
Abstract
Dihydroartemisinin (DHA) is a traditional antimalarial drug. DHA plays a crucial role in preventing pulmonary hypertension (PH); however, its regulatory function on microRNAs (miRNAs) in PH remains unclear. This study aimed to investigate whether DHA exerts its protective functions by regulating miR-335 in PH. Hypoxia-induced PH models were induced both in vitro and in vivo. Mice were treated with various concentrations of DHA, and pulmonary arterial smooth muscle cells (PASMCs) were treated with DHA, miR-335 inhibitor, miR-335 mimic, or Van Gogh-like 2 (Vangl2) plasmid. The expression of miR-335 and Vangl2, pulmonary arterial remodeling index; right ventricular hypertrophy index; and proliferation and migration indexes were measured. DHA improved pulmonary vascular remodeling and alleviated PH in vivo. miRNA sequencing and real-time PCR results further show that the increase in hypoxia-induced miR-335 was avoided by DHA administration, and miR-335 increased the hypoxia-induced PASMC proliferation and migration. MiRNA databases and dual-luciferase reporter assay show that miR-335 directly targets Vangl2, and Vangl2 decreased the hypoxia-induced PASMC proliferation and migration. The miR-335 inhibitor failed to inhibit hypoxia-induced proliferation and migration upregulation in Vangl2 knockdown PASMCs, and the effect of DHA can be blocked by miR-335 upregulation. In hypoxic PH, MiR-335 is increased, whereas Vangl2 is decreased. MiR-335 can significantly promote the hypoxia-induced proliferation and migration of PASMCs by targeting the Vangl2 gene. DHA effectively reverses the hypoxia-induced upregulation of miR-335 expression, avoiding the miR-335-mediated downregulation of Vangl2 and thereby promoting the expression of Vangl2 to prevent PH.
Introduction
Pulmonary hypertension (PH) is a devastating and chronic disease identified by an abnormal increase in pulmonary circulation pressure, eventually resulting in right ventricular (RV) failure and death. Hypoxia triggers hypoxic pulmonary vasoconstriction and pulmonary vascular remodeling, leading to the formation of hypoxic PH, which is the third category of PH classification (Simonneau et al., 2019). Pulmonary arterial smooth muscle cells (PASMCs) are the major components responsible for pathologic vascular remodeling, and the role of PASMC in pulmonary vascular remodeling includes abnormal proliferation, cell migration, and phenotypic transformation (Wang et al., 2021). Despite much effort made in the treatments that improve the symptoms and outcomes of PH, the management of PH is still poor (Hoeper et al., 2021). Thus, it is essential to search for novel drugs and therapeutic targets for PH.
Dihydroartemisinin (DHA) is a semisynthetic derivative of artemisinin with a longer half-life, higher water solubility, and antiparasitic activity than artemisinin (Tiwari and Chaudhary, 2020). Many studies have reported new activities of DHA in addition to antimalarial activity, such as anti-inflammatory, antitumor, and antibacterial activities (Zhang et al., 2016; Yang et al., 2019; Bai et al., 2021). Studies have shown that DHA has a certain protective effect on the cardiovascular system (Yin et al., 2018). We previously reported that DHA is active in inhibiting the hypoxia-induced increase in human pulmonary artery endothelial cell proliferation, migration, and oxidative stress (Yu et al., 2018a). Nevertheless, the underlying regulatory mechanisms are still unclear.
MicroRNA (miRNA) is a class of noncoding small RNA that negatively regulates target gene expression. MiRNA regulates gene expression at the post-transcriptional level by regulating the degradation of target gene mRNA. MiRNA also modulates the target mRNA translational repression by specifically binding to the 3′-untranslated region of the target gene (Van Meter et al., 2020). MiRNA is a key regulator of a variety of cellular processes, and it plays a key role in cardiovascular diseases including PH. Growing evidence suggests that miRNAs play a significant role in the pathogenesis of PH and in maintaining pulmonary vascular homeostasis (McLendon et al., 2015; Wallace et al., 2015; Yu et al., 2018b). Recent studies have reported that blocking miR-335-3p could attenuate hypoxic PH in mice (Fan et al., 2020). Our early verification through high-throughput sequencing and PCR experiments showed that DHA can reverse the overexpression of miR-335 under hypoxia. Therefore, we speculated that miR-335 might be involved in the attenuation of PH by DHA.
Van Gogh-like 2 (Vangl2) is the core gene of planar cell polarity (PCP) signaling pathway, encoding a four-pass transmembrane protein (Bailly et al., 2018). Aberrant Vangl2 has been shown to be related to the occurrence and development of cancer, lung, and kidney disorders (Piazzi et al., 2013; Papakrivopoulou et al., 2018; Cheong et al., 2020). It has been shown that mutation in the PCP gene Vangl2 leads to disruption of lung development and defects in lung structure (Yates et al., 2010). Our previous experiments showed that the expression of Vangl2 decreased under hypoxia, and Vangl2 was found to be one of the target genes of miR-335 in the miRNA database. Therefore, we assumed that Vangl2 might be a potentially useful target for hypoxia-induced PH, and miR-335 might promote PH by targeting Vangl2.
Therefore, we aimed to better understand the mechanism of DHA in alleviating hypoxic PH and focused on the miRNA level in this study. Presequencing showed that miR-335 might be involved in hypoxic PH, and we predicted that there might be a functionally regulated target gene Vangl2. We further evaluated whether DHA attenuates hypoxia-induced PH by inhibiting the negative regulatory effects of miR-335 on Vangl2. This will provide new results on the regulation of DHA at the miRNA level and experimental evidence for its future applications.
Materials and Methods
Reagents
DHA was purchased from TCI (Tokyo, Japan). Rabbit antibody against Vangl2 (Lot No. 21492-1-AP) was obtained from Proteintech (Chicago, IL). Rabbit antibody against matrix metalloproteinase 2 (MMP2; Lot No. ab92536) was obtained from Abcam (Cambridge, United Kingdom). Rabbit antibodies against β-actin (Lot No. 4970) and proliferating cell nuclear antigen (PCNA; Lot No. 13110) were obtained from Cell Signaling Technology (Beverly, MA). Horseradish peroxidase (HRP)-conjugated goat anti-rabbit IgG antibody (Lot No. BL003A) was obtained from Biosharp (Hefei, China).
Animal models
Male C57BL/6 mice (2–3 months old, 20–30 g) and Sprague-Dawley rats (180–220 g) were purchased from Vital River Laboratory Animal Technology (Beijing, China). The experimental scheme and housing related to the animals were approved by the Animal Ethics Committee of Wenzhou Medical University (institutional approval No.: SYXK [Zhe] 2020–0014). The laboratory mice and rats were kept in a humidity of 55–65% and 20–24°C temperature. The laboratory mice were randomized into seven groups: normoxia (N, saline-treated) group, hypoxia (H, saline-treated) group, H + DHA (30 mg/kg i.g.) group, H + DHA (60 mg/kg i.g.), H + DHA (90 mg/kg i.g.), H + DHA (120 mg/kg i.g.), and H + DHA (150 mg/kg i.g.). The laboratory rats were randomized into three groups: N group, H group, H + DHA (45 mg/kg i.g.) group. There were 5 mice and 5 rats in each group, a total of 35 mice and 15 rats. All groups except the N group were housed in a normobaric chamber with 10% oxygen (O2) for 21 days, and the N group was kept in ambient air.
Measurement of hemodynamic and RV hypertrophy
The mice and rats were anesthetized with 20% urethane (1 mL/100 g, i.p.), and catheters were then inserted into the RV, pulmonary artery, and left carotid arteries (Cai et al., 2019). The right ventricular systolic pressure (RVSP), a surrogate for the pulmonary artery pressure, was measured by catheterization via jugular vein to right ventricle. Pressure transducers (PowerLab 8/35 Multichannel Biological Signal Recording System, AD Instruments, Australia) were used to record the RVSP, mean pulmonary artery pressure (mPAP), and mean carotid artery pressure (mCAP). Animals were then killed for further examination. The RV was isolated from the left ventricle (LV) and septum (S), and the RV hypertrophy was indicated by the ratio of RV to (LV + S) weight.
Measurement of pulmonary arterial remodeling
Pulmonary arterial remodeling was assessed using the method depicted by Cai et al. (2019). In brief, the connective tissue around the lung was removed, and 4% paraformaldehyde was used to fix the lung tissue for 24 h. Then, the lung tissue was cut into 5-μm-thick slices after being embedded in paraffin. Sections were then stained with hematoxylin–eosin (HE), and an optical microscope was used to evaluate the structure of pulmonary arteries with external diameters of 25–100 μm. Then, Image-Pro Plus 6.0 (Media Cybernetics, Bethesda, MD) was utilized to analyze the ratios of the wall thickness (WT) to the total thickness (TT) and the pulmonary artery wall area (WA) to the total area (TA).
Immunofluorescence detection is also a method to evaluate the WT of small pulmonary arteries in mice. First, the paraffin section of lung tissue is placed in sodium citrate buffer for antigen retrieval in a microwave oven. The slices were sealed with 5% fetal bovine serum (FBS) and incubated with 0.5% α-SMA primary antibody overnight at 4°C. After incubating with secondary antibody (1:200) for 1 h at room temperature the next day and washing with phosphate-buffered saline (PBS) three times, DAPI was added to the slice. Finally, a fluorescence microscope was used to observe the immunolabeled sections.
sRNA sequencing and bioinformatics analysis
The RNA required for sequencing in this study was extracted from mouse lung tissue. Total RNA was isolated using TRIzol (Invitrogen). The integrity and quantity of the products were evaluated using Agilent 2200 TapeStation (Agilent Technologies) and Qubit®2.0 (Life Technologies). For sRNA sequencing, 1 μg of the mixed total RNA was used for sRNA library construction. sRNA libraries were constructed using NEBNext® Multiplex Small RNA Library Prep Set for Illumina (NEB) and sequenced using a HiSeq 2500 (Illumina) platform at RIBOBIO (Guangzhou, China).
In total 1915 miRNAs were identified. A differential expression analysis was performed using “Limma” package with a cutoff p < 0.05 and |fold change (FC)| of >1.5. Heat maps and volcano plots were drawn on the dif-miRNA of N group versus H group and H group versus H + DHA group (H + D) using “pheatmap” (
Validation of Gene Expression Omnibus database analysis
GSE53408 was selected from the Gene Expression Omnibus (GEO;
qRT-PCR
Total RNA was isolated from mouse lung tissues and rat PASMCs using a UNlQ-10 Column Trizol Total RNA Isolation Kit (Sangon Biotech, Shanghai, China). MiRNA and mRNA were quantified using a UV–vis spectrophotometer (NanoDrop, Thermo Fisher Scientific), and only when the OD260/OD280 ratio of miRNA and mRNA were between 1.8 and 2.0, the miRNA and mRNA was used for subsequent reverse transcription using a miRNA First-Strand cDNA Synthesis Kit (stem-loop method) and AMV First-Strand cDNA Synthesis Kit obtained from Sangon Biotech. Finally, quantitative real-time PCR was conducted using a miRNA qPCR Kit and SG Fast qPCR Master Mix (Sangon Biotech) on CFX96 Real-Time PCR Detection System (Bio-Rad). U6 snRNA was selected as the internal control for miRNA, and the relative miRNA and mRNA expressions were calculated by the 2−ΔΔCt method. The primer sequences are given in Table 1.
The Sequence of Mimic, Inhibitor, siRNA, and Primers
Cell treatment
PASMCs were isolated from male Sprague-Dawley rats (180–220 g) using mature methods as previously mentioned (Huang et al., 2018). For all cell experiments, a total of five rats were used (institutional approval No.: SYXK [Zhe] 2020-0014). PASMCs from passages 4–6 were used for the study. PASMCs were assigned to the following groups: (1). N, H, H + miR-335 inhibitor (100 nM); (2) N, H, H + Vangl2 plasmid; (3) N, H, H + miR-335 inhibitor (100 nM), H + miR-335 inhibitor (100 nM) + Vangl2 siRNA (50 nM); (4) N, H, H + DHA (5 μM), H + DHA (5 μM) + miR-335 mimic (50 nM). The N group was cultivated in a normal incubator with 21% O2, 74% nitrogen (N2), and 5% carbon dioxide (CO2) at 37°C for 48 h. H groups were cultured with 5% O2, 90% N2, and 5% CO2 at 37°C for 48 h. In the rescue experiment, various chemical stimuli were added to PASMCs at the same time before the cells were cultivated in a hypoxic incubator.
Proliferation assays
PASMCs were seeded into 96-well plates at a density of 10,000 cells/well. After overnight attachment, PASMCs were subjected to various treatments for 48 h. Then, cell counting kit-8 (CCK8, Dojindo, Japan) reagent (10 μl) was added into each well. PASMCs were incubated at 37°C for 2 h, and the absorbance at 450 nm was measured. A 5-ethynyl-2′-deoxyuridine (EdU) cell proliferation assay kit (RiboBio, Guangzhou) was also used to assess cell proliferation. EdU is the modified analog of thymidine and incorporated into newly synthesized DNA. The EdU-positive rate was indicated by the ratio of EdU-positive cells (red cells) to the Hoechst 33342-positive cells (blue cells).
Transwell migration chamber assay
For the Transwell assay, a 24-well Transwell system (8 μm; Corning Inc., NY) was used. In the lower chamber of Transwell plates, Dulbecco's modified Eagle medium (DMEM) containing 5% FBS was added with the experimental treatments in each group. PASMCs were suspended in DMEM and seeded into the upper chambers at a density of 5000 cells/well. After incubating for 48 h under normoxic or hypoxic conditions, the migrated PASMCs were fixed with 4% paraformaldehyde and stained with crystal violet. A microscope (DMi8; Leica, Germany) was used to count the migrated PASMCs.
Wound-healing assay
PASMCs were seeded into six-well plates at a density of 200,000 cells/well. After the wells were overgrown with cells, the cell monolayers were scratched with 200-μL pipette tips in the center of the well, and PASMCs were incubated with the experimental treatments for 48 h. The 0 and 48 h images of PASMCs were captured using a Nikon microscope (Tokyo, Japan), and ImageJ software (NIH) was acquired to determine the level of wound closure.
Immunoblot analysis
PASMCs were lysed using a radioimmunoprecipitation assay following standard procedures after washing three times with PBS. The protein content was assayed using a bicinchoninic acid protein assay kit (Pierce Biotechnology, Rockford, IL). Subsequently, 10% sodium dodecyl sulfate–polyacrylamide gel electrophoresis was applied to separate the protein under denaturing conditions and then transferred to polyvinylidene fluoride (PVDF) membranes (Bio-Rad). After blocking for 1 h with 5% nonfat milk, the PVDF membranes were washed with Tris-buffered saline with Tween 20. Then, the membranes were incubated at 4°C overnight with rabbit primary antibodies against Vangl2 (1:500), PCNA (1:1000), MMP2 (1:1000), and β-actin (1:1000). Next, they were incubated with goat antirabbit secondary antibody (1:10000) labeled with HRP for 1 h. Bio-Rad ChemiDoc MP imaging system was used to examine the protein bands, and the quantitation was carried out using Image Lab software (Bio-Rad).
Plasmid, siRNA, mimic, and inhibitor transfection
For Vangl2 overexpression and knockdown, PASMCs were transfected with plasmid synthesized by Bio-report (Hangzhou, China) and small-interfering RNA synthesized by RiboBio (Guangzhou). PCDNA3.1 and nontargeted control siRNA (siRNA negative control) were taken as the negative control (NC). For miR-335 overexpression and knockdown, PASMCs were transfected with the mimic and inhibitor synthesized by RiboBio (Guangzhou). The sense sequences for the siRNA, mimic, and inhibitor are given in Table 1. Attractene Transfection Reagent (Qiagen, Hilden, Germany) was used for siRNA, plasmid, mimic, or inhibitor transfection.
Dual-luciferase reporter assay
The 3′-untranslational region (UTR) of Vangl2 mRNA with a putative/mutant miR-335 binding site was cloned into the pmiR-RB-ReportTM vector (Bio-report). Firefly luciferase was selected as the reporter, and Renilla luciferase was selected as the control. PASMCs were transfected with 50 nM miR-335 mimic and 100 ng pmirGLO vector with Attractene transfection reagent. Dual-Luciferase® Reporter Assay System (E1910; Promega) was used to measure luciferase activities after incubating for 48 h.
Statistical analysis
Statistical analyses and drawing were conducted using GraphPad Prism 7.0 (CA). The results are given as mean ± standard deviation. Single and multiple comparisons were conducted with Student's t-test and one-way analysis of variance, respectively, and p < 0.05 was considered statistically significant.
Results
DHA can alleviate hypoxia-induced pulmonary arterial remodeling and reverse hypoxia-induced PH in mice
To verify the protection of DHA on mice under hypoxia and the optimal concentration, different concentrations of DHA were administered to C57BL/6 mice by gavage under continuous hypoxia conditions for 3 weeks (Fig. 1A). After hypoxia, the mice showed significant elevations in RVSP, whereas the mice administered with different concentrations of DHA showed the attenuation of RVSP elevations, except 30 and 60 mg/kg (Fig. 1B). No significant difference was observed in the mCAP of each group, indicating that the effect of DHA in lowering pulmonary artery pressure was not achieved by reducing systemic arterial pressure (Fig. 1C).
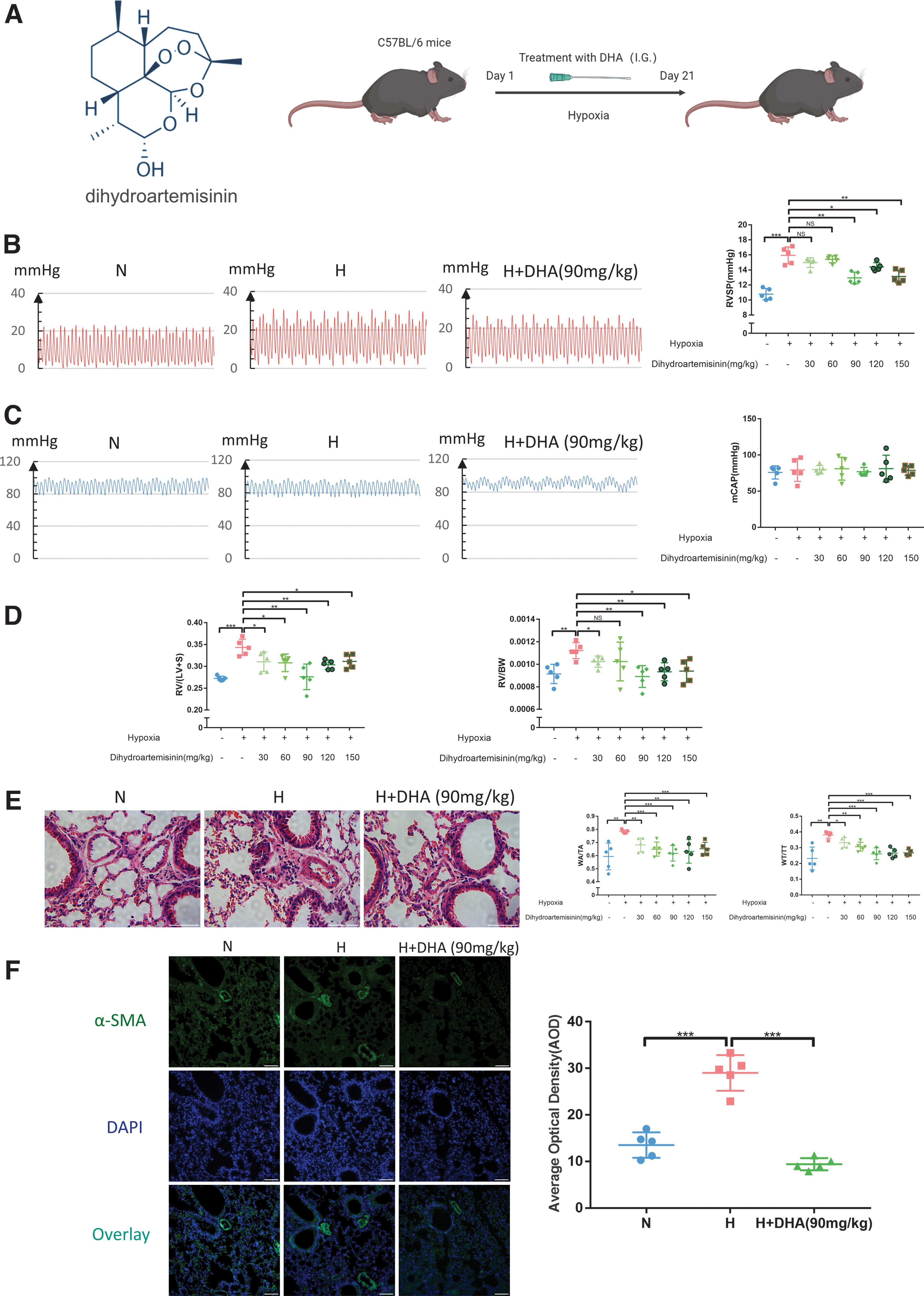
DHA can alleviate hypoxia-induced pulmonary arterial remodeling and reverse hypoxia-induced PH in mice.
As given in Figure 1D, compared with the N group, the RV/(LV + S) and RV/body weight levels strikingly increased in the H group, and DHA significantly suppressed the hypoxia-induced increase at the ventricular remodeling level. Moreover, sections stained with HE showed that hypoxia-induced pulmonary arterial remodeling was alleviated in the H + DHA group compared with the H group, and WA/TA (%) and WT/TT (%) showed the same trend (Fig. 1E). The immunofluorescence experiment also provided the same conclusion as the previous experiment (Fig. 1F). Based on the above indicators and drug dosage, the optimal drug concentration of DHA to reduce the hypoxia-induced PH of mice is 90 mg/kg.
We continued to observe the effect of DHA on hypoxic PH in rats. According to the optimal concentration of DHA in mice and the conversion of drug concentration, we set the concentration of DHA at 45 mg/kg (half of the concentration in mice), and observed the pulmonary artery pressure and pulmonary vascular remodeling in rats after drug administration. It was found that the results were consistent with mice (Supplementary Fig. S1). In conclusion, DHA can improve hypoxia-induced pulmonary remodeling and alleviate hypoxia-induced PH.
DHA treatment significantly inhibited the hypoxia-induced upregulation of miR-335 levels
To further elucidate the intrinsic mechanism of DHA in the treatment of PH, the lung tissues of N, H, and H + D (90 mg/kg) groups of mice were collected for miRNA high-throughput sequencing. After screening some conditions (Fig. 2A), 29 (16 upregulated and 13 downregulated) and 258 (153 upregulated and 105 downregulated) differentially expressed miRNAs were identified based on N group versus H group and H group versus H + D group (Fig. 2B). After analyzing the Venn diagram, 11 dif-miRNAs were identified (Fig. 2C). A total of 11 dif-miRNAs in different groups were visualized with heat maps (Fig. 2D).
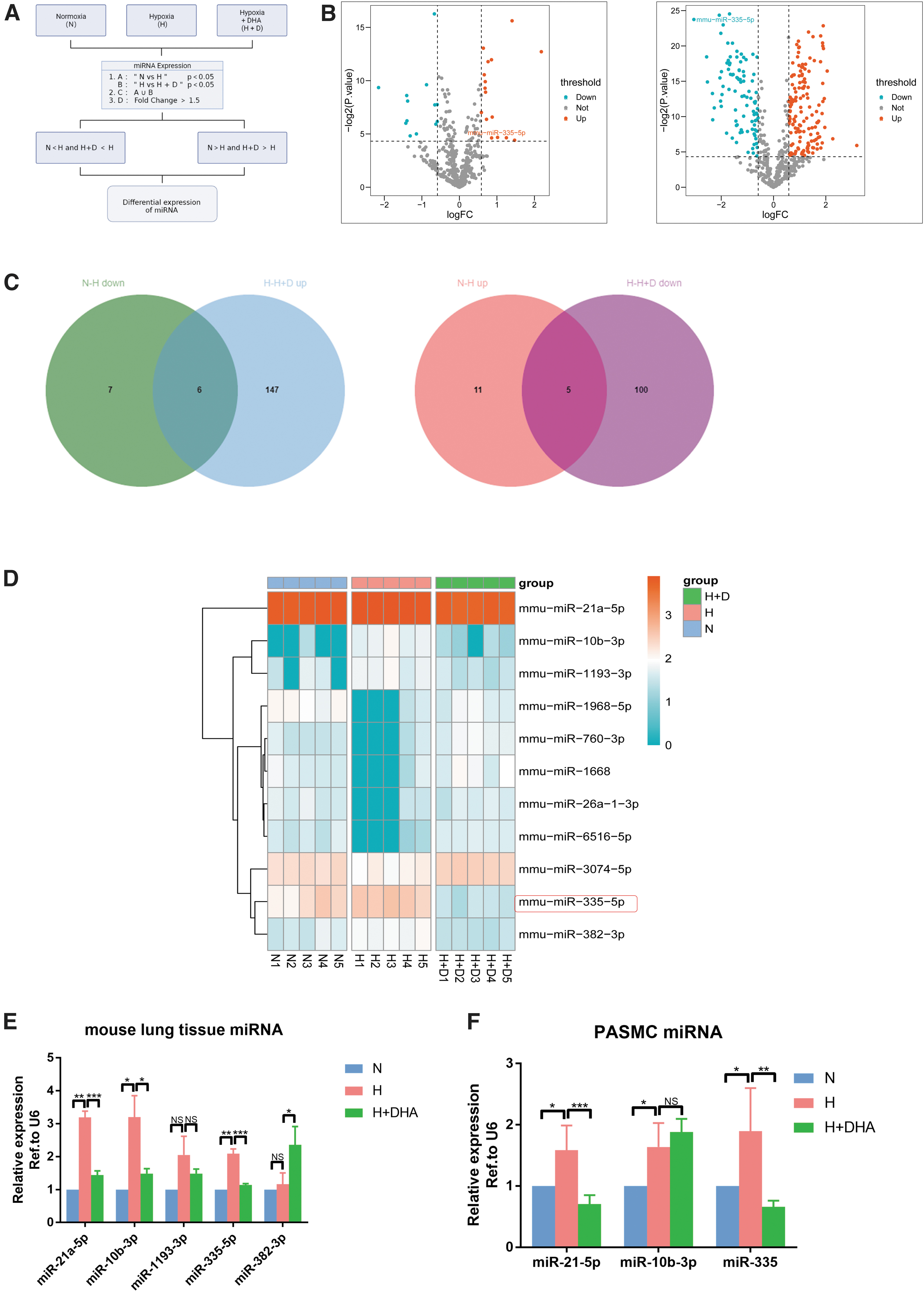
DHA treatment significantly inhibited the hypoxia-induced upregulation of miR-335 levels.
Because of the large number of miRNAs in the intersection, more attention was paid to the pathogenic miRNAs that had an increase in PH caused by hypoxia, and PCR verification was conducted on the miRNAs in the intersection. Because PASMCs play a key role in the process of pulmonary vascular remodeling, we wanted to know which miRNA expression trend in PASMC was consistent with lung tissue. Combined with miRNA sequencing results and PCR validation for lung tissue and PASMCs, it was found that among the differentially expressed miRNAs, the expressions of miR-21 and miR-335 (the previous name of miR-21a-5p and miR-335-5p) significantly increased in the H group compared with the N group, but decreased after the application of DHA compared with the H group (Fig. 2E, F). Because many studies have been reported on miR-21 and PH, we focused on the role of miR-335 in PH in this study. Earlier studies reported that the upregulation of miR-335-3p aids the induction of PH during hypoxia, so we assumed that miR-335-5p also has similar functions.
MiR-335 increases hypoxia-induced PASMC proliferation and migration
Next, we evaluated miR-335 functionality. Previously, we performed phenotypic experiments on negative controls for miR-335 mimic and inhibitor. Both of them were found to have no effect on the proliferation and migration of rat PASMCs (Supplementary Fig. S2A, C). In proliferation-related experiments (CCK8 and EdU assays), we found that miR-335 inhibitor can inhibit the high proliferation rate of cells under hypoxia (Supplementary Fig. 3A, B). Transwell and wound-healing assays showed that miR-335 inhibitor suppressed hypoxia-induced PASMC migration (Figs. 3C, D). Proteins were visualized by immunoblotting, and it was found that mir-335 inhibitor can reduce the expression of cell proliferation index PCNA and migration index MMP2 (Fig. 3E). These results indicate that miR-335 can facilitate hypoxia-induced PASMC proliferation and migration.
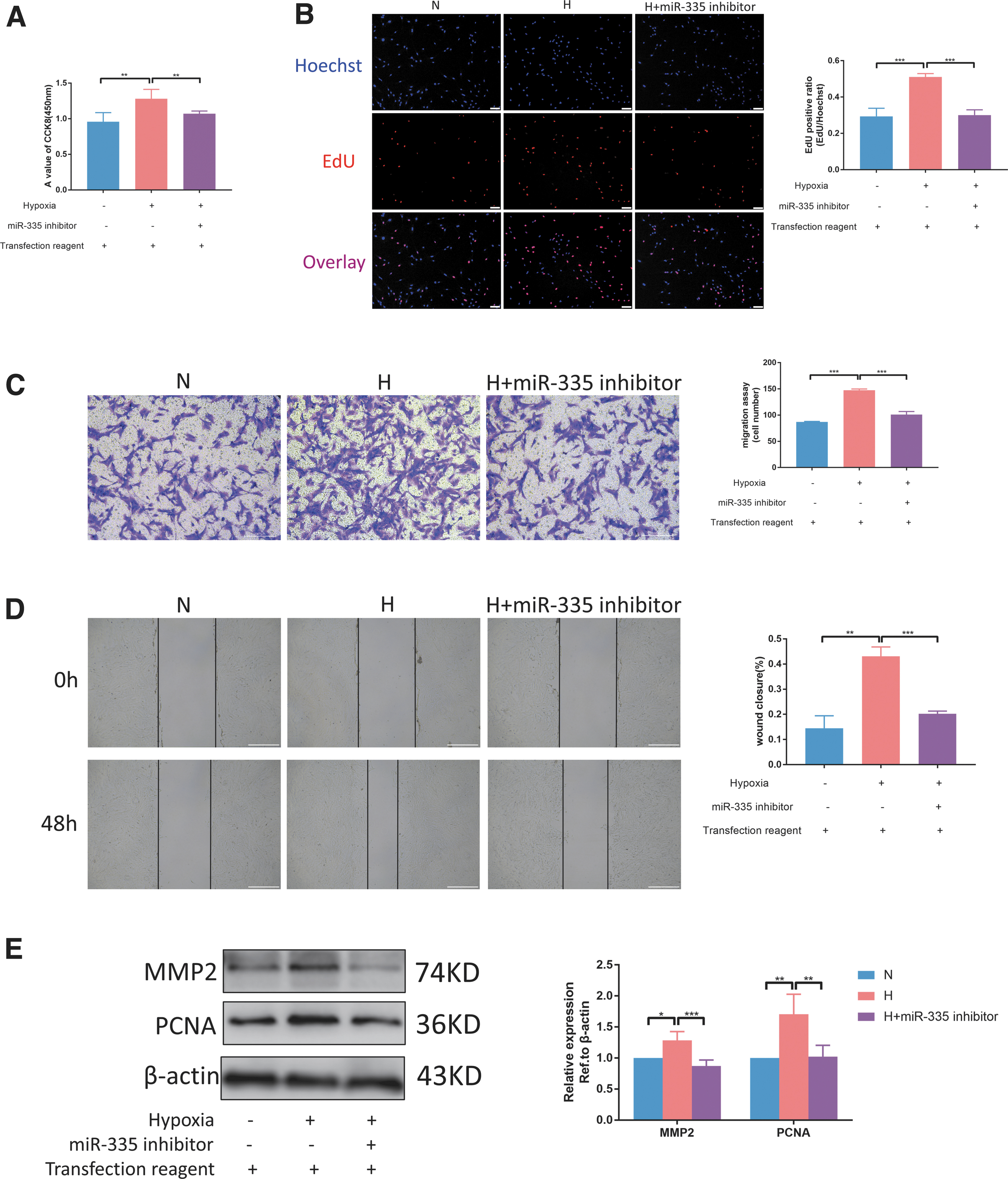
MiR-335 increases hypoxia-induced PASMC proliferation and migration.
MiR-335 can directly target Vangl2 and inhibit its expression
After studying the function of miR-335, its downstream mechanism was elucidated. It is well known that miRNA mainly negatively regulates the expression of its target genes. Therefore, three miRNA databases were selected to find the target genes of miRNA-335. Venn diagram was used to obtain the intersection of the miRNA target genes, and 22 genes in total were found (Fig. 4A, B). We focused on Vangl2, one of the PCP genes, whose mutation leads to disruption of lung development and defects in lung structure. This can result in PH, so we evaluated the role of Vangl2 in hypoxia-induced PH.
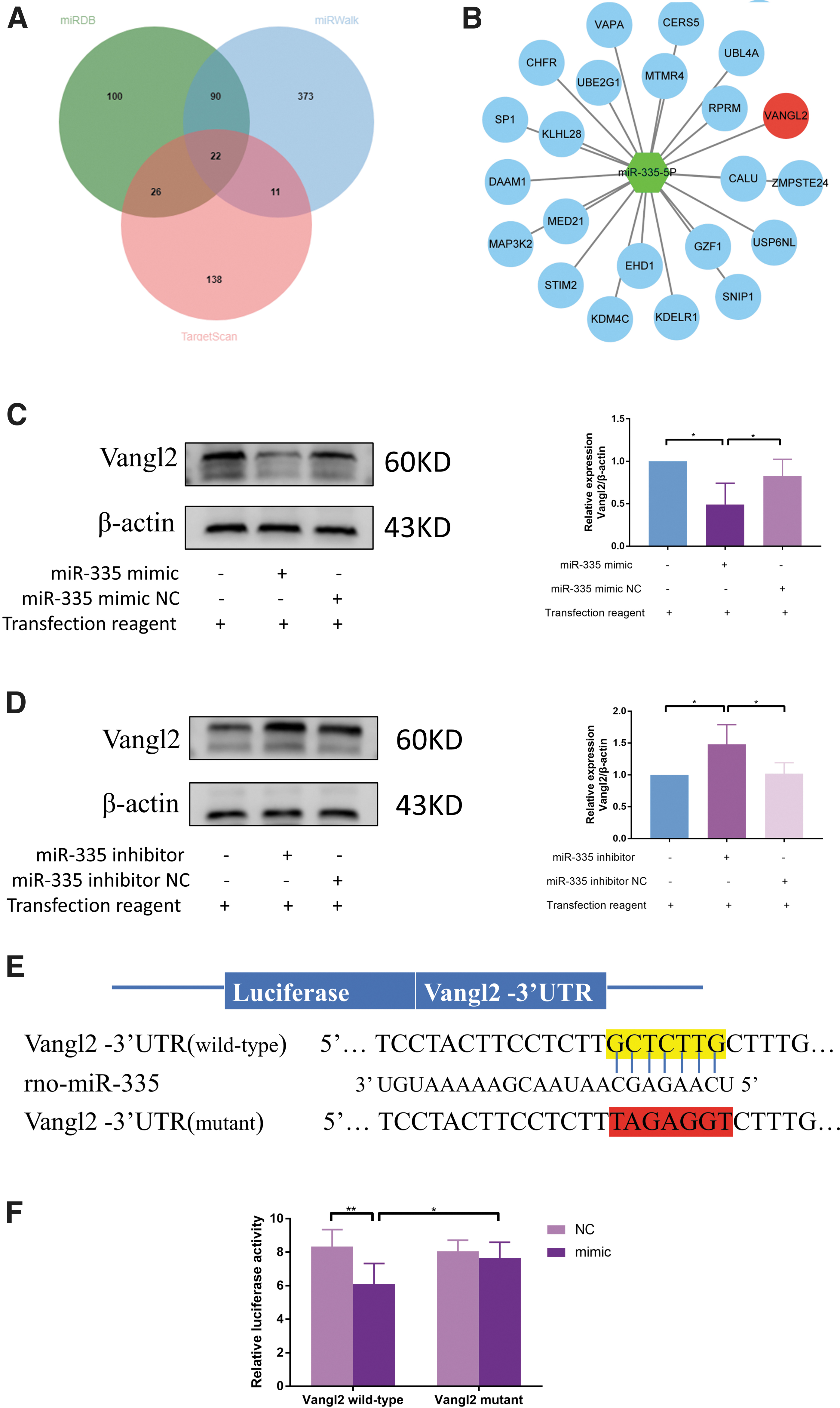
MiR-335 can directly target Vangl2 and inhibit its expression.
Before that, the targeting relationship between miR-335 and Vangl2 was first verified. Immunoblotting analyses confirmed that miR-335 inhibited Vangl2 expression (Fig. 4C). In contrast, miR-335 inhibition reversed the expression of Vangl2 in PASMCs (Fig. 4D). The transfection efficiency of miR-335 mimic and inhibitor was indirectly verified by this experiment. To further confirm this hypothesis, a luciferase reporter gene assay was performed to determine the direct interaction between miR-335 and Vangl2. The results show that the expression of Vangl2 expression was significantly decreased by miR-335 mimic transfection compared with control miRNA transfection. However, no effect was observed when the miR-335 mimic was cotransfected with the mutant Vangl2 3′-UTR (Fig. 4E, F). These results show that miR-335 can specifically target and suppress the expression of Vangl2.
Decreased Vangl2 protein expression in PH
To identify DEGs between the control group and PAH group, and after standardization of the datasets (Fig. 5A, B), 2859 DEGs were extracted from GSE53408 based on the defined criteria. The DEGs are shown in the volcano plots and the heatmaps whose clustering was performed with Euclidean distance (Fig. 5C, D), of which GSE53408 included 8 upregulated genes and 71 downregulated genes. As can be seen from the heatmap, the expression trend of Vangl2 gene in the two groups was obvious, and the expression level in the control group was higher than the PAH group. Cluster analysis was performed among the 23 samples (there were 12 patients with PH in PAH group, and 11 normal persons in control group; Fig. 5E). We found that the samples in the normal group and the PAH group were clustered into two categories, respectively, and all the patients clustered together, and also all the normal samples clustered together. The expression level of Vangl2 gene was significantly higher in the normal control group than the PAH group (Fig. 5F).
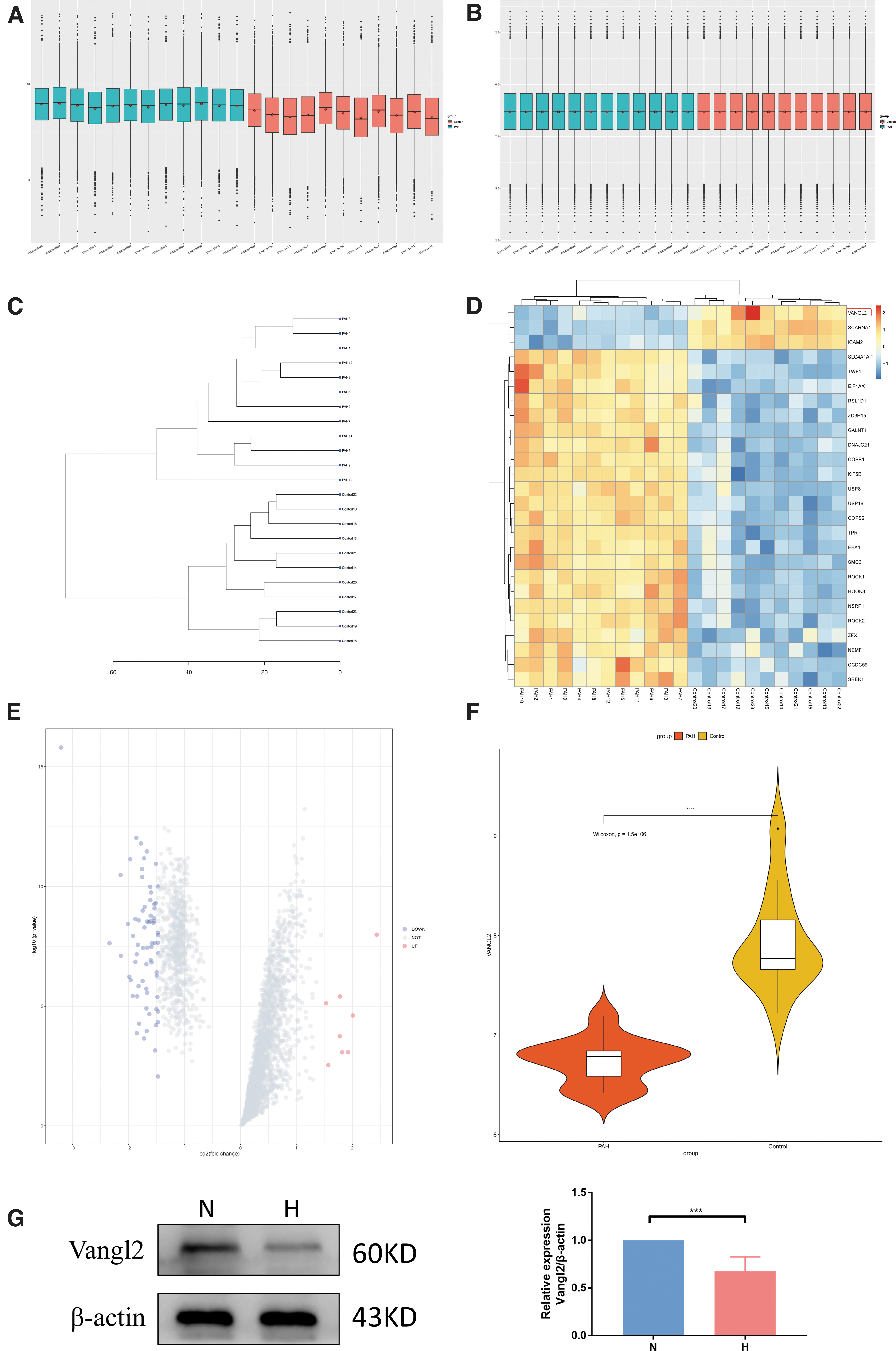
Decreased Vangl2 protein expression in pulmonary hypertension.
Next, the expression of Vangl2 under hypoxia in rat PASMC was evaluated; its expression was downregulated under hypoxia by immunoblotting (Fig. 5G).
Vangl2 decreases hypoxia-induced PASMC proliferation and migration
Immediately after, the function of Vangl2 gene was evaluated. Before the phenotypic experiment, the Vangl2 plasmid was tested, and it was indeed effective at the protein level (Fig. 6A). CCK8 and transwell experiments showed that the NC of Vangl2 plasmid and siRNA had no effect on the proliferation and migration of rat PASMCs (Supplementary Fig. S2B, D). CCK8 and EdU assays indicate that Vangl2 can inhibit hypoxia-induced PASMC proliferation (Fig. 6B, C). Transwell and wound-healing assays showed that Vangl2 suppressed hypoxia-induced PASMC migration (Fig. 6D, E). The trends of PCNA and MMP2 were also consistent with the results of previous phenotypic experiments (Fig. 6F). These results indicate that Vangl2 can facilitate hypoxia-induced PASMC proliferation and migration.
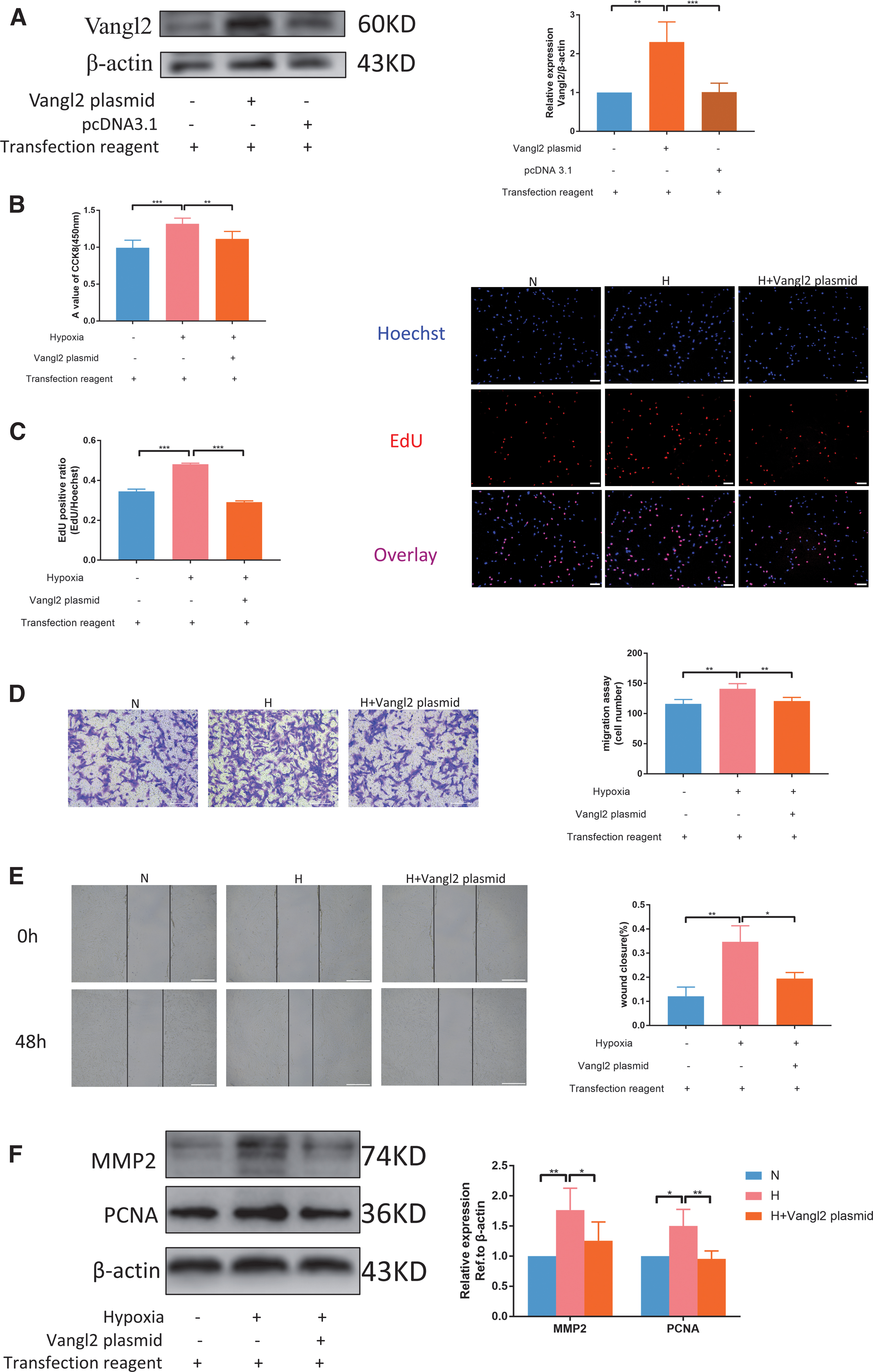
Vangl2 decreases hypoxia-induced PASMC proliferation and migration.
MiR-335 promotes hypoxia-induced PASMC proliferation and migration by downregulating Vangl2 expression
The effects of miR-335 and its target gene Vangl2 on PASMCs under hypoxia were evaluated, but whether miR-335 can affect cell proliferation and migration by downregulating Vangl2 is still not known. Before evaluating the effect of miR-335/Vangl2 pathway on cell phenotype, we verified and screened Vangl2 siRNA to be used later. From the results of the immunoblotting experiment, it can be concluded that all the three siRNAs were effective, and the second one with the highest knockdown efficiency was selected (Fig. 7A). In Vangl2 knockdown cells, the miR-335 inhibitor failed to inhibit hypoxia-induced PASMC proliferation (Fig. 7B, C).Also, the miR-335 inhibitor decreased the migration ability, which was reversed by cotransfection with Vangl2 siRNA (Fig. 7D, E). In addition, immunoblotting analysis also showed that the administration of Vangl2 siRNA reversed the low expression of PCNA and MMP2 caused by miR-335 inhibitor (Fig. 7F). These results indicate that miR-335 can increase hypoxia-induced PASMC proliferation and migration by downregulating Vangl2 expression.
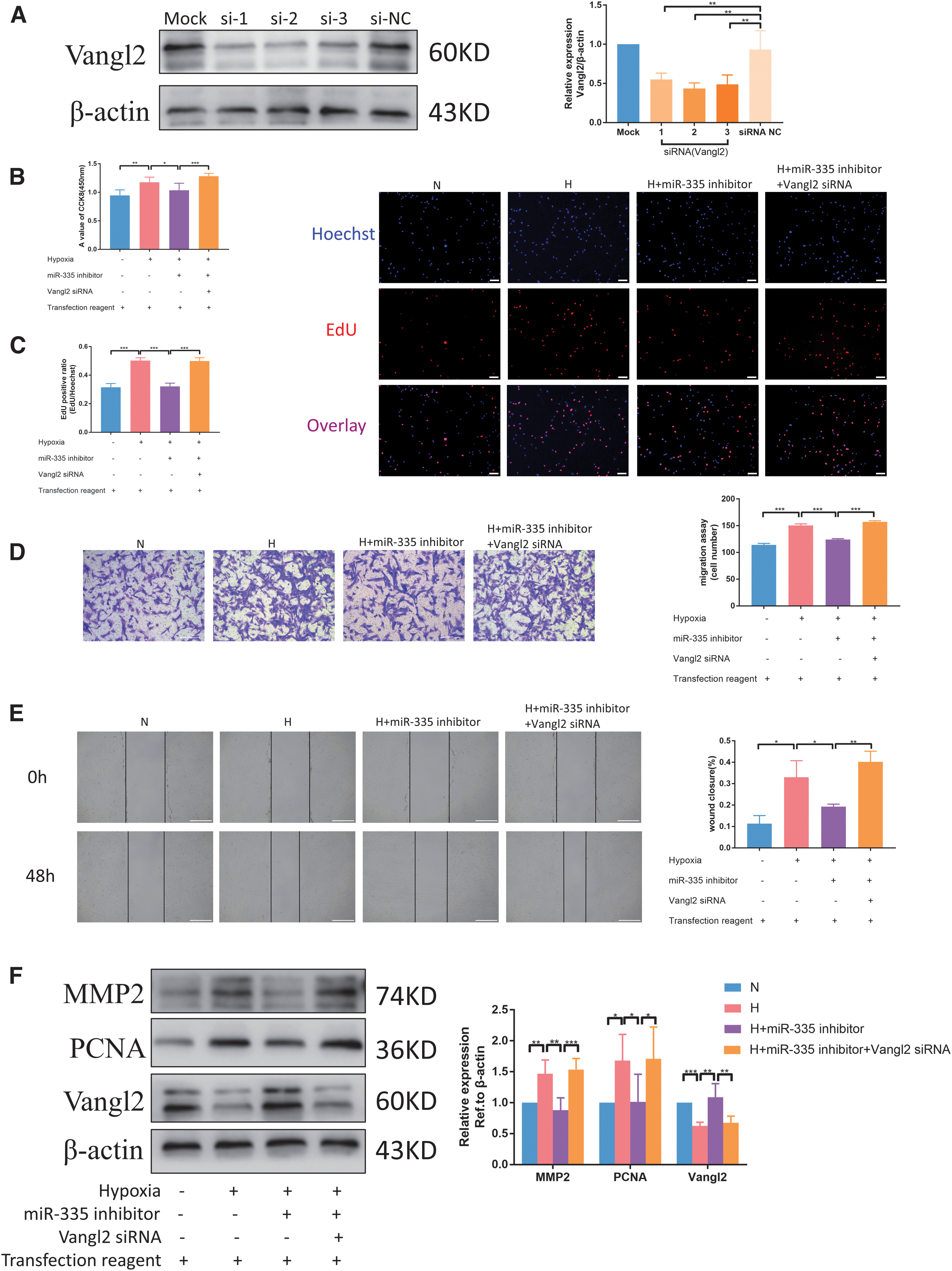
MiR-335 promotes hypoxia-induced PASMC proliferation and migration by downregulating Vangl2 expression.
DHA attenuates chronic hypoxia-induced PH by inhibiting the negative regulatory effects of miR-335 on Vangl2
After studying the miR-335/Vangl2 pathway, we returned to DHA to evaluate whether it inhibits hypoxia-induced PASMC proliferation and migration through this pathway. First, the most effective concentration of DHA on PASMCs was determined by conducting a CCK8 experiment. As given in Figure 8A, 5 μM was the optimal drug concentration of DHA to reduce hypoxia-induced PASMC proliferation. In miR-335 overexpression cells, DHA failed to inhibit hypoxia-induced PASMC proliferation (Fig. 8B, C). In addition, DHA decreased the migration ability, which was reversed by cotransfection with miR-335 mimic (Fig. 8D, E). In addition, immunoblotting analysis also revealed that the administration of miR-335 mimic reversed the high expression of Vangl2 and low expression of PCNA and MMP2 caused by DHA (Fig. 8F).
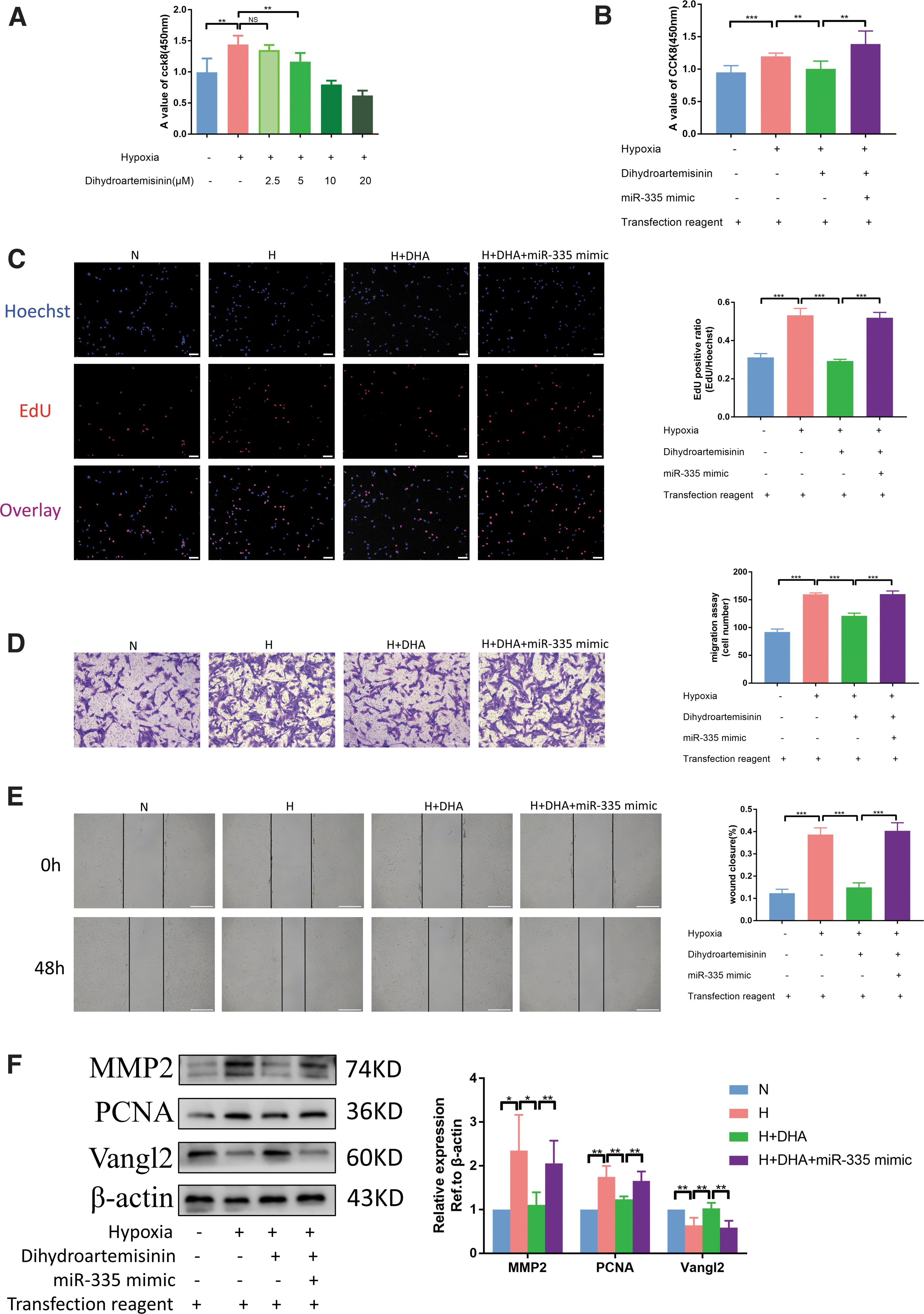
DHA attenuates chronic hypoxia-induced PH by inhibiting the negative regulatory effects of miR-335 on Vangl2.
Earlier we found that DHA treatment significantly inhibited the hypoxia-induced upregulation of miR-335 levels in mouse. We continued to observe the regulation of DHA on mouse Vangl2 by PCR experiments. It was found that the expressions of Vangl2 mRNA significantly decreased in the H group compared with the N group, but increased after the application of DHA compared with the H group (Supplementary Fig. S3). These results indicate that DHA can decrease hypoxia-induced PASMC proliferation and migration through miR-335/Vangl2 pathway.
Taken together, our results show that DHA can improve pulmonary vascular remodeling by inhibiting the negative regulatory effects of miR-335 on Vangl2 and ultimately alleviate PH (Fig. 9).

Signaling pathways in this experiment. DHA displayed a protective role on PASMCs under hypoxia by inhibiting the negative regulatory effects of miR-335 on Vangl2.
Discussion
PH is a rare but fatal disease caused by several pathological changes in the pulmonary vascular system. PH is characterized by vasoconstriction, excessive secretion of substances that promote proliferation and migration, as well as vasodilation and impaired release of antiproliferation and migration molecules (Tajsic and Morrell, 2011). Although studies on the pathophysiology of PH have made significant progress in recent years, the progress of PH therapy is not adequate. Therefore, it is essential to develop new effective therapies for other molecular pathways in the pathogenesis of PH to supplement the current treatment options.
DHA is an artificial semisynthetic derivative obtained after the reduction of artemisinin; its antimalarial activity is much better than artemisinin (Tu, 1999). In addition to treating malaria, DHA has been widely evaluated to determine its antitumor activity because of its low side effects, high efficiency, and certain specificity (Dai et al., 2021). Some studies on DHA have been reported in the cardiovascular field, but not many. Studies have shown that DHA reverses experimental PH by relieving pulmonary vascular remodeling in rats (Tang et al., 2020). In this study, we detected RVSP using right heart catheterization, and used HE staining and immunofluorescence to reflect the degree of pulmonary vascular thickening. It was found that DHA could ameliorate pulmonary arterial remodeling and reduce pulmonary artery pressure in mice.
From the results of statistical analysis, 90, 120, and 150 mg/kg concentrations of DHA can reverse hypoxia-induced PH in mice. In Figure 1B, 120 mg/kg concentration was less effective than 150 mg/kg. But in Figure 1D, the result was reversed. Owing to factors such as individual differences in mice, it is difficult to know which of the two concentrations is more effective from these experiments. But after comprehensively considering the toxicity of the drug and therapeutic effect, we believe that 90 mg/kg is the relatively best dose in vivo in alleviating hypoxic PH in mice. DHA was applied to rats under hypoxia, and the same results were obtained as mice. Unlike existing studies and our previous study, we selected a hypoxia-induced mouse model in vivo and rat PASMC model in vitro, but not MCT-induced rat PAH model and human pulmonary arterial endothelial cell model.
After Dicer digestion and processing of the single-stranded RNA precursor (pre-miRNA) with a hairpin structure of ∼70 bases in size, it becomes a mature miRNA with a length of ∼20–24 nt (Woldemariam et al., 2019). It can guide the silencing complex to hinder its translation or degrade mRNA by base-pairing interactions with the target gene mRNA (Mori et al., 2019). Nowadays, extensive studies have been conducted on miRNA and PH, so this study mainly focuses on whether DHA regulates miRNA to relieve hypoxic PH. Because the regulation of miRNA by DHA has not been previously studied in hypoxic PH, this entry point is also our innovative point for elucidating the mechanism of DHA.
Through the miRNA sequencing of the three groups of N, H, and H + DHA, and verified by PCR experiments, we found that the expression of miR-21 and miR-335 increased in the H group and decreased after the application of DHA. Owing to individual differences in mice and the species and conditions of cell modeling, only miR-21 and miR-335 satisfy the requirements of our research. There are many related reports between miR-21 and PH at present (Parikh et al., 2012; Green et al., 2017; Chang et al., 2021), so for the innovation of this study, we did not conduct further research on the mechanism of miR-21 in PH, but paid more attention to the role of miR-335.
It has been shown that blocking miR-335-3p could attenuate hypoxic PH in mice. The two arms of some miRNA precursors produce a functional mature miRNA, targeting different sites, and they are generally named and distinguished by “−5p” and “−3p.” Because both are derived from the same miRNA precursor, we speculated that miR-335-5p might be the same as miR-335-3p, which can promote PH. We used CCK8, EdU, and western blotting to assess PASMC proliferation, and PASMC migration was detected using transwell migration chamber and wound-healing assays. We found that inhibiting miR-335 could decrease hypoxia-induced PASMC proliferation and migration. Thus, we inferred that miR-335 can be categorized as a pathogenic molecule in hypoxic PH, which is just the opposite of its role in some tumor diseases (Ye et al., 2021). In our view, the cause of this phenomenon might be related to different disease models.
Vangl2 is the vertebrate counterpart of Drosophila melanogaster Vang Gogh/Strabismus protein, which was discovered 20 years ago owing to its core role in PCP. Studies have shown that a low expression of Vangl2 is associated with the occurrence of some cardiovascular diseases (Phillips et al., 2007; Yuan et al., 2014; Duan et al., 2018). Poobalasingam et al. (2017) reported that the disruption of Vangl2 can affect lung homeostasis and repair in adults. Additional studies have reported that PCP activation can prevent vessel loss in PAH (Yuan et al., 2015). Three miRNA databases were used to predict the downstream target genes of miR-335, and Vangl2 was found to be one of the target genes of miR-335. Through western blotting and dual luciferase reporter assay, we found that miR-335 can target Vangl2 and downregulate its expression.
Through the datasets and western blotting, we found that the expression level of Vangl2 gene was significantly lower in PH group than normal group. To evaluate the effect of Vangl2 on PASMC under hypoxia and the downstream mechanism of miR-335, we regulated Vangl2 and set up a rescue group based on the regulation of miR-335, and paid attention to the functions of Vangl2 at the two levels of cell proliferation and migration. As a result, it was found that Vangl2 reduces hypoxia-induced PASMC proliferation and migration, and the miR-335 inhibitor fails to inhibit hypoxia-induced PASMC proliferation and migration when cotransfected with siVangl2.
Therefore, we emphasize that Vangl2 plays a certain protective role in hypoxic PH, and miR-335 regulates hypoxic PH by targeting Vangl2. Before investigating whether DHA alleviates hypoxic PH by regulating the miR-335/Vangl2 pathway, we determined the optimal concentration of DHA to act on PASMCs by CCK8 assay. Subsequently, a rescue experiment was set up for DHA treatment, and it was concluded that DHA can improve pulmonary vascular remodeling by inhibiting the negative regulatory effects of miR-335 on Vangl2 and ultimately alleviate PH.
Through the overall review of our research and comparison with a previous study, some relatively innovative views were found. First of all, increasing studies have focused on the pharmacological mechanism of DHA, but the relationship between DHA and miRNA has not been evaluated. Although related studies have been reported on the treatment of PH by DHA, we report for the first time that DHA can alleviate PH by downregulating miR-335. Next, the phenotypic study between miR-335 and PH is also a novelty of this study, such as Vangl2 and PH. Finally, a joint regulation of two potential targets was executed to determine the connection between miR-335/Vangl2 pathway and PH.
However, this study also has several weaknesses. To begin with, we did not design a regulation experiment on miR-335 in vivo. Besides, we only studied the phenotypic regulation of Vangl2, but did not involve its downstream pathways. Otherwise, in RNA sequencing we did not normalize with PASMC markers, so single-cell sequencing of lung tissue may be a better approach. The heart is also affected in PH, so it will be interesting to see whether DHA can alleviate heart PH-related phenotype in future studies. Because human PASMCs are not involved in this experiment, the connection between human and rat PASMCs is not exactly described in this article. However, both human and rat PASMCs proliferate and migrate abnormally under hypoxia. Human and rat miR-335 sequences are identical (Table 2), and Vangl2 expression is also reduced in PH. Therefore, we speculate that the miR-335/Vangl2 pathway may be involved in the process of human hypoxic PH.
The Sequence of miR-335
miRNA, microRNA.
Combined with the previous evidence, our findings in this study show that miR-335 targets Vangl2 to form a regulatory axis and participate in the remission of PH by DHA.
Conclusions
MiR-335 expression is increased in hypoxic PH, which can significantly promote hypoxia-induced proliferation and migration of PASMCs. Vangl2 is a potentially beneficial therapeutic target in hypoxic PH, and the regulation of PASMCs phenotype by miR-335 can be achieved by targeting Vangl2. DHA effectively reverses the hypoxia-induced upregulation of miR-335 expression, relieving the miR-335-mediated downregulation of Vangl2 and thereby promoting Vangl2 expression to prevent PH. Our study proves the important and different roles of miR-335 and Vangl2 in the proliferation and migration of PASMCs induced by hypoxia, and we found a novel mechanism that DHA can attenuate hypoxia-induced PH by inhibiting the negative regulatory effects of miR-335 on Vangl2 for the first time. This research provides new insights and thoughts on the role of miRNA in PH and the application of DHA. Based on the similarity in function and morphology of human and rat PASMCs and the conservation of miRNAs, we have reasons to believe the role of miR-335/Vangl2 pathway in human PH and the pharmacological effects of DHA.
Footnotes
Acknowledgment
The authors thank the Key Laboratory of Heart and Lung of Wenzhou Medical University for support for this study.
Authors' Contributions
Y.L., H.C., X.H., and L.W. designed experiments and wrote the article. Y.L., H.C., J.W., and L.Z. conducted the experiments. Y.Y., M.X., L.S., and C.Z. collected the data and participated in the statistical analyses. All authors read and approved the final version of the article.
Availability of Data and Material
All data are contained within the article. The original data supporting the conclusions of this study are available from the authors.
Ethics Approval
All animal procedures performed conformed to the guidelines from Directive 2010/63/EU of the European Parliament on the conservation of animals used for scientific aims or the current NIH guidelines and were approved by the Animal Ethics Committee of Wenzhou Medical University. In addition, during the study, all animals were carefully handled and killed humanely.
Disclosure Statement
No competing financial interests exist.
Funding Information
This work was supported by the Chinese National Natural Science Foundation Grant (No. 91839106).
Supplementary Material
Supplementary Figure S1
Supplementary Figure S2
Supplementary Figure S3
References
Supplementary Material
Please find the following supplemental material available below.
For Open Access articles published under a Creative Commons License, all supplemental material carries the same license as the article it is associated with.
For non-Open Access articles published, all supplemental material carries a non-exclusive license, and permission requests for re-use of supplemental material or any part of supplemental material shall be sent directly to the copyright owner as specified in the copyright notice associated with the article.