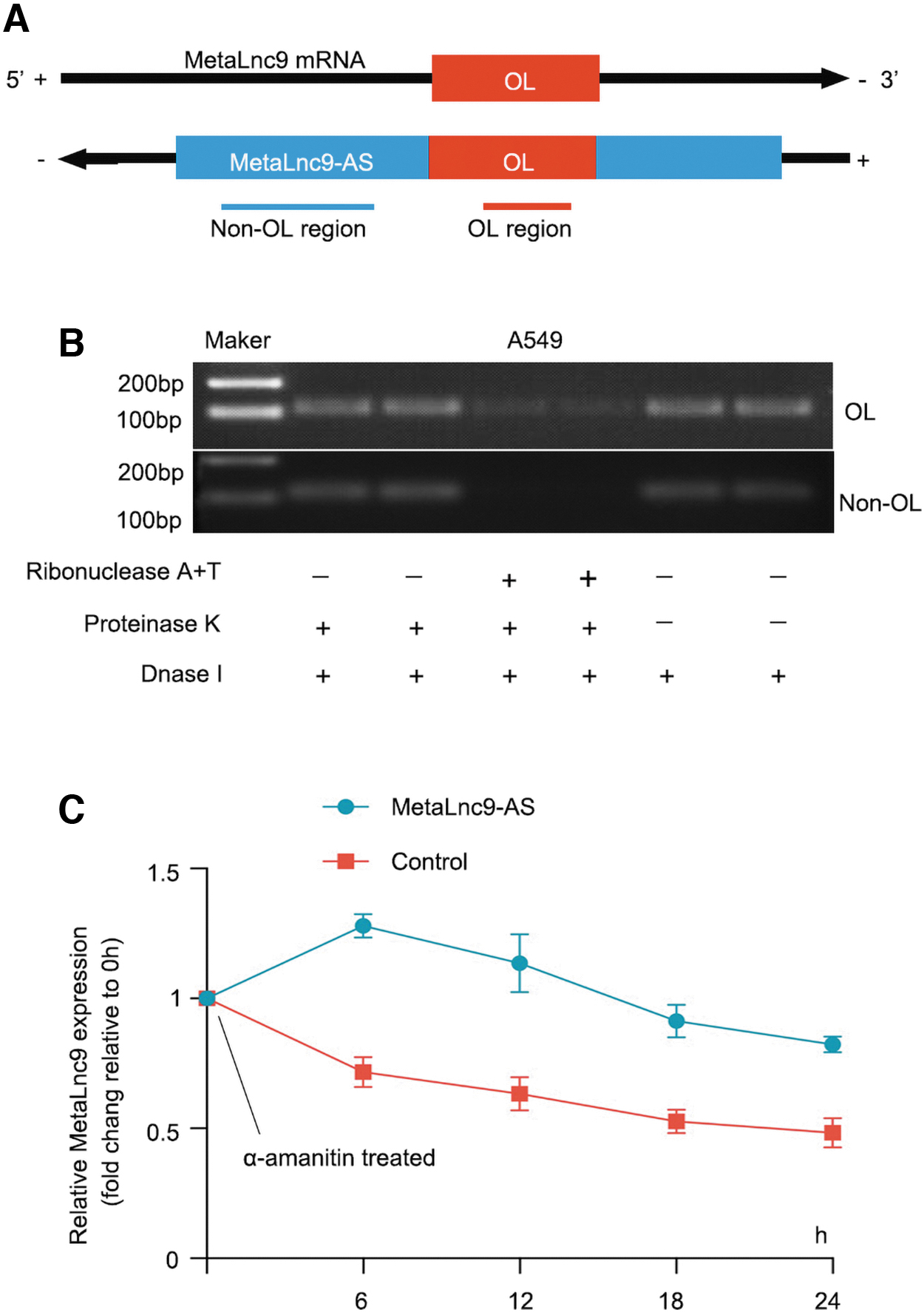
MetaLnc9-AS increases MetaLnc9 RNA stability by forming RNA duplex.
To explore whether MetaLnc9 RNA stability was influenced by MetaLnc9-AS, we used α-amanitin (an inhibitor of RNA synthesis) to block new RNA synthesis in A549 cells with overexpression of MetaLnc9-AS over a 24 h period, then detected the subsequent expression levels of MetaLnc9. We found MetaLnc9-AS promoted the stability of MetaLnc9 compared with the control (Fig. 6C). Collectively, these results demonstrated that MetaLnc9-AS activates its sense partner by forming RNA–RNA duplex at their complementary OL region and this duplex increases its stability.
Discussion
Recent advances in high-throughput sequencing, including microarray and RNA-seq, have allowed researchers to discover a number of novel disease-associated lncRNAs (Wilusz et al., 2009). While thousands of lncRNAs have been detected in many cancers, detailed functional characterization has been limited to only a small portion of those identified (Hu et al., 2018). LncRNAs can have a variety of roles within the cell, including nuclear regulation of transcription and mRNA stability and cytoplasmic regulation (Zhang et al., 2019). Here, we identified a new antisense RNA, MetaLnc9-AS, that is transcribed from the negative strand of the MetaLnc9 locus.
Antisense transcripts, which is a class of lncRNAs, are transcribed from the strand opposite to that of the sense transcript of either non-protein-coding or protein-coding genes (Katayama et al., 2005). Antisense transcripts appear to function in cis (i.e., those that affect alleles on the DNA strand from which they are produced, usually locally) and in trans (i.e., those that affect alleles on different DNA strands) (Pan and Zhao, 2021). For example, the stabilization of PHO84 antisense transcripts correlates with a decrease in the association of Rrp6 with the PHO84 gene (Camblong et al., 2007). Long non-coding antisense RNA KRT7-AS is activated in gastric cancers and supports cancer cell progression by increasing KRT7 expression (Huang et al., 2016).
Acetylcholinesterase-AS, another AS RNA, directly represses acetylcholinesterase expression via epigenetic modification of the acetylcholinesterase promoter region (Xi et al., 2014). BDNF-AS tonically represses BDNF sense RNA transcription by altering chromatin structure at the gene locus (Modarresi et al., 2012). WDR83 and DHPS were capable of forming an RNA duplex at OL 3′ untranslated regions and this duplex increased their mutual stability (Su et al., 2012). The above data revealed that such antisense RNA–sense RNA interactions may have critical roles in regulating gene expression at the transcriptional or post-transcriptional.
In this study, we found MetaLnc9-AS and MetaLnc9 was concordantly upregulated in NSCLC cell lines and tissues. Importantly, we found that MetaLnc-9-AS acted as an oncogene and significantly enhanced the migration and metastasis of lung cancer cells in vitro and in vivo. Mechanically, MetaLnc9-AS stabilized MetaLnc9 RNA by forming protective RNR–RNA duplex and promoted MetaLnc9 RNA expression. Thus, MetaLnc9-AS illustrates a novel type of long non-coding antisense RNA function and mechanism in NSCLC. Further studies of this may provide a complete understanding of their mechanism in tumor progression and eventually open avenues for novel therapeutic interventions in NSCLC.
Footnotes
Authors' Contributions
Conception and design: Z.Y. and T.Y. Development of methodology: K.L. and H.P. Analysis and interpretation of data: Z.Y. and L.S. Writing and revision of the article: Z.Y., K.L., and T.Y. All the authors reviewed the article before submission and approved the final article.
Disclosure Statement
No competing financial interests exist.
Funding Information
This work was supported by the National Natural Science Foundation of China (grant no.: 81802270).
Supplementary Material
Supplementary Figure S1
Supplementary Figure S2
Supplementary Figure S3
Supplementary Figure S4
Supplementary Figure S5
Supplementary Table S1
Supplementary Table S2
Supplementary Table S3
Supplementary Table S4
Supplementary Table S5
Supplementary Table S6
References
Supplementary Material
Please find the following supplemental material available below.
For Open Access articles published under a Creative Commons License, all supplemental material carries the same license as the article it is associated with.
For non-Open Access articles published, all supplemental material carries a non-exclusive license, and permission requests for re-use of supplemental material or any part of supplemental material shall be sent directly to the copyright owner as specified in the copyright notice associated with the article.