
Abstract
Osteoarthritis (OA) is a chronic joint disease characterized by the deterioration of cartilage and subchondral bone in the joints. Currently, there is no complete cure for OA, only treatments designed to temporarily relieve pain and improve function. Compared with the high cost of surgical treatment, medical treatment of OA is more acceptable and cost-effective. Rutin, as a flavonoid, has been shown to have anti-OA properties. We evaluated the effects of rutin on chondrocytes in lipopolysaccharide (LPS)-induced OA and on OA in rats induced by anterior cruciate ligament transection. We found that rutin effectively reduced the expression levels of inducible nitric oxide synthase (iNOS), cyclooxygenase-2 (COX-2), tumor necrosis factor-α (TNF-α), and matrix metalloproteinase 13 (MMP-13) and increased the expression of Col II and aggrecan (p < 0.001). In addition, we also found that rutin increased the expression of cystathionine-β-synthase (CBS) and inhibited the expression of Rho-related coiled-coil protein kinase (ROCK) in chondrocytes (p < 0.05), thereby effectively inhibiting the inflammatory progression of OA. We concluded that rutin inhibits the inflammatory progression of OA through the CBS-mediated RhoA/ROCK signaling pathway.
Introduction
Osteoarthritis (OA) is a highly prevalent degenerative joint disease, affecting 250 million people worldwide (GBD 2017 Risk Factor Collaborators, 2018). With the aging of the global population, the rise in obesity and the increase in the number of joint injuries, OA has become more and more common (Rosen et al., 2020). OA is characterized by local inflammation and chronic strain of the joints. As time progresses, joint function gradually declines, eventually leading to disability (Wan et al., 2021). The current treatment for OA is mainly through artificial joint replacement surgery, but surgical treatment is more expensive, and there will inevitably be incidences of infections around the prosthesis, refractures around the prosthesis, vascular and nerve damage, and other complications (De Martino et al., 2019; Khan et al., 2021).
Nonsteroidal anti-inflammatory drugs, cyclooxygenase-2 (COX-2) inhibitors, opioids, and other drug treatments are designed to relieve pain and control symptoms, but they can also cause gastrointestinal and cardiovascular adverse reactions (Bahmani et al., 2017; Mitchell et al., 2021). Recent studies have shown that natural plant flavonoids can reduce the risk of cardiovascular disease by tightly binding to the peroxidase active site of COX-2 and acting as a natural cofactor of COX-2 (Chen et al., 2020). In view of the increasing incidence of OA in the future, and to further improve the efficacy of drugs in the treatment of OA, it is necessary to explore and develop new drugs for the treatment of OA.
Rutin is a flavonoid compound, also known as rutoside and quercetin-3-rutin. Chemically speaking, it is a glycoside containing quercetin and disaccharide sugar (Hu et al., 2021). According to a large number of studies, rutin participates in various pharmacologically active mechanisms such as antiarthritis, antioxidation, anticancer, blood sugar control, neuroprotection, cardioprotection, liver protection, and kidney protection (Ganeshpurkar and Saluja, 2017).
Guardia et al. (2001) found that in an adjuvant arthritis rat model, rutin inhibits the acute and chronic phases of inflammation and is most active in the chronic phase of inflammation. Horcajada et al. (2015) confirmed that rutin slows down inflammation and catabolizes cartilage markers in OA lesions in Hartley guinea pigs. Chen et al. (2021) verified that rutin inhibits the inflammatory response and extracellular matrix degeneration stimulated by advanced glycation end products in a mouse model of OA by targeting TRAF-6 and BCL-2 proteins. Based on these observations, rutin has great potential in the treatment of OA, and further research into the pharmacological mechanisms involved in its effects is needed.
Wang et al. (2016) found that rutin can inhibit the RhoA/ROCK signaling pathway by reducing reactive oxygen species, thereby significantly preventing hyperglycemia from destroying the renal endothelial barrier function. Rho-related coiled-coil protein kinase (ROCK) is a downstream effector of the small GTP binding protein RhoA, and has two homologous isomers: ROCK1 and ROCK2 (Quadir et al., 2021). ROCK1 and ROCK2 are expressed during cartilage differentiation of primary cells and ATDC5 (chondrocyte line derived from mouse teratocarcinoma) cells (Wang et al., 2004). Interleukin (IL)-1β, IL-6, transforming growth factor-α (TGF-α), and other inflammatory factors play a key role in promoting the progression of OA. TGF-α activates the RhoA/ROCK signaling pathway to induce degradation of articular cartilage (Appleton et al., 2010).
RhoA/ROCK signaling pathway can also be regulated by the endogenous gas H2S catalyzed by the enzyme (Zhang et al., 2021a, 2021b). The production of endogenous H2S is mainly catalyzed by the enzymatic pathway through cystathionine-β-synthase (CBS), cystathionine-γ-lyase (CSE), and 3-mercaptopyruvate sulfur transferase (3-MST) (Paul et al., 2021). Burguera et al. found that the biosynthesis of endogenous H2S gas was reduced in OA cartilage, but there was no difference in serum H2S concentration between OA patients and control donors. in addition, the differential expression of CBS was significantly greater than that of CSE, which could indicate that in OA, the activity of CBS in cartilage is stronger than CSE (Burguera et al., 2020).
Based on previous research, this study aimed to establish an lipopolysaccharide (LPS)-induced ATDC5 chondrocyte model and a rat OA model to explore whether rutin can act on CBS and consequently affect the RhoA/ROCK signaling pathway to achieve a therapeutic effect in OA.
Materials and Methods
CBS, ROCK bioinformatics analysis
Protein–protein interaction network construction
The protein–protein interaction network of CBS and ROCK was created by the search tool used to search the online database of interacting genes/proteins (STRING) (
Cultivation of ADTC5 chondrocytes
The mouse chondrocyte cell line ATDC5 was obtained from the American Type Culture Collection (ATCC, Manassas, VA). ATDC5 cells were cultured in DMEM/F12 medium (Gibco, Carlsbad, CA), supplemented with 10% fetal bovine serum (Gibco) and 2 mM L-glutamine (Gibco), 100 U/mL penicillin, and 100 μg/mL streptomycin (Gibco) in an incubator at 37°C under 5% CO2. According to previous references, ATDC5 cells were treated with 5 μg/mL LPS (Sigma, St. Louis, MO) for 24 h to establish an in vitro model of OA (Wu et al., 2018).
Cell transfection
We synthesized the CBS overexpressing plasmid pcDNA3.1 (GenePharma, Shanghai, China), siRNA (GenePharma) and negative control (Blank) (GenePharma). According to the manufacturer's protocol, all cells were transfected with lipofectamine® 3000 reagent (Invitrogen, Thermo Fisher Scientific, Waltham, MA). Cells were cultured in an incubator at 37°C and 5% CO2 for 24 h, and then real-time quantitative reverse transcription polymerase chain reaction (RT-qPCR) was used to analyze cell transfection efficiency.
Cell viability determination
Cell viability was evaluated using a Cell Counting Kit-8 (CCK-8; Dojindo Molecular Technologies, Gaithersburg, MD). ATDC5 cells were seeded into 96-well culture dishes, at a density of 5 × 103 cells per well and treated with different concentrations of LPS (0, 2, 5, 10, 20, or 50 μg/mL) for 12 h. After stimulation, CCK-8 solution (10 μL/well) was added to the culture medium. Then, the cells were incubated at 37°C under 5% CO2 for 2 h, before the absorbance was measured at 450 nm using a microplate reader (Bio-Rad, Hercules, CA). According to the results, 5 μg/mL LPS was selected for subsequent experiments.
Hematoxylin and eosin staining
OA knee cartilage tissue was obtained from OA patients, and normal knee cartilage tissue was obtained from patients who underwent amputations due to trauma. The knee cartilage was decalcified in 18% EDTA, fixed in 4% paraformaldehyde, dehydrated in ethanol, and embedded in paraffin, then sectioned at a thickness of 10 μm. Finally, the sections were stained with hematoxylin and eosin (HE). All images were captured under a light microscope (Zeiss, AXIO; Carl Zeiss Microscopy GmbH, Jena, Germany).
Western blotting
Total protein was extracted in radioimmunoprecipitation assay (RIPA) lysis buffer (P0013B; Beyotime Institute of Biotechnology, Jiangsu, China), and the concentration was measured using a BCA kit. Total protein was separated by sodium dodecyl sulfate-polyacrylamide gel electrophoresis (SDS-PAGE) and transferred to a polyvinylidene difluoride (PVDF) membrane. The primary anti-rabbit antibodies against CBS, CSE, 3-MST, ROCK1, ROCK2, inducible nitric oxide synthase (iNOS), COX-2, tumor necrosis factor-α (TNF-α), Col II, Aggrecan, and matrix metalloproteinase 13 (MMP-13) were purchased from Abcam (Cambridge, MA), and β-actin antibody was purchased from Sigma-Aldrich (St. Louis, MO). The PVDF membranes were blocked in 5% nonfat milk, then placed in primary antibody dilution buffer (1:1000) and incubated together overnight at 4°C.
After washing with Tris-HCl, NaCl and Tween20 buffer at room temperature, they were incubated with horseradish peroxidase-conjugated anti-rabbit IgG secondary antibody diluted at 1:1000 (Cell Signaling Technology, Danvers, MA) for 1 h. Finally, an ECL kit (Thermo Fisher Scientific) was used to observe the immunostained bands, and ImageJ 6.0 software (NIH, Bethesda, MD) was used to calculate the results.
Immunofluorescence
ATDC5 cells were inoculated into wells of a 24-well plate (5 × 104/per well) and allowed to settle overnight. The cells were then prefixed with 4% paraformaldehyde for 10 min, then fixed in precooled methanol for another 10 min. Then, the cells were incubated at 4°C overnight with the following main antibodies: anti-iNOS, anti-Col II, anti-Aggrecan, and anti-MMP-13 (all at 1:1000; Abcam). Next day, the cells were incubated with goat anti-rabbit IgG secondary antibody (1:5000; Abcam). The nuclei were counterstained with DAPI (Beyotime) for 5 min, then the cells were observed under a CX23 fluorescence microscope (Olympus Corporation, Tokyo, Japan). Each group was photographed three times, and ImageJ 6.0 software (NIH) was used to calculate the results.
Real-time PCR
According to the manufacturer's protocol, total RNA was extracted from the cell line using TRIzol® reagent (Thermo Fisher Scientific). Total RNA was reverse transcribed into cDNA using a PrimeScript RT kit (Takara Bio, Inc., Shiga, Japan). Then, SYBR premix was used and RT-qPCR was performed using Ex Taq II kit (Takara). Real-time qPCR was performed according to the following protocol in triplicate: 2 min at 94°C, then 35 cycles (30 s at 94°C; 45 s at 55°C). The formula 2−△△ct was used to calculate the relative gene expression level as a fold change, and β-actin was used as an internal reference. All primers used in this study are shown in Table 1.
Sequences of Primers Used for Gene Amplification
3-MST, 3-mercaptopyruvate sulfur transferase; CBS, cystathionine-β-synthase; COX-2, cyclooxygenase-2; CSE, cystathionine-γ-lyase; iNOS, inducible nitric oxide synthase; MMP-13, matrix metalloproteinase 13; ROCK, Rho-related coiled-coil protein kinase; TNF-α, tumor necrosis factor-α.
Animals
A total of 21 Sprague–Dawley (SD) rats, weighing 180–200 g, 4–5 weeks old, were purchased from the Experimental Animal Center of Anhui Medical University (Hefei, China). The animal care and laboratory animal use protocol were approved by the Ethics Committee of the First Affiliated Hospital of Anhui Medical University. The ethical approval reference number for this study is 20200906. After adaptive feeding for 1 week, the rats were randomly divided into three groups: Control group (n = 7, injection of 0.9% saline into the joint cavity, 400 μL), OA group (n = 7, intra-articular injection of 0.9% saline, 400 μL), and OA+rutin group (n = 7, intra-articular injection of 10 mg/mL rutin, 400 μL).
To establish an in vivo model of OA, rats were anesthetized by intraperitoneal injection of 2% pentobarbital (40 mg/kg), and then the right knee joint capsule was incised inside the patellar tendon. After that, the anterior cruciate ligament was cut off using microsurgery. Twenty-four hours after operation, rats in the OA+rutin group were injected with rutin into the joint cavity three times a week. All experimental animals were sacrificed 6 weeks after the operation. Then, the knee joint tissues of the rats were collected for follow-up experiments.
Statistical analysis
All data are expressed as mean ± standard deviation (x ± SD) or repeated value. Statistical analysis was performed using GraphPad Prism 8.0 (GraphPad Software, Inc., San Diego, CA). Each experiment was repeated three times. The comparison between two groups was performed by t-test, and the comparison among multiple groups was evaluated by one-way analysis of variance (ANOVA). p < 0.05 is considered to indicate a significant difference.
Results
CBS, ROCK bioinformatics data analysis
Searches through the STRING online database revealed that proteins related to the role of CBS are as follows: CBSL (CBS), CTH (CSE), AHCYL1, AHCYL2, BHMT2, AHCY, BHMT, SDS, MTR, SHMT1, and SHMT2 (Fig. 1A). GO enrichment function description data showed that CBS is mainly present in the cytosol, cytoplasm, and extracellular exosomes and plays the role of binding to pyridoxal phosphate and catalyzing transsulfuration of adenosine homocysteinase, and participates in the metabolism of sulfur amino acids, one-carbon metabolism, and other biological processes (Fig. 1C). KEGG pathway enrichment results showed that CBS is mainly involved in cysteine and methionine metabolism, glycine, serine, and threonine metabolism and other biological pathways (Fig. 1E).
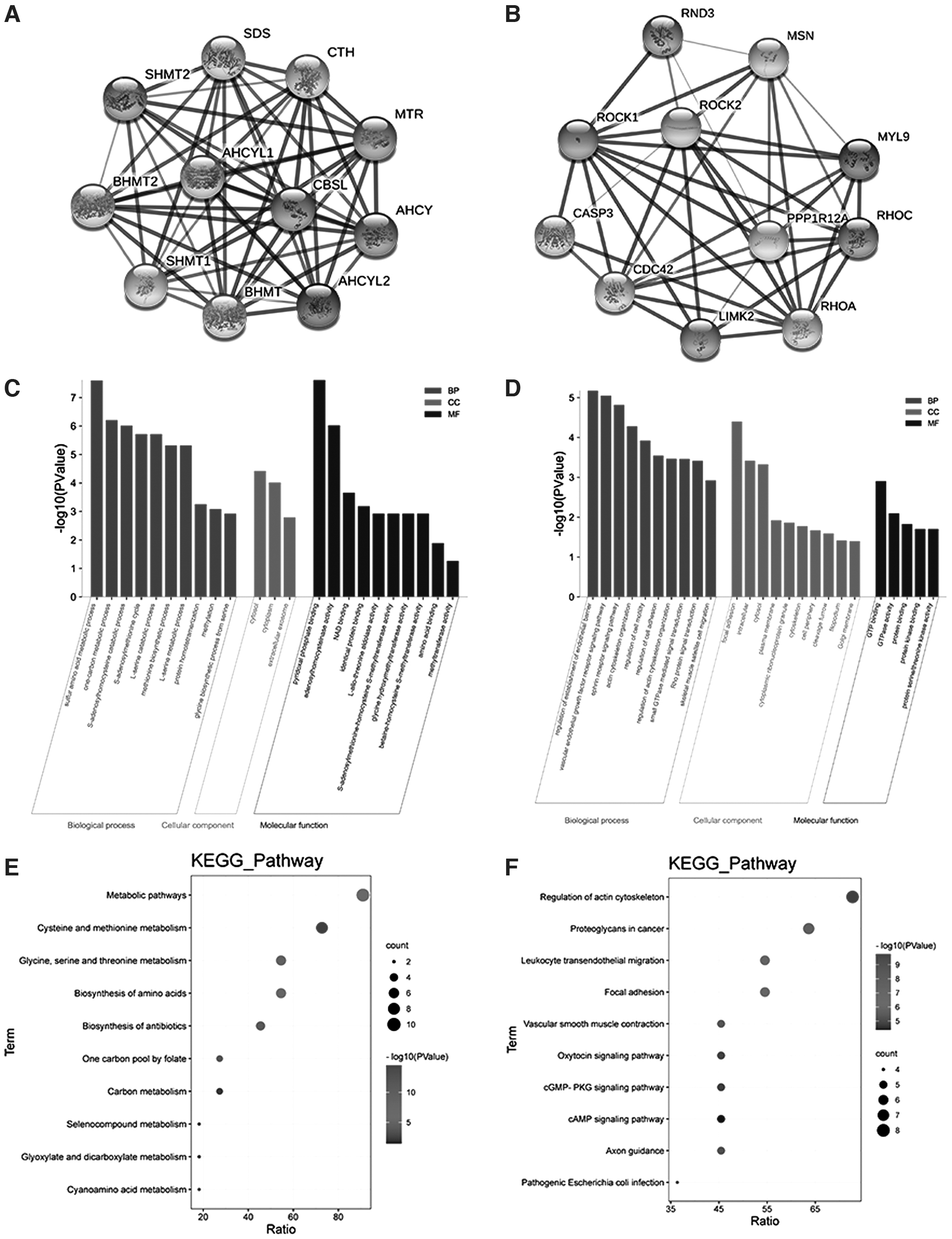
CBS and ROCK bioinformatics data analysis.
Similarly, proteins related to ROCK are as follows: ROCK1, ROCK2, RND3, MSN, MYL9, PPP1R12A, RHOA, RHOC, LIMK2, CDC42, CASP3 (Fig. 1B). The enrichment function data of GO showed that ROCK is mainly present in adhesion spots, cell cytosol, and plasma membrane, exerts the molecular function of binding to GTP and GTPase activity, and participates in biological processes such as sulfur amino acid metabolism and one-carbon metabolism (Fig. 1D). KEGG pathway enrichment results showed that CBS is mainly involved in regulation of actin cytoskeleton organization, regulation of cell movement, regulation of cell adhesion, and other biological pathways (Fig. 1F).
Expression of CBS and ROCK in OA articular cartilage
X-ray imaging showed that the inner joint space of the right knee joint of the rats was narrowed, exhibiting joint edge hyperplasia and osteophyte formation, as well as subchondral bone sclerosis (Fig. 2A). Magnetic resonance imaging showed that the articular cartilage was thinned and the joint space was narrowed. The hyperplastic bone under the articular surface exhibited a low signal on T1WI and T2WI, showing discontinuity of the cortical bone with low signal (Fig. 2B). Examination of specimens of the surgically resected knee joints showed that the articular cartilage was gray and white, and the cartilage surface was rough and there were cracks and depressions, thinning of the cartilage layer, and osteophyte formation (Fig. 2C).
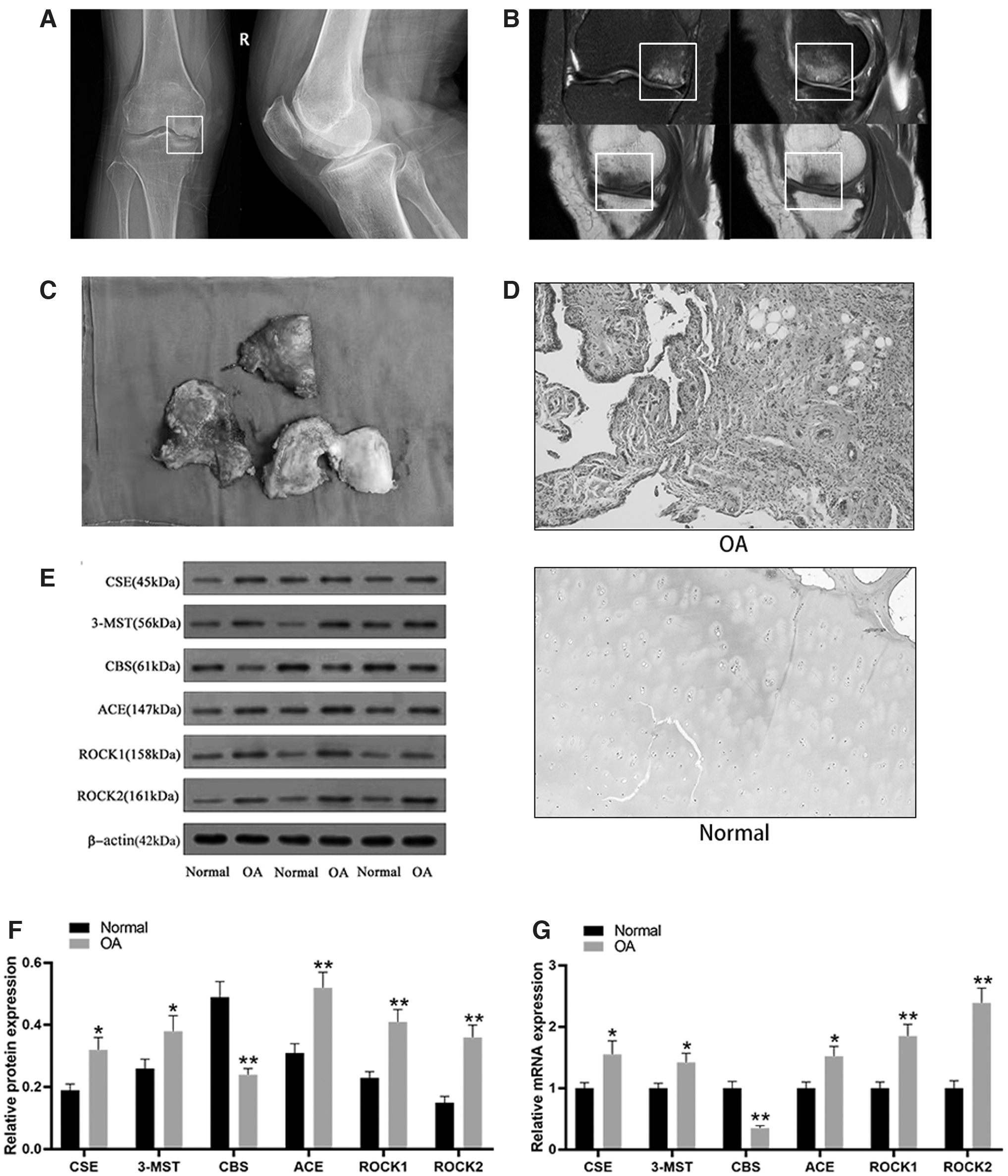
Expression of CBS and ROCK in OA articular cartilage.
HE staining of OA cartilage tissue showed cartilage tissue defects, a reduction in the number of chondrocytes, the layers were unclear, the cells were arranged in a disorderly manner, and infiltration of a large number of inflammatory cells was observed, while in normal cartilage tissue, the chondrocytes were arranged in an orderly manner, with distinct layers and normal shape (Fig. 2D). Compared with normal cartilage tissue, the expression of ROCK1 and ROCK2 proteins in OA cartilage tissue was significantly increased, while the expression of CBS protein was significantly decreased (p < 0.001) (Fig. 2E, F). In agreement with these data, the mRNA expression of ROCK1 and ROCK2 in OA cartilage tissue was also significantly increased (p < 0.001), while the expression of CBS protein was significantly decreased (p < 0.001) (Fig. 2G).
Rutin inhibits the inflammatory response of LPS-induced chondrocytes
Rutin, chemical name 3,3′,4′,5,7-pentahydroxyflavone-3-rhamnoside, molecular formula C27H30O16, is a natural flavonoid glycoside (Fig. 3A). The CCK-8 assay can measure cell viability, and the results show that as the concentration of LPS increases, the survival rate of chondrocytes gradually decreases. However, after treating chondrocytes with LPS at a concentration of 5 μg/mL, the cell survival rate remained at more than 80% (p < 0.05). Therefore, 5 μg/mL LPS was selected for follow-up experiments (Fig. 3B). Compared with the control group, the protein and mRNA expression levels of iNOS, COX-2, TNF-α, and MMP-13 in chondrocytes of the LPS-induced group were significantly increased, while the expression levels of Col II and Aggrecan were significantly decreased (p < 0.001).

Rutin inhibits the inflammatory response in LPS-induced chondrocytes.
After treatment of LPS-induced chondrocytes with different concentrations of rutin at 5, 10, or 20 μg/mL, compared with the LPS-induced group, the protein and mRNA expression levels of iNOS, COX-2, TNF-α, and MMP-13 gradually but significantly decreased as the concentration of rutin increased, while the expression levels of Col II and Aggrecan gradually and significantly increased (p < 0.001) (Fig. 3C–E). Immunofluorescence was used to detect the expression of related proteins in cells–iNOS and MMP-13 in the cytoplasm, Col II in the nucleus, and Aggrecan both in the nucleus and the cytoplasm. The expression levels of each of these followed the same trends as mentioned above for the mRNA (Fig. 3F–J). Therefore, it can be concluded that rutin effectively alleviates the inflammatory response of chondrocytes induced by LPS.
Rutin relieves the inflammatory response of LPS-induced chondrocytes by promoting the expression of CBS
Compared with the control group, the expression levels of CBS protein and mRNA in chondrocytes of the LPS-induced group were significantly reduced (p < 0.001). After treatment of LPS-induced chondrocytes with different concentrations of rutin at 5, 10, or 20 μg/mL, compared with the LPS-induced group, as the concentration of rutin increased, the protein and mRNA expression levels of CBS gradually but significantly increased (p < 0.001) (Fig. 4A–C). Compared with the control group, the protein and mRNA expression levels of iNOS, COX-2, TNF-α, and MMP-13 were significantly increased in chondrocytes of the LPS-induced group, while the expression levels of Col II and Aggrecan were significantly decreased (p < 0.001).
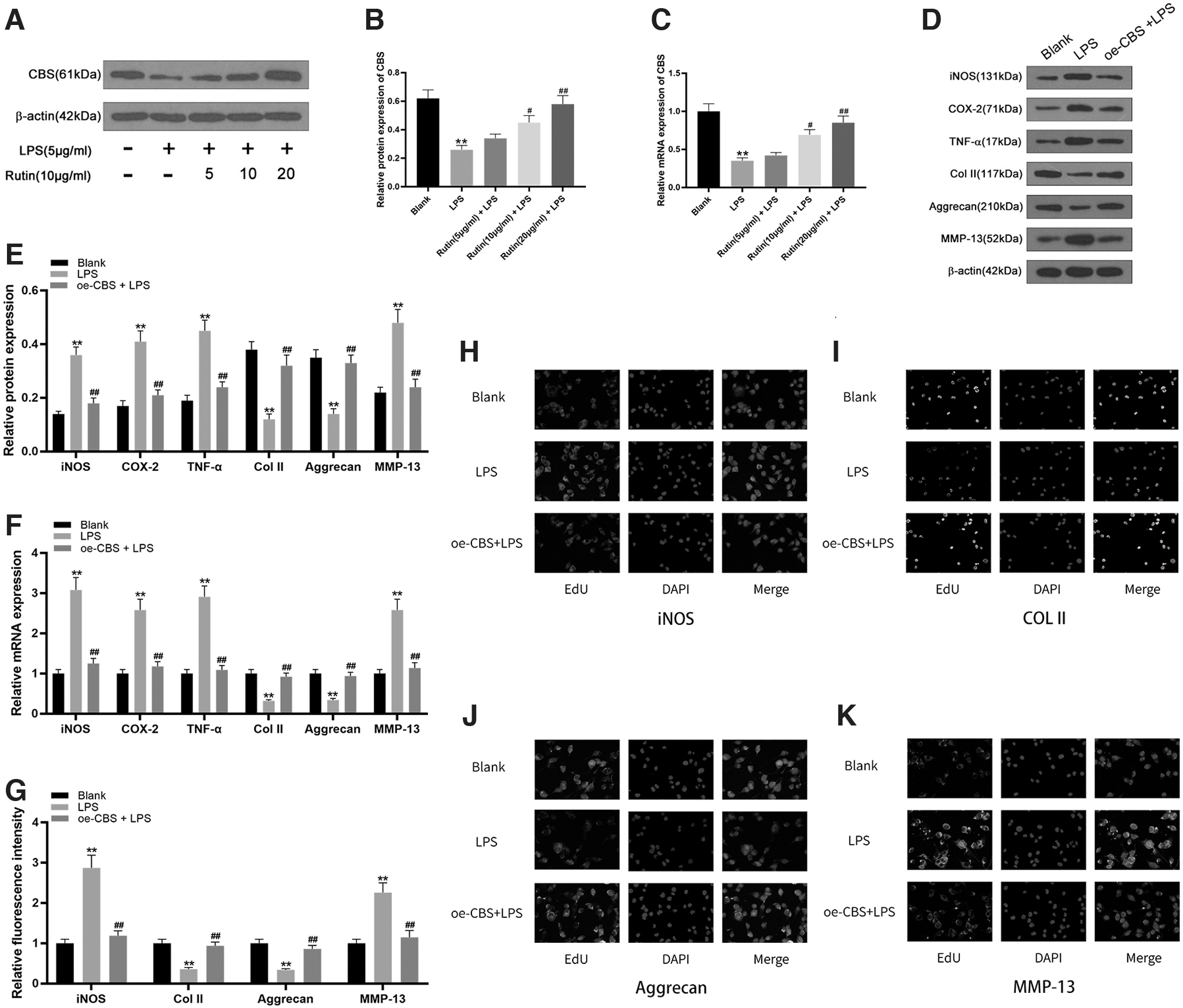
Rutin relieves the inflammatory response in LPS-treated chondrocytes by promoting expression of CBS.
After treating LPS-induced chondrocytes with CBS overexpression vector (oe-CBS), compared with the LPS-induced group, the protein and mRNA expression levels of iNOS, COX-2, TNF-α, and MMP-13 in the oe-CBS-induced group were significantly reduced, while the expression levels of Col II and Aggrecan were significantly increased (p < 0.001) (Fig. 4D–F). Immunofluorescence detection of iNOS, MMP-13, Col II, and Aggrecan protein expression in chondrocytes confirmed these findings (Fig. 4G–K). Therefore, it can be concluded that rutin alleviates the inflammatory response of LPS-induced chondrocytes by promoting the expression of CBS.
Rutin relieves the inflammatory response of LPS-induced chondrocytes by reducing the expression of ROCK
Compared with the control group, the protein and mRNA expression levels of ROCK1 and ROCK2 in chondrocytes of the LPS-induced group were significantly increased (p < 0.001). After treating LPS-induced chondrocytes with rutin at different concentrations of 5, 10, or 20 μg/mL, compared with the LPS-induced group, as the concentration of rutin increased, the protein and mRNA expression levels of ROCK1 and ROCK2 gradually but significantly decreased (p < 0.001) (Fig. 5A–C). Compared with the control group, the protein and mRNA expression levels of iNOS, COX-2, TNF-α, and MMP-13 significantly increased in the chondrocytes of the LPS-induced group, while the expression levels of Col II and Aggrecan significantly decreased (p < 0.001).
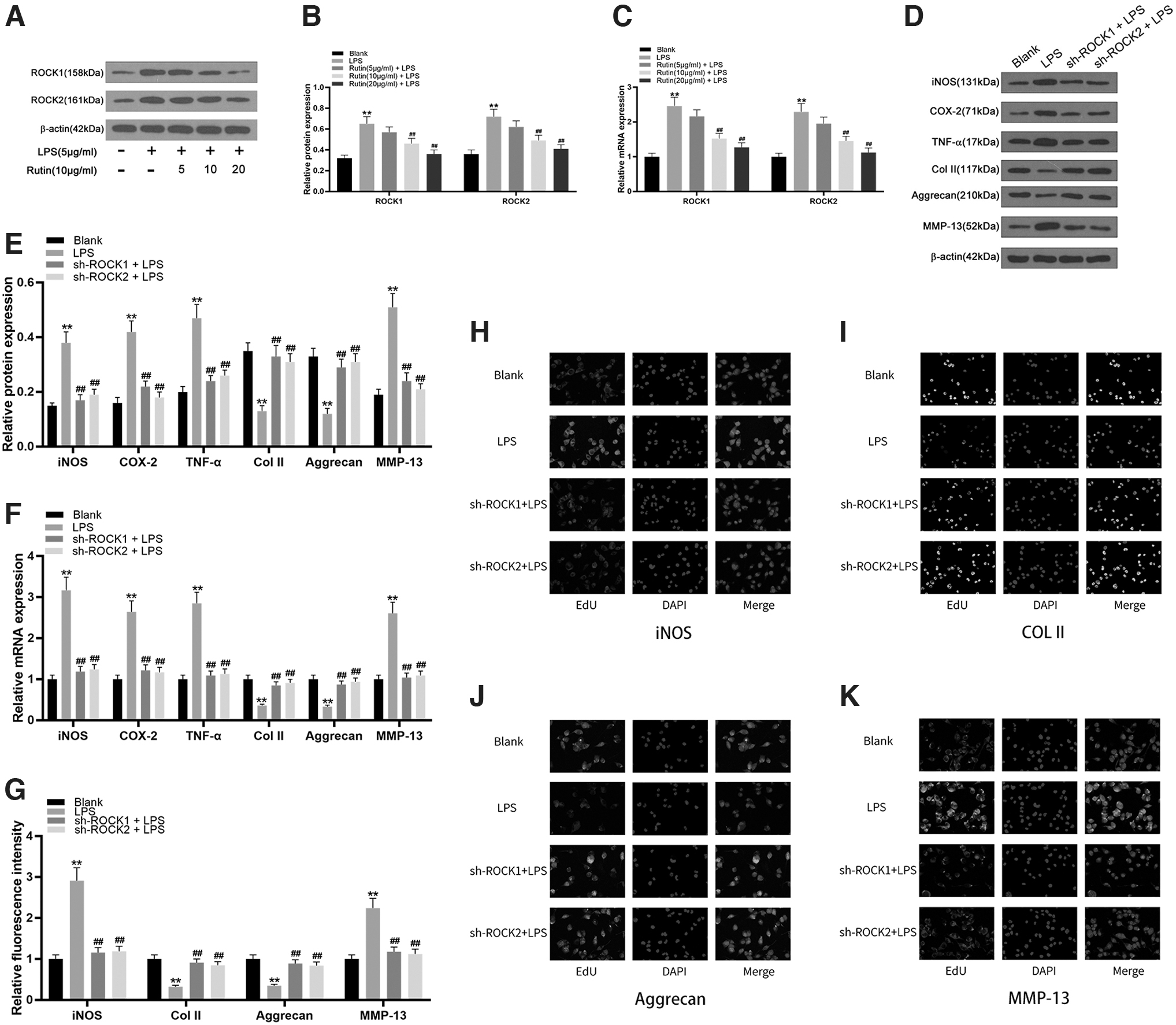
Rutin relieves the inflammatory response of LPS-induced chondrocytes by reducing the expression of ROCK.
After treating the LPS-induced chondrocytes with plasmid ROCK1 and ROCK2 inhibition vectors (sh-ROCK1, sh-ROCK2), compared with the LPS-induced group, the protein and mRNA expression levels of iNOS, COX-2, TNF-α, and MMP-13 in the sh-ROCK1- and sh-ROCK2-treated groups were significantly reduced, while the expression levels of Col II and Aggrecan rose significantly (Fig. 5D–F). Immunofluorescence was used to detect iNOS and mRNA in chondrocytes. The expression of MMP-13, Col II, and Aggrecan proteins were as described above for mRNA (Fig. 5G–K). Therefore, it can be concluded that rutin reduces the expression of ROCK to alleviate the inflammatory response of LPS-induced chondrocytes.
Rutin reduces expression of ROCK by promoting expression of CBS
Compared with the control group, the protein expression levels of ROCK1 and ROCK2 in chondrocytes of the LPS-induced group were significantly increased (p < 0.001), while the protein expression levels of ROCK1 and ROCK2 in the oe-CBS treatment group were significantly lower (p < 0.001) (Fig. 6A, B). The mRNA expression levels of ROCK1 and ROCK2 showed the same trends (Fig. 6C). Therefore, it can be concluded that rutin alleviates the inflammatory response of LPS-induced chondrocytes by promoting the expression of CBS and subsequently reducing the expression of ROCK.
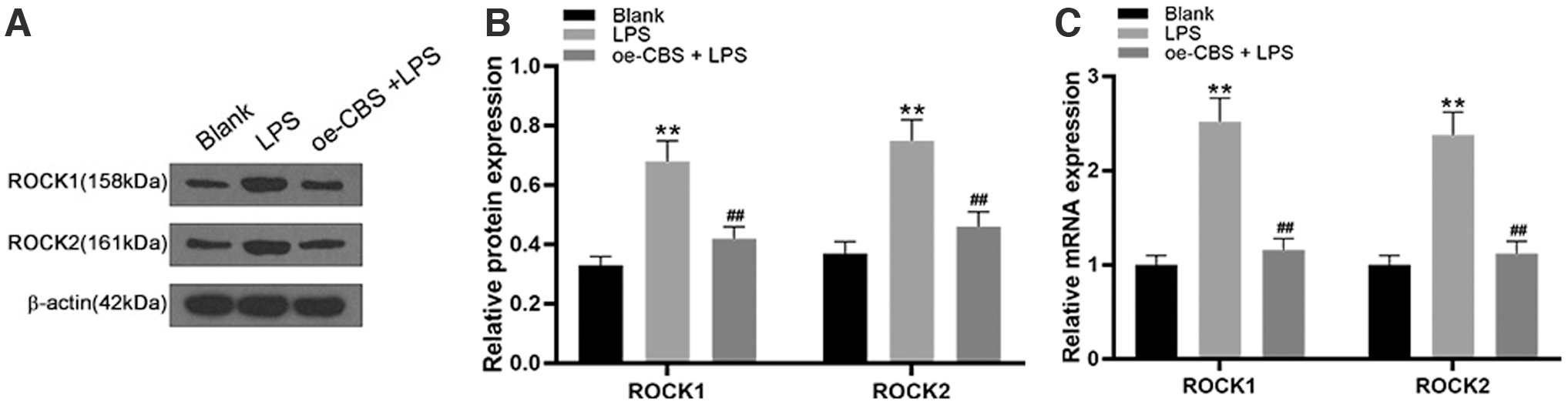
Rutin reduces the expression of ROCK by promoting the expression of CBS.
Rutin relieves OA inflammation in rats
To verify the effect of rutin on OA, rats in both the control group and OA group were injected with 0.9% saline in the knee joint cavity, while the rats in the OA+Rutin group received injections of 10 mg/mL rutin (Fig. 7A). There was no swelling in the knee joints of the rats in the control group, and their movements were normal; the knee joints of the rats in the OA group were obviously red and swollen, and their activities were obviously restricted. In contrast, although the knee joints of the rats in the rutin treatment group remained swollen, their degree of activity was significantly improved compared with the OA group (Fig. 7B).
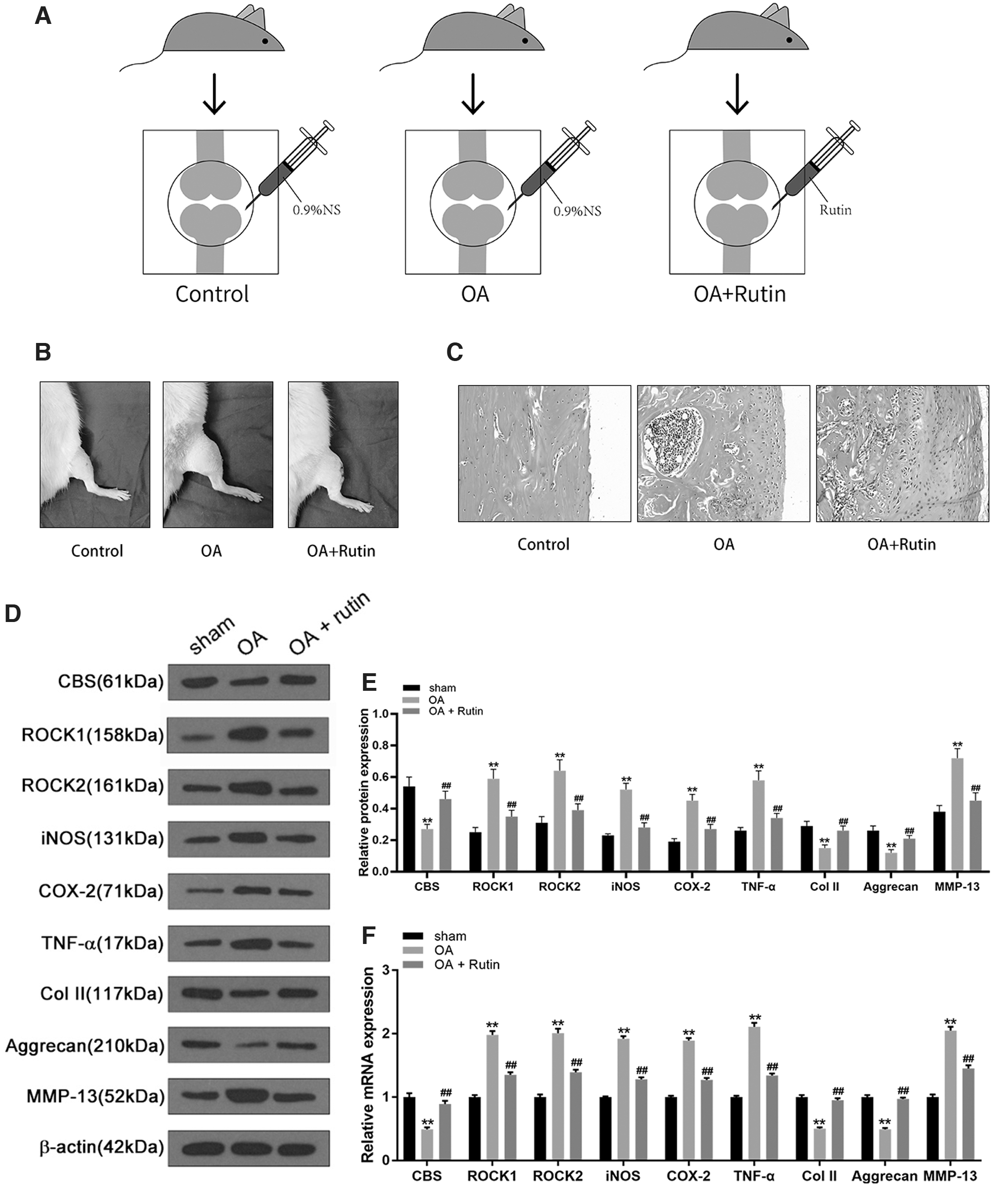
Rutin relieves OA inflammation following anterior cruciate ligament transection in rats.
HE staining of articular cartilage showed that the number and arrangement of chondrocytes in the control group were normal, without inflammatory cell infiltration; the number of chondrocytes in the OA group was significantly reduced, the cell arrangement was disordered, with a large number of infiltrating inflammatory cells; while the number of cartilage cells in the rutin treatment group was higher than that in the OA group, the cell arrangement was improved, and inflammatory cell infiltration decreased (Fig. 7C). Compared with the control group, the protein and mRNA expression levels of ROCK1, ROCK2, iNOS, COX-2, TNF-α, and MMP-13 in cartilage tissue of the OA group were all significantly increased, while the expression levels of CBS, Col II, and Aggrecan were significantly reduced (p < 0.001).
Compared with the OA group, the expression levels of ROCK1, ROCK2, iNOS, COX-2, TNF-α, and MMP-13 in the rutin treatment group were significantly reduced, while the expression levels of CBS, Col II, and Aggrecan significantly increased (p < 0.001) (Fig. 7D–F). These findings suggest that rutin effectively alleviates the inflammatory response of OA in rats and could play an important role in the treatment of OA.
Discussion
OA is considered to be a total joint disease characterized by the destruction and degradation of articular cartilage, subchondral bone edema and sclerosis, abnormal angiogenesis, and osteophyte formation (Yajun et al., 2021). Stabilization of articular cartilage and bone can ensure the normal weight-bearing of the joints. The destruction of this normal physiological relationship breaks the balance between cartilage cell anabolism and catabolism, resulting in cartilage damage and a reduction in joint space which ultimately leads to pain and limited joint mobility (Xia et al., 2014). Generally speaking, cartilage degeneration, abnormal subchondral bone metabolism, and synovial inflammation contribute to the pathophysiology of OA. These properties are also the main goals of many OA drugs and therapies (Li et al., 2021).
In our study, using the OA chondrocyte injury model and the rat OA model, we explored the effect of rutin in the treatment of OA and its potential mechanism through the CBS-mediated RhoA/ROCK signaling pathway. Our experimental study found that rutin effectively inhibits the inflammatory progression of OA by promoting the expression of CBS, and then negatively regulating the expression of the RhoA/ROCK signaling pathway.
In mammals, CBS is a key enzyme in the initiation and rate-limiting steps of the transsulfuration pathway, and is mainly involved in the production of endogenous H2S gas in the enzymatic pathway. In the cardiovascular system, the endogenous H2S catalyzed by CSE has the effects of regulating vascular tension and promoting angiogenesis and cardioprotection (Li et al., 2011). In the nervous system, CBS is mainly located in astrocytes and neuronal stem cells, and protects endothelial function through anti-inflammatory, antioxidant, and vascular generation effects (Wang et al., 2013). In this study, we found that CBS protein and mRNA levels were low in human OA samples, although the differential expression was significantly better than CSE and 3-MST, which indicated that the CBS activity in OA cartilage was higher than CSE and 3-MST.
In addition, release of IL-1β, TNF-α, nitric oxide (NO), prostaglandin E2, and other local inflammatory molecules affects the early progress of OA (Stefik et al., 2021). Studies have shown that inflammatory factors can also regulate the expression of CBS. Goren et al. (2019) found that among the proinflammatory cytokines, interferon-γ and IL-4 upregulated CBS gene transcription in normal human keratinocytes. However, Du et al. (2014) found that after exposure to the environmental toxin rotenone, mouse microglia will polarize to M1 to produce proinflammatory mediators (IL-1β, IL-6, TNF-α, iNOS, etc.), thus negatively regulating the expression of CBS.
Moreover, after we used rutin to interfere with OA chondrocytes, we found that the expression level of CBS was significantly increased, and effectively inhibited the expression of iNOS, COX-2, TNF-α, and MMP-13 inflammatory factors, as was observed in OA chondrocytes. The gene expression of CBS has also been found to effectively inhibit the expression of inflammatory factors, which shows that rutin effectively promotes the expression of CBS to inhibit the inflammatory progression of OA. However, a deficiency of this study is that we have not yet confirmed whether rutin can affect the content of endogenous H2S in OA chondrocytes, and further experimental studies are needed.
Ras homologous gene family member A (RhoA) is a small GTPase protein in the Rho family (Zhou and Zheng, 2013). ROCK is a downstream effector of RhoA, and has two homologous isomers: ROCK1 and ROCK2, which react to the prototype family member RhoA (Amano et al., 2010; Montagnoli et al., 2021). The RhoA/ROCK signal pathway is a very important signal transduction system, which is closely related to cell growth, differentiation, migration, and development (Loirand et al., 2006; Chen et al., 2013). This pathway is also required for neurite growth, bone formation, dorsal closure, and myogenesis (McBeath et al., 2004). Cytoskeletal remodeling is related to the pathogenesis of OA. RhoA/ROCK signaling participates in the early abnormal stimulation response of OA through modification of the chondrocyte actin cytoskeleton (Deng et al., 2019).
RhoA/ROCK promote the secretion of TGF-α, IL-1β, and leptin, and also induce the degradation of chondrocyte ECM (Wang et al., 2004; Pritchard et al., 2008; Long et al., 2013). Interestingly, TGF-α activates the RhoA/ROCK signaling pathway to induce degradation of articular cartilage (Appleton et al., 2010). Leptin also activates the RhoA/ROCK pathway and induces reorganization of the chondrocyte cytoskeleton, causing changes in cell shape (Liang et al., 2011). From this, we can conclude that in the early progress of OA, chondrocytes secrete inflammatory factors as a result of abnormal stimulation, which then activate the RhoA/ROCK signaling pathway. Two-way positive feedback regulation of inflammatory factors and the RhoA/ROCK signaling pathway accelerate the inflammatory progress of OA.
Based on these findings, we investigated whether inhibition of the RhoA/ROCK signaling pathway has an obstructive effect. Our experiments confirmed this, with the expression levels of iNOS, COX-2, TNF-α, and MMP-13 in the sh-ROCK1 and sh-ROCK2 treatment groups being significantly lower, while the expression levels of Col II and Aggrecan were significantly higher. In addition, the intervention of rutin effectively reduced the expression of ROCK1, ROCK2, and inflammatory factors. This shows that rutin effectively negatively regulates expression of the RhoA/ROCK signaling pathway to inhibit the inflammatory progression of OA.
In general, rutin has great pharmacological potential in inhibiting the inflammatory progression of OA. However, this study has certain limitations. We only found that rutin promotes the expression of CBS in OA chondrocytes, and then reduces the expression of ROCK, thereby inhibiting the inflammatory progression of OA. However, most studies have shown that CBS is the key enzyme for the enzymatic synthesis of endogenous H2S. H2S is an important endogenous gaseous signal transmitter in the human body, which participates in various physiological activities of apoptosis, angiogenesis, inflammation, metabolism, proliferation, and oxygen sensing (Zhang et al., 2021a, 2021b). In addition, the RhoA/ROCK signaling pathway can be activated by the endogenous H2S gas transmitter.
Zhang et al. (2021a, 2021b) confirmed that H2S catalyzed by the enzyme reaction inhibits the proliferation of reactive astrocytes after cerebral ischemia/reperfusion injury in mice by inhibiting the RhoA/ROCK pathway and promotes the recovery of nerve function. Zhang et al. (2020) confirmed that 3-MST/H2S protects brain endothelial cells from OGD/R-induced damage through mitotic protection and inhibition of the RhoA/ROCK pathway. Nalli et al. (2017) confirmed that CSE is selectively expressed in colonic smooth muscle cells, and that CSE-derived endogenous H2S inhibits muscle contraction through S-sulfur hydration of RhoA and inhibition of the RhoA/ROCK signaling pathway. Therefore, in the future, we will further study the role of CBS-derived endogenous H2S in inhibition of the progression of OA inflammation by rutin.
Conclusions
In summary, we found for the first time that rutin negatively regulates RhoA/ROCK signaling by promoting the expression of CBS, and effectively inhibits the inflammatory progress of OA. This study provides a theoretical basis for further research into the underlying mechanism of OA development and shows that rutin may be an effective candidate for therapeutic use in OA.
Ethics Statement
Clinical specimen experiments and animal experiments were reviewed and approved by the Ethics Committee of the First Affiliated Hospital of Anhui Medical University. The procedures used in this study adhere to the tenets of the Declaration of Helsinki.
Footnotes
Acknowledgments
Thanks to pharmacology laboratory teachers for their technical support and assistance.
Authors' Contributions
C.S., Y.W., and J.W. designed this study; Y.W. and T.Z. completed OA specimen experiments; C.S., Y.W., and R.Z. performed cell culture studies; Y.Z., J.X., and Y.D. performed in vivo studies; R.Z. and Y.Z. collected data; C.S. and Y.W. analyzed data; J.W. and Y.H. revised the article.
Disclosure Statement
No competing financial interests exist.
Funding Information
This study was supported by the Open Fund Project of the Key Laboratory of Anti-inflammatory and Immunological Drugs of the Ministry of Education (KFJJ-2020-04); Major Project of Natural Science Foundation of Anhui Province Colleges and Universities (KJ2020ZD17); 2020 Natural Science Foundation of Anhui Medical University (2020xkj169).