
Abstract
Skeletal muscle mass is closely related to strength and health. Multiple genes and signaling pathways are involved in the regulation of skeletal muscle hypertrophy. miR-29 can participate in various processes of skeletal muscle development through different target genes. However, studies are needed on the function of miR-29 in skeletal muscle during mouse puberty. We used mice in which overexpression of miR-29ab1 cluster could be induced specifically within skeletal muscle, and investigated the effects of miR-29 overexpression on skeletal muscle at 1 month of age. We found that the overexpression of miR-29ab1 cluster in juvenile mice caused skeletal muscle mass and myofiber cross-sectional area to increase. The study on the mechanism of miR-29 inducing skeletal muscle hypertrophy had found that miR-29 achieved its function by inhibiting the expression of Mstn. At the same time, injured myofibers were present within miR-29ab1 cluster overexpressing skeletal muscle. The damage of skeletal muscle may be due to the inhibition of the type IV collagen by miR-29. These results indicate that although the overexpression of miR-29ab1 cluster can induce skeletal muscle hypertrophy in mouse juvenile, it simultaneously causes skeletal muscle damage.

Introduction
Skeletal muscle has tremendous plasticity partially due to the dynamic balance between protein synthesis and degradation. Increased protein synthesis leads to muscle hypertrophy, whereas increased protein degradation leads to muscle atrophy. Sufficient muscle mass and strength are important for performance of exercise, but these characteristics are also associated with health and longevity (Schoenfeld, 2010; Wackerhage et al, 2019). However, it is not easy to make muscle hypertrophy; hypertrophy often depends on resistance training and proper nutrition (Celis-Morales et al, 2018).
Expression changes of certain genes can affect skeletal muscle mass, such as knockdown of MSTN causes hypertrophy, whereas overexpression of IGF1 induces hypertrophy. MSTN is involved in the regulation of skeletal muscle by binding to the activin receptor IIB, recruiting ALK4/ALK5, causing the phosphorylation of Smad2/3, and allowing the Smad2/3/4 complex to enter the nucleus, thereby causing skeletal muscle atrophy (Han et al, 2013). IGF1 induces skeletal muscle hypertrophy, which is related to the activation of Akt. The activation of Akt can inhibit skeletal muscle degradation and promote skeletal muscle hypertrophy (Bodine et al, 2001b; Pallafacchina et al, 2002).
MiRNAs play important roles in the process of skeletal muscle hypertrophy, such as skeletal muscle star miRNAs, miR-1 and miR-133. miR-1 and miR-133 can reduce the phosphorylation level of Akt by inhibiting Igf-1, Igf-1R, and HSP70 and affect skeletal muscle hypertrophy (Elia et al, 2009; Huang et al, 2011). Muscle-specific miR-128a can inhibit the expression of IRS1 and reduce the activity of Akt, and thereby, the inhibition of miR-128a can cause skeletal muscle hypertrophy (Motohashi et al, 2013). miR-486 can activate Akt signaling pathway by inhibiting PTEN, so overexpression of miR-486 can resist skeletal muscle atrophy, while inhibition of miR-486 can cause muscle fiber atrophy (Hitachi et al, 2014; Xu et al, 2012).
The miR-29 family comprises miR-29a, miR-29b, and miR-29c. Although miR-29 is not a skeletal muscle-specific miRNA, miR-29 family is closely related to the development of skeletal muscle in mice after birth. The expression of miR-29 gradually increases with the development of skeletal muscle (Liu et al, 2019). In neonatal mice, the relative amount of miR-29 is minimal, and the ectopic expression of miR-29ab1 cluster inhibits the expression of type IV collagen and causes skeletal muscle dysplasia (Liu et al, 2019). In adult mice, the expression of miR-29 was at a high level, and knockout of miR-29 would cause skeletal muscle atrophy. Correspondingly, overexpression of miR-29ab1 cluster could resist skeletal muscle atrophy (Liu et al, 2021; Liu et al, 2019). The increased expression of miR-29 in elderly mice has been suggested to be a possible inducer of skeletal muscle senescence (Hu et al, 2014).
Well, there are few studies on the function of miR-29 in mouse puberty, so our study tested the effect of overexpression of miR-29ab1 cluster in puberty mice. We found in 1-month-old mice, miR-29ab1 cluster mainly induced muscle hypertrophy by repressing Mstn expression. However, similar to the results of inducing miR-29ab1 cluster overexpression after birth, inducing miR-29ab1 cluster overexpression during puberty could cause skeletal muscle damage. These results indicate that the effect of miR-29 on skeletal muscle is multifaceted and dynamic.
Materials and Methods
Mice
This study was approved by the Institutional Animal Care and Use Committee of China Agricultural University (SKLAB-2015-01-03).
ACTA1-rtTA mice and TRE-miR-29 mice were previously reported (Liu et al, 2019) and generated based on the Tet-on system. In brief, the ACTA1-rtTA vector was generated by replacing the original PCMV promoter with the Acta1 promoter in the pTet-on Advanced vector. TRE-miR-29 vectors were constructed to clone the sequence of miR-29ab1 cluster into the multiple cloning site of the pTRE-Tight vector. The corresponding transgenic mice were obtained by pronuclear microinjection (Beijing Jifulin Biotech).
Doxycycline (2 g/L) dissolved in 5% sucrose supplied as drinking water, which was exchanged every 3 days. Doxycycline is a nontoxic compound widely used in medicine (Urlinger et al, 2000).
Mice were raised at an appropriate temperature (25°C ± 1°C) as well as humidity (60–70%) with ad libitum access to food as well as water, 12-h light/12-h dark cycle. Mice were sacrificed by cervical dislocation and muscles, hearts, and white adipose tissue of epididymis were isolated, weighed, and either stored at −80°C or fixed in 4% PFA.
All mice used in the experiments were male. Doxycycline (Dox) was added to drinking water from 1 month of age to induce miR-29ab1 cluster overexpression. A total of 4 wild-type mice (C57BL/6), 21 control, and 21 dTG mice were used.
Among them, three control and three dTG mice were used to detect the expressions of related genes 3 days after induction; three control and three dTG mice were used to detect the expressions of Mstn 5 days after induction; three control and three dTG mice were used to detect glucose tolerance; five control and five dTG mice were used to detect the relative weight of muscle and fat as well as the expression of Mstn 28 days after induction; three control and three dTG mice were used to detect the relative weight of heart 28 days after induction; and four dTG mice failed to achieve overexpression of miR-29ab1 cluster, and the corresponding four control mice did not appear in the final experimental results. Two wild-type mice were used for control agomir injections and two wild-type mice were used for miR-29b agomir injections.
Agomir injections
One-month-old wild-type mice were continuously injected with 5 nmol agomir (control or miR-29b) (RiboBio) in the left tibialis anterior muscle on days 0, 2, and 4, and then the samples were taken at 6 days for follow-up detection.
Glucose tolerance test
After 28 days of Dox feeding, the glucose tolerance test (GTT) was performed. The day before the GTT, mice fasted overnight and drank freely. Each mouse was first intraperitoneally injected with 100 μL glucose (2 g/kg). Blood samples were collected from the tail at 0, 30, 60, 90, and 120 min after injection to measure the blood glucose concentration of each mouse. Blood samples collected at 0, 30, and 120 min were required for serum separation; serum was used to detect insulin concentration (80-INSMS-E01; Alpco).
Analysis of real-time PCR
The total RNA of tissues was isolated using Total RNApure Reagent (Beijing Zoman Biotechnology Co., Ltd.). RNA was reverse transcribed to cDNA using M-MLV Reverse Transcriptase (Promega). The RT primer sequences for miR-29a and miR-29b are consistent with the previous article (Liu et al, 2019).
For analysis of miRNA expression, the sequences of the primers are consistent with the previous article (Liu et al, 2019).
For analysis of mRNA expression, the sequences of the main primers are as follows:
Col4a1, 5′-TGGGAAAGACGGTGAAAAAG-3′ and 5′-AAAGGCATGGTGCCTATCAC-3′;
Col4a2, 5′-CATCCGTCGGAGATGAAGAT-3′ and 5′-CAAACAGGAAGCCATCTGGT-3′;
Mstn, 5′-CTGTAACCTTCCCAGGACCA-3′ and 5′-GCAGTCAAGCCCAAAGTCTC-3′;
CTSL, 5′-GTGGACTGTTCTCACGCTCA-3′ and 5′-TGTCATTAGCCACAGCGAAC-3′;
Atrogin-1, 5′-AGCGCTTCTTGGATGAGAAA-3′ and 5′-ACGTCGTAGTTCAGGCTGCT-3′;
MuRF1, 5′-TGGAAACGCTATGGAGAACC-3′ and 5′-AACGACCTCCAGACATGGAC-3′; and
GAPDH, 5′-GGCTGCCCAGAACATCAT-3′ and 5′-CGGACACATTGGGGGTAG-3′. GAPDH was used as the internal standard.
Western immunoblot analysis
The experimental procedure of Western immunoblot was as described in previous studies (Liu et al, 2017). Antibodies used were anti-GAPDH (2118S; Cell Signaling Technology) and anti-p53 (2524S; Cell Signaling Technology).
Histology and immunofluorescence
The isolated skeletal muscles were fixed in 4% PFA. Then paraffin embedding and sectioning were performed. Sections were deparaffinized and hydrated, as well as subjected to H&E staining or immunofluorescence. For immunofluorescence, the antigen retrieval was accomplished in high barometric pressure, blocked using 5% goat serum, and primary antibodies used included anti-eMyHC (Developmental Studies Hybridoma Bank) and anti-Collagen IV (ab6586; Abcam).
Cell culture
HEK293T cells (Cell Resource Center, Beijing, China) were cultured in DMEM containing 10% fetal bovine serum and 1% streptomycin and penicillin at 37°C with 5% CO2; 0.5 mL of HEK293T cells with a concentration of 0.8 × 105/mL were seed to the 24-well plate before transfection by the Lipo6000™ (Beyotime Biotechnology).
Luciferase reporter assay
The Mstn 3'UTR was amplified from mouse (C57BL/6) cDNA (including the putative miR-29 binding sites), and this fragment was cloned into the 3′UTR of Renilla luciferase in the psiCHECK™-2 vector. The mutant Mstn 3′UTR (the putative binding sites for miR-29 were mutated) was generated by fusion PCR. The vector and mimics (or control) were co-transfected into HEK293T cells and finally cultured for 24 h.
Luciferase activity was measured using the Dual Luciferase Assay System (Promega) and the Synergy HT Multi-Detection Reader (BioTek Instruments). Firefly luciferase activity served as an internal reference.
Statistical analysis
Venn diagram is generated by jvenn (Bardou et al, 2014). Statistical analysis was performed using SPSS 16.0. The experimental results were presented as mean ± SEM, and analyzed by Student's t-test. * means statistically significant (p < 0.05) and ** means statistically significant (p < 0.01). The cross-sectional area (CSA) was analyzed using Image J.
Results
The transgenic mice (dTG mice) (Liu et al, 2019) used in this study were generated by the Tet-On System, so the expression level of miR-29ab1 cluster can be regulated by adjusting Dox amount. The expression of type IV collagen was significantly lower in the wild-type 1-month-old mice muscle than in the newborn mice muscle, and overexpression of miR-29 may not cause skeletal muscle dysplasia as overexpression of miR-29 after birth (Liu et al, 2019).
Therefore, to explore the influence of miR-29 overexpression on skeletal muscle development at puberty, we began adding Dox to drinking water when the mice were 1-month old (Fig. 1A). Interestingly, the sustained induction of the miR-29ab1 cluster for 28 days in the 1-month-old mice resulted in significantly increased muscle mass in dTG mice (Fig. 1B–D). In skeletal muscle hypertrophy mice, the ratio of fat is usually changed (McPherron et al, 1997). We also found the relative weight of the white adipose tissue of epididymis (eWAT) in dTG mice was reduced (Fig. 1B–D).
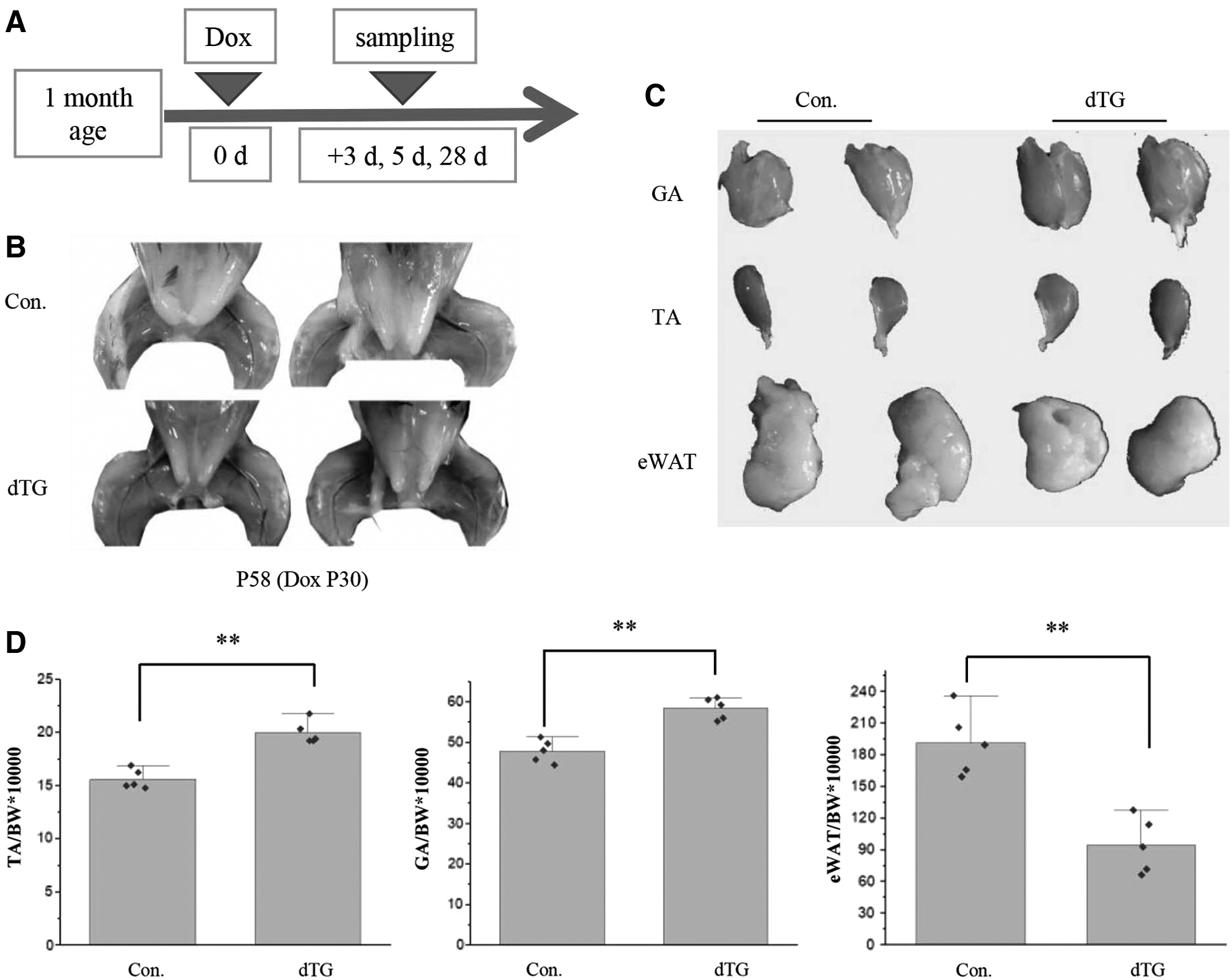
miR-29ab1 cluster overexpression induces muscle hypertrophy in 1-month-old mice. TA, tibialis anterior muscles; GA, gastrocnemius muscles; eWAT, white adipose tissue of epididymis; BW, body weight; Con., the littermate control mice; dTG mice, the miR-29ab1 cluster overexpression mice. P30, 30 days of postnatal development.
Skeletal muscle hypertrophy is often accompanied by cardiac hypertrophy, which is often very dangerous for individuals. So we examined whether skeletal muscle-specific overexpression of miR-29 has an effect on the heart. We examined whether there would be the ectopic expression of miR-29 in heart of dTG mice. Q-PCR analysis revealed that there was no difference in the expression of miR-29b in the hearts of control and dTG mice (Fig. 2A), and the relative weight of the hearts was not affected (Fig. 2B, C).
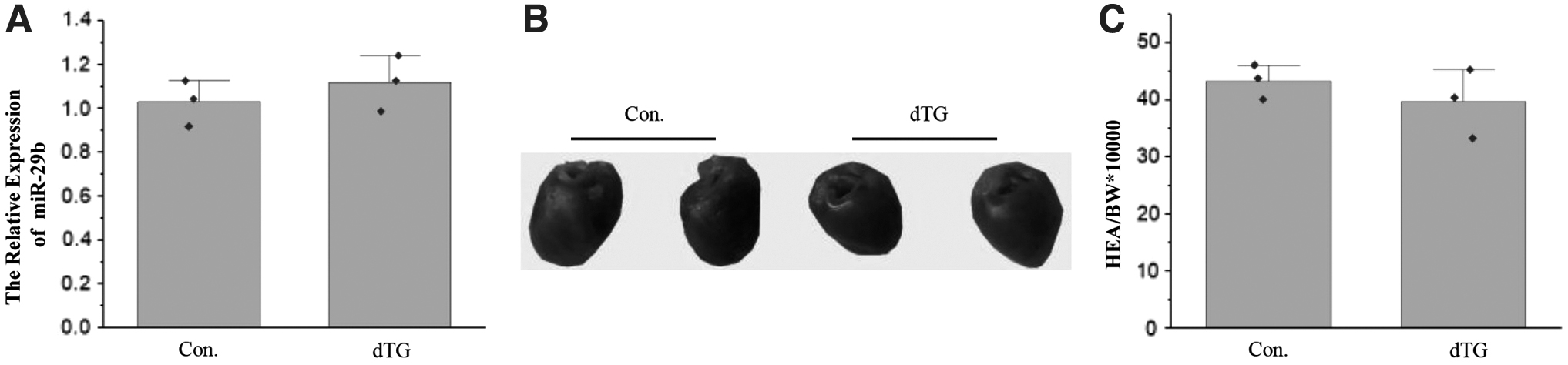
The relative weight of the hearts was not affected.
Changes in the proportion of muscle and fat weight may affect the sensitivity of blood glucose and insulin levels. According to an intraperitoneal blood glucose test, the fasting blood glucose level was lower in dTG mice than in the control mice (Fig. 3A), and dTG mice regulated blood glucose at lower insulin levels (Fig. 3A, B).
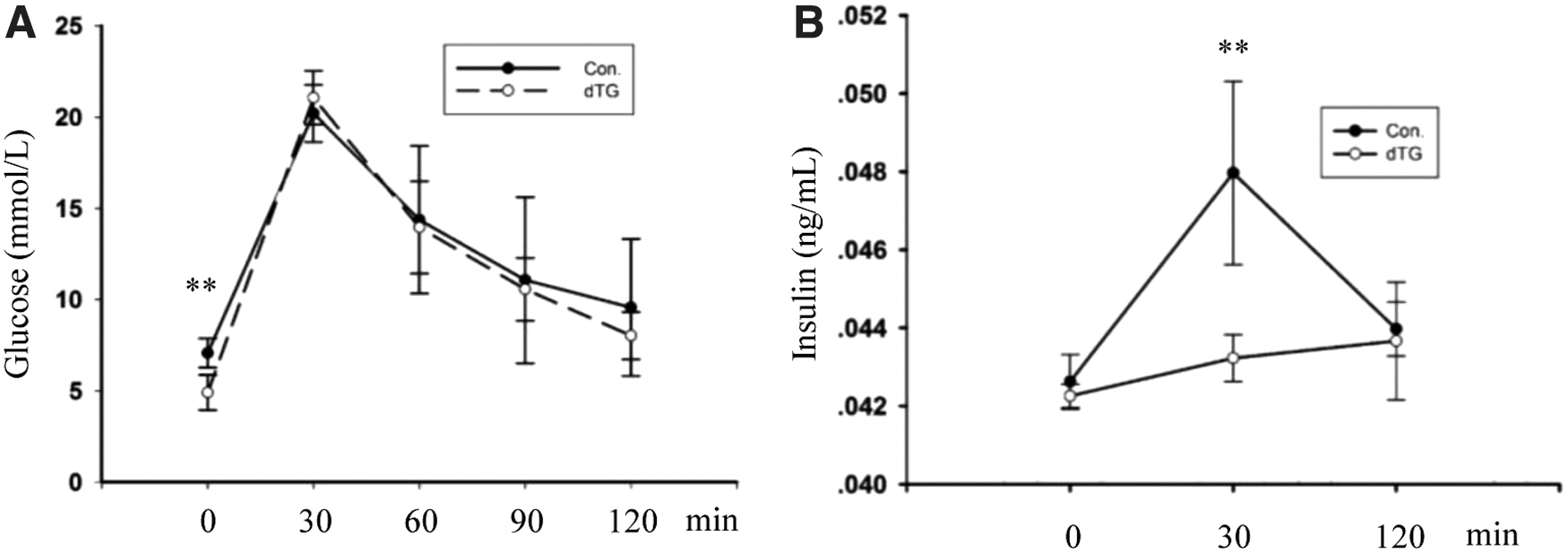
Changes of blood glucose in dTG mice. Results of glucose test in dTG mice and control mice. The mice were treated with Dox at P30. Fasting was at P57, assays were performed at P58. The values represent the mean ± SEM (n = 3). **p < 0.01.
Consistent with the increase in muscle mass, H&E staining revealed that the muscle CSA in dTG mice was larger than that in the controls (Fig. 4A). According to the quantification of the fiber size, the CSA of the TA muscle fibers was also significantly increased in dTG mice (Fig. 4B).
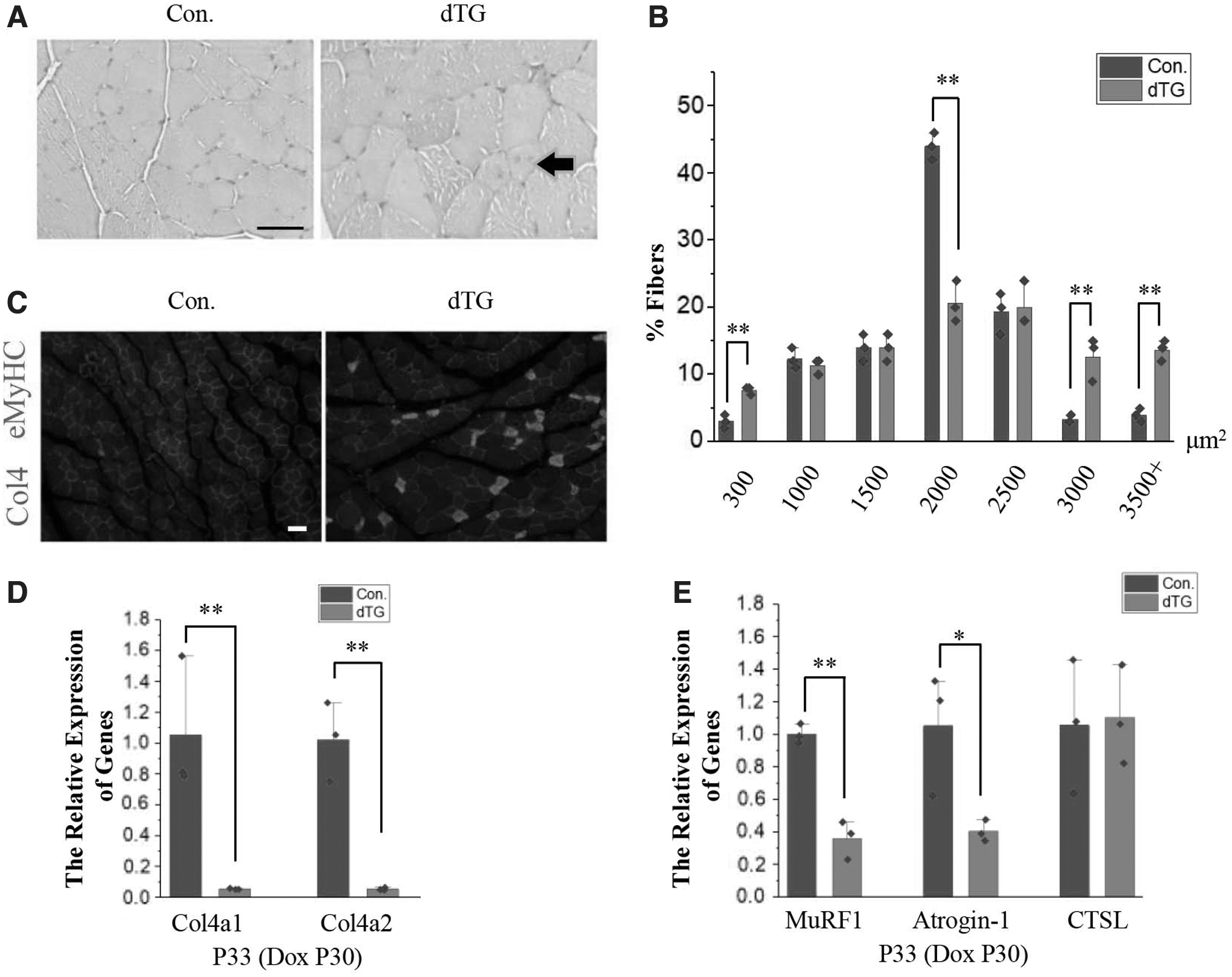
Hypertrophy coexists with damage in the skeletal muscles of dTG mice.
At the same time, we found several muscle fibers contained centrally located nuclei (black arrows) of dTG mice (Fig. 4A). The eMyHC-positive myofibers were observed in dTG mice (Fig. 4C). Postnatal overexpression of miR-29 can lead to skeletal muscle dysplasia by repressing Col4a1 and Col4a2 (Liu et al, 2019). And the expression of Col4a1 and Col4a2 was also significantly decreased in the Dox-treated dTG mice during puberty (Fig. 4D). Inhibition of the expression of type IV collagen may be the cause of skeletal muscle damage, but it is not the cause of skeletal muscle hypertrophy.
Meanwhile, miR-29 was reported to resist skeletal muscle atrophy, and MuRF1 is a target gene of miR-29 (Liu et al, 2021). The downregulation of MuRF1 and Atrogin-1 can prevent muscle atrophy, but does not result in skeletal muscle hypertrophy (Bodine et al, 2001a; Wang and Pessin, 2013). So while the expression of MuRF1 and Atrogin-1 was inhibited (Fig. 4E), this may not be the reason for skeletal muscle hypertrophy.
We also verified the regulatory effect of miR-29 on MuRF1 and Atrogin-1 by injecting agomir of miR-29b into the tibialis anterior muscle of wild-type mice. The results of 6 days after the first injection showed that miR-29b agomir could inhibit the expression of MuRF1 and Atrogin-1 (Supplementary Fig. S1A, B). The transcription factor p53 is also closely related to skeletal muscle metabolism, and its expression is increased during skeletal muscle atrophy. Upregulated expression of p53 can cause atrophy, and inhibition of its expression can resist skeletal muscle atrophy (Fox et al, 2014; Langer et al, 2022; Schwarzkopf et al, 2006). Therefore, we also detected the expression changes of p53 after agomir injection, and the results showed that overexpression of miR-29b inhibited the expression of p53 (Supplementary Fig. S1C).
To analyze the regulatory mechanism of miR-29, we used targetscan7.1 (
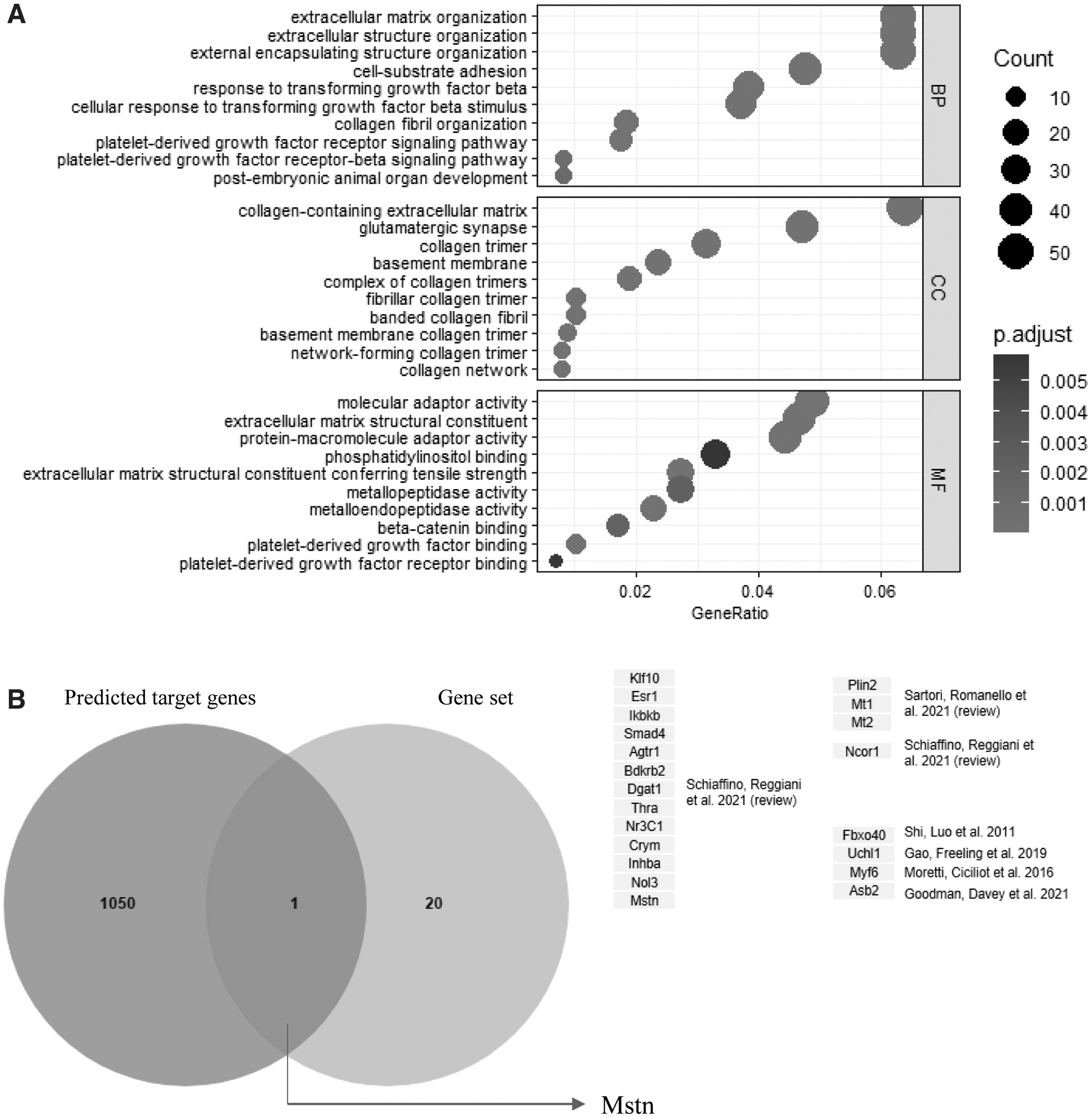
miR-29 target gene analysis.
The sequence complementary to the seed sequence of miR-29 in the 3'UTR of the Mstn gene was conserved in multiple species (Fig. 6A). In our study, according to the Q-PCR analysis, the miR-29ab1 cluster and miR-29b agomir decreased the expression level of Mstn (Fig. 6B and Supplementary Fig. S2). To further verify whether miR-29 could directly act on the complementary sequences of the 3'UTR of the Mstn gene, the predicted binding site of miR-29 in the 3'UTR of the Mstn gene was cloned into the 3'UTR of a Renilla luciferase in the psiCHECK-2 vector (Fig. 6C). Also, the predicted miR-29 binding site in the 3'UTR of the Mstn gene was mutated and cloned into the same position of the vector (Fig. 6C).
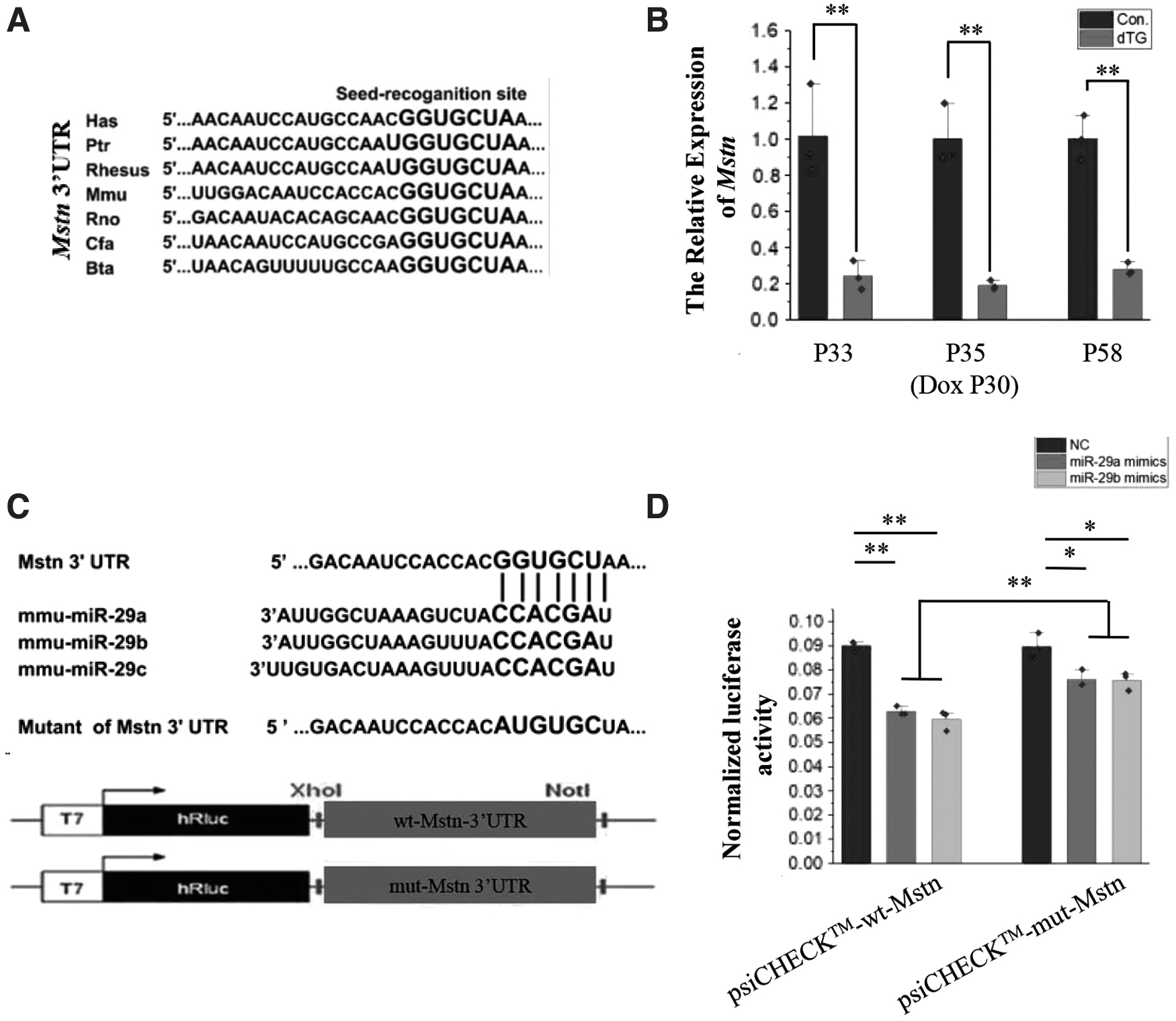
Mstn is a direct target gene of miR-29a, b.
Compared with cells co-transfected with NC and psiCHECK-wt-Mstn vector (psiCHECK-2 vector containing wild-type Mstn 3'UTR), the relative activity of luciferase in cells co-transfected with miR-29a or miR-29b mimics and psiCHECK-wt-Mstn vectors decreased significantly (Fig. 6D). The relative activity of luciferase in the cells co-transfected with miR-29a or miR-29b mimics and psiCHECK-mut-Mstn vector (psiCHECK-2 vector containing mutant Mstn 3'UTR) was still different from that in the cells co-transfected with NC and psiCHECK-mut-Mstn vector, but the range of change was reduced (Fig. 6D). Therefore, miR-29 may regulate the expression of Mstn through this complementary site.
Discussion
In this study, we found that overexpression of miR-29ab1 cluster could cause skeletal muscle hypertrophy at puberty by inhibiting Mstn. MSTN is important for skeletal muscle metabolism and is considered a master regulator of skeletal muscle growth. MSTN expression has been reported to be increased in several human diseases associated with skeletal muscle wasting, such as cancer cachexia, AIDS, and heart failure (Han et al, 2013). Moreover, its overexpression induces muscle atrophy (Zimmers et al, 2002). Studies have shown that miR-29 can resist muscle atrophy by repressing MuRF1 (Liu et al, 2021), and our results showed that Mstn was regulated by miR-29. Therefore, the regulation of skeletal muscle atrophy by miR-29 should include the co-suppression of Mstn and MuRF1. This shows that miR-29 is very important as a therapeutic factor for skeletal muscle atrophy.
Although miR-29 could inhibit Mstn, induction of skeletal muscle hypertrophy by miR-29 is not feasible, because miR-29 has an important inhibitory effect on the extracellular matrix-related genes, and skeletal muscle hypertrophy requires upregulated expression of the extracellular matrix-related genes. Therefore, under certain conditions, hypertrophy induced by miR-29 can lead to skeletal muscle damage.
Because of the important inhibitory effect of miR-29 on extracellular matrix-related genes, miR-29 is considered to be an important therapeutic factor for fibrosis in various tissues and organs. Studies have shown that miR-29 can treat cardiac fibrosis (van Rooij et al, 2008). In view of the effect of miR-29 on Mstn, the possible side effects of miR-29 for cardiac therapy need to be reconsidered.
Based on previous research results (Liu et al, 2021; Liu et al, 2019), as well as our present findings, we could establish the physiological role of miR-29 in mice after birth. In neonatal mice, miR-29 is at a low expression level, and such an expression level is beneficial for the establishment of the extracellular matrix of skeletal muscle. During the development of skeletal muscle in mice, the expression of miR-29 gradually increases, the expression of extracellular matrix-related proteins declines, and the upregulated expression of miR-29 is to contribute to the development of skeletal muscle. Therefore, the overexpression or knockout of miR-29 after birth will cause severe skeletal muscle dysplasia.
Insulin resistance is an important metabolic disease, and studies in experimental animals show that insulin resistance can be resisted by regulating Mstn (Tang et al, 2014; Wilkes et al, 2009). Therefore, the results of this article on the regulation of Mstn by miR-29 may provide new ideas for the prevention of insulin resistance. Insulin resistance mostly occurs in middle-aged and elderly people (Shou et al, 2020), so this study will follow up to verify the role of miR-29 in resisting insulin resistance in adult and elderly mice.
Conclusion
In conclusion, it is not feasible to induce skeletal muscle hypertrophy by overexpression of miR-29 in either animal husbandry or human fitness. While miR-29 may be an important target against adult skeletal muscle atrophy, the effect of miR-29 is related to the inhibition of Mstn and MuRF1.
Footnotes
Acknowledgment
We would like to appreciate Yunping Dai for his kind help in animal model preparing.
Data Availability Statement
The data and material are available from the corresponding author on reasonable request.
Disclosure Statement
The authors declare no conflict of interests.
Funding Information
This research was supported by National Key Research and Development Program of China, grant number 2021YFF1000603; the National Natural Science Foundation of China, grant number 31790412 and 31970712; the China Agriculture Research System of MOF and MARA, ePlan 111, grant number B12008; the National Research Facility for Phenotypic and Genotypic Analysis of Model Animals (Beijing); the Project for Extramural Scientists of the State Key Laboratory of Agrobio Technology, grant number 2020SKLAB6–9; the Innovation Foundation of Inner Mongolia University of Science & Technology, grant number 2019QDL-B43; and Research Program of Science and Technology at Universities of Inner Mongolia Autonomous Region of China, grant number NJZY20096.
Supplementary Material
Supplementary Figure S1
Supplementary Figure S2
References
Supplementary Material
Please find the following supplemental material available below.
For Open Access articles published under a Creative Commons License, all supplemental material carries the same license as the article it is associated with.
For non-Open Access articles published, all supplemental material carries a non-exclusive license, and permission requests for re-use of supplemental material or any part of supplemental material shall be sent directly to the copyright owner as specified in the copyright notice associated with the article.