
Abstract
Varicoceles (VCs) have received widespread attention as a primary factor affecting male fertility and a pathological condition that may lead to decreased sperm count and motility in patients. Many studies have shown that an imbalance of local antioxidant balance exists in patients with VC, leading to an obvious increase in the content of reactive oxygen species (ROS) and may cause reductive stress. Excessive ROS may aggravate spermatogenesis dysfunction and affect male fertility. Poly(ADP-ribose) polymerase (PARP) is an enzyme associated with DNA repair in eukaryotic cells, can be activated by DNA fragments with structural damage, and has been considered a DNA damage receptor in DNA damage repair and apoptosis. We built a rat model of VC and an oxidative damage model of a spermatocyte-derived cell line (GC-2 cells) induced by hydrogen peroxide to study the role of PARP1 in VC. Differentially expressed genes (DEGs) were obtained by RNA sequencing in the testes of VC rats. Analysis of DEGs revealed some genes with significantly altered expression, which were validated in rat and cell models. Immunofluorescence, real-time quantitative PCR analysis, Western blot, and flow cytometry were used to analyze the changes between the control group and the VC or hydrogen peroxide group. Overall, we found that PARP1 protein expression increased in VC rats and in the hydrogen peroxide-induced oxidative stress model of GC-2 cells. Parthanatos may be one of the factors leading to reduced reproductive capacity in VC patients. Our study provides novel insights into the mechanisms of male infertility induced by oxidative stress and provides a new therapeutic target for VC.
Introduction
According to the World Health Organization, infertility will become the third biggest disease in the world in the 21st century after tumor and cardiovascular and cerebrovascular diseases (2014). Varicoceles (VCs) are one of the most important factors affecting male reproductive, directly or indirectly affecting 60% of infertility patients (He et al, 2021). VC can cause abnormal varicose veins, which affect the testicle's local blood circulation, resulting in coniferous tubule damage, germ cell loss, testicular atrophy, a reduction in numbers of Leydig cells, and a decline in testosterone levels. This causes parthenogenesis disorder and has been associated with an increase in cell apoptosis, resulting in testicular sperm production function obstacles (Agarwal et al, 2012; Hamada et al, 2013).
The psychophysiology mechanisms of male infertility are as follows: (1) testis hypoxia, (2) heat stress, and (3) spermatozoon cell apoptosis. Pathological changes may lead to an increase in local oxidative stress, resulting in a significant increase in local reactive oxygen species (ROS) content and leading to further damage to tissues and cells. Overproduction of ROS may lead to changes in several molecular biological pathways (Wang et al, 2022a).
Low levels of ROS are required for parthenogenesis, sperm activation, and chromosomal reactions. White blood cells are the main source of ROS in the male reproductive system (Barati et al, 2020; Ranawat and Bansal, 2009). The testis produces excessive ROS in inflammatory and ischemia environments, and the ROS content in the infertile male reproductive system is obviously increased. In leukocytospermia and VC, white blood cells rise in the semen, damaging the sperm and leading to miscarriage (Mongioì et al, 2020). Excess ROS may lead to inflammation, which may further induce ROS production (Baazm et al, 2020). The influence of heavy metals on drinking water or food, and nanopolymers through the blood–testosterone barrier can also lead to the generation and accumulation of excessive ROS in testicular tissue (Kozopas et al, 2020; Marmar, 2001).
Many researchers have found that ROS induced by oxidative stress may play a crucial role in male infertility. In VC, the level of ROS in the testis is obviously increased, and the activity and expression of superoxide dismutase (SOD) and other antioxidant enzymes are significantly decreased (Oh et al, 2016).
Many studies have shown that VC can lead to testicular cell damage and apoptosis, and cause spermatogenic cell apoptosis and sperm apoptosis, and some researchers have found the accumulation of iron ions in testis of VC patients and speculated that ferroptosis may be one of the pathogenic factors of VC (Sanei-Ataabadi et al, 2020; Shaygannia et al, 2021). Parthanatos, as a newly discovered noncaspase-dependent programmed cell death, is characterized by poly(ADP-ribose) polymerase-1 (PARP1) dependence (Wang and Ge, 2020). PARP1, a 113-kDa protein, accounts for more than 90% of intracellular PARP activity (Alemasova and Lavrik, 2019) and is a DNA repair enzyme that mainly exists in the nucleus of eukaryotes. Under normal physiological conditions, PARP1 repairs DNA damage.
In the pathological state, a large amount of DNA damage occurred, and the overactivation of PARP1 catalyzed the decomposition of intracellular NAD into nicotinamide and poly-ADP ribose (PAR) accumulation in the nucleus (Gibson and Kraus, 2012). The accumulation of PAR and the consumption of NAD inhibited the activity of the mitochondrial oxidative respiratory chain complex enzyme and blocked the tricarboxylic acid cycle pathway. In mitochondrial energy metabolism disorders, the release and transfer of the apoptosis-inducing factor and macrophage migration inhibitor to nuclear transfer, chromatin agglutination, and DNA fragmentation mediate parthanatos (Kutuzov et al, 2020). PARP1 also regulates cellular homeostasis and maintains genomic stability under physiological conditions (Rajawat et al, 2020). Considering that PARP1 plays a crucial role in DNA damage induced by oxidative stress, many studies have found that PARP1 plays a crucial role in Lipopolysaccharid- and hydrogen peroxide-induced cell oxidative damage models (Azzouz et al, 2021; Huang et al, 2020).
To verify whether PARP1 plays a role in male infertility caused by oxidative stress induced by VC, we explored the function of PARP1 in male infertility in a rat model with VC- and hydrogen peroxide-induced spermatocyte oxidative stress. Western blot (WB), real-time quantitative PCR (RT-qPCR), immunofluorescence, and flow cytometry were used to explore whether parthanatos was related to male infertility caused by oxidative stress in VC. It may provide new insights for the diagnosis and treatment of VC.
Materials and Methods
Animals
Rats were purchased from Skbex Biotechnology Co. Ltd (Anyang, Henan, China) and kept in a specific pathogen-free environment. Rats were maintained in an environment with 12 h of night/day at 25°C with free access to food and water. This research received ethical approval from the Ethics Committee of Bengbu Medical College.
Establishment of a rat VC model
Twenty SD rats were randomly divided into two groups: the VC group (n = 10) and the control group (n = 10). The rats were anesthetized with 10% chloral hydrate (3.5 mL/kg), and hair was removed from the central abdomen. The abdominal cavity was opened up, and the contents of the abdominal cavity were pushed to the right. The connection between the renal vein and the spermatic vein was then separated. The renal vein was ligated with a 3-0 silk wire and a 0.8-mm metal rod. After obvious stenosis of the renal vein was observed, the metal rod was pulled out to restore the renal vein to 0.8 mm. The abdominal cavity was rinsed with 200,000 units of penicillin, and the contents were restored to their original positions. The peritoneum and pericardium were sutured and disinfected with iodophor (Turner, 1983; Wang et al, 2022b).
Animal tissue preparation
Rats in the VC group and control group were sacrificed for cervical dislocation 8 weeks after surgery. The abdominal cavity of the rats was opened to separate the kidney, testis, and epididymis. Half of the testicular epididymis head was fixed in 4% paraformaldehyde (PFA fix solution) solution for hematoxylin and eosin (H&E) staining, and the caudal epididymis was clipped in 1 mL of phosphate-buffered saline (PBS) solution and a 37°C water bath to allow sperm to swim out sufficiently for sperm count. The remaining half of the testicular tissue was frozen and stored in a −80°C freezer for later RNA and protein extraction.
Histological analyses
H&E staining was used to detect histological changes in the testis and epididymis in the VC rats, and the tissue was fixed overnight in 4% PFA solution. Paraffin-embedded and sectionalized slices were dewaxed three times in
Determination of lipid peroxide and reduced glutathione levels in testicular tissue
The contents of lipid peroxide (LPO) and glutathione (GSH) in the testicular tissue of the 8-week VC group and the control group were detected with an LPO content determination kit (Jiancheng, Nanjing, China) and a trace reduced GSH content determination kit (Jiancheng). In addition, an Enhanced BCA Protein Assay Kit (Beyotime, Shanghai, China) was used to detect the protein content of the tissue homogenate.
VC testicular tissue transcriptome deep sequencing and bioinformatic analysis
RNA was extracted from the control group (n = 3) and VC group (n = 3) and then tested for RNA sample quality, enriched mRNA, and synthetic double-stranded cDNA with breaking RNA. The end of the DNA was repaired, and poly(A) was added to the DNA. Fragments were selected, and PCR amplification was performed. The library quality of Illumina sequencing data was inspected.
The sequences were compared with rat genome fragments. Transcriptional assembly was performed to obtain differentially expressed genes (DEGs), and DEGs were used to obtain heat maps, volcanic maps, and KEGG maps.
Transferase-mediated dUTP nick-end labeling assay
Transferase-mediated dUTP nick-end labeling (TUNEL) was detected with the One Step TUNEL Apoptosis Assay Kit (Beyotime) to show the breakage of DNA. Xylene dewaxing was performed for 5–10 min, before switching to fresh xylene and dewaxing for another 5–10 min. Further soaking was performed in the following sequence: anhydrous ethanol for 5 min, 90% ethanol for 2 min, 70% ethanol for 2 min, and distilled water for 2 min. DNase-free protease K drops (20 μg/mL) were applied at 37°C for 15–30 min and washed with PBS three times. Samples were incubated with 50 μL of TUNEL solution at 37°C for 60 min away from light and washed with PBS three times. The tablets were sealed with antifluorescence quenching solution and observed under a fluorescence microscope.
Cell culture and treatment and construction of an oxidative stress model
GC-2 cells were cultured in DMEM (Life Technologies) with 10% fetal bovine serum (Thermo, Rockford) and 1% penicillin and streptomycin (100 U/mL of penicillin and 100 μg/mL of streptomycin (Beyotime) and incubated at 37°C and 5% CO2. The cells were divided into two groups: the normal control group: normal medium culture and no drug treatment; and the hydrogen peroxide group: GC-2 cells were treated with 0, 200, 300, and 400 μM of hydrogen peroxide for 30 min and 0, 100, 200, 300, 400, and 500 μM of hydrogen peroxide for 24 h.
CCK8 assay
Cells were cultured with different mediums of hydrogen peroxide in the 96-well plate for 30 min and 24 h, and then the medium was removed and washed with PBS. Subsequently, 100 μL of CCK8 (Beyotime) mix (10 μL of CCK8 and 90 μL of DMEM) were added to each well and incubated for 1 h. Cell-free blank control plates with the same volume of CCK8 and basal medium were prepared, and each well was measured with a microplate reader at 450 nm OD. The growth curve was drawn according to the measured OD value.
Cell apoptosis and the cell cycle
Cell apoptosis and the cell cycle were analyzed with a flow cytometer. To assess cell apoptosis, cells were collected after treatment with hydrogen peroxide for 30 min and 24 h. An Annexin V-FITC Apoptosis Detection Kit (Beyotime) was used to stain the cells and determine apoptosis in each group. To assess the cell cycle, cells were collected after treatment with hydrogen peroxide for 30 min and 24 h and fixed overnight with 70% ethanol. Then, 100 μg/mL of RNase A (Beyotime) and 25 μg/mL of propidium iodide (PI) (Beyotime) were added, and the cells were incubated in a warm bath at 37°C for 30 min.
RNA isolation and RT-qPCR
TRIzol (Invitrogen, CA) was used to extract RNA from testicular tissue and GC-2 cells. After removing genomic DNA, mRNA was reverse transcribed into cDNA using the PrimeScript RT reagent kit (Accurate, Changsha, China). RT-qPCR was performed using the ABI System with the SYBR Premix Ex Taq II kit (Accurate). The relative expression levels of Arntl2, Cox8b, HAMP, CFB, Pax8, Chrnb3, catalase, NRF-2, GPX1, GSR, SOD3, and TXNRD1 mRNA were normalized using the endogenous GAPDH mRNA. Primers for genes are listed in Supplementary Table S1.
Protein extraction and Western blotting
Testicular tissue and GC-2 cell proteins were lysed in Radio Immunoprecipitation mix buffer (Millipore, Billerica), complete TMEDTA free protease inhibitor cocktail (Roche, Mannheim, Germany), and phosphatase inhibitors. Samples were centrifuged at 12,000 rpm at 4°C for 10 min, and the supernatant with the protein content was detected by Western blotting. The protein spots were loaded on the Sodium Dodecyl Sulfate PolyAcrylamide Gel glue and transferred to nitrocellulose membranes (Biosharp, Hefei, China). The primary antibody was incubated overnight, and the secondary antibody was incubated for 2 h. An enhanced chemiluminescence substrate (Thermo) was used to visualize the luminescence substrate. Protein levels of PARP1 (ABclonal, Wuhan, China), γ-H2AX (ABclonal), and PAR (CST, Boston) were standardized with β-actin (ABclonal) as the control.
Immunofluorescence
GC-2 cells were cultured in 24-well plates. When the cells grew to 70%, a medium containing different concentrations of hydrogen peroxide was added for 30 min and 24 h. After removing the medium, PBS was washed twice, fixed with 4% PFA for 20 min, transparent with 0.5% Triton X-100 for 20 min, and sealed with 1% for 40 min. The primary antibody (PAR and γ-H2AX) was incubated for 2 h, and the fluorescence secondary antibody was incubated for 1 h. DAPI staining was performed for 2 min, and photographs were taken by a living cell workstation.
Statistical analyses
Each experiment was carried out at least three times, and the results were provided as mean ± standard deviation. The differences between the two groups were analyzed using the Student's t-test. Statistical significance was defined as a p-value of <0.05. SPSS Statistics version 20.0 was used for all statistical analyses.
Results
VC results in the decline of testicular parameters in rats
The diameter of the spermatic veins in the VC group was obviously expanded (0.415 ± 0.0081 vs. 1.328 ± 0.069) (Fig. 1A, D), and the physical ratio of the testis (3.825 ± 0.143 vs. 1.925 ± 0.106) (Fig. 1B, D) and epididymis (2.108 ± 0.128 vs. 1.318 ± 0.033) (Fig. 1B, E) was significantly decreased. Sperm count (2.05 ± 0.521 vs. 0.777 ± 0.426) decreased significantly (Fig. 1G), but there was no significant difference between the two kidneys (1.702 ± 0.038 vs. 1.692 ± 0.025) (Fig. 1H) after 8 weeks. The GSH content decreased (Fig. 2D), and LPO increased in the testicular tissue of VC rats (Fig. 2E). The VC group had irregular spermatogenic tubules, a thinner spermatogenic epithelium, and a vacuolated germinal epithelium, and no elongated sperm or round sperm was found in the lumen compared with the control group. Spermatozoa decreased and were apoptotic in the epididymis (Fig. 2A).

Testicular histology and sperm quality in VC rats.

Morphological and histological examination of testis in VC rats.
The TUNEL test showed that the positive signal of TUNEL increased significantly in the VC group (Fig. 2B).
RNA sequencing of differential genes in VC rats
Three testicular tissues from the VC group and control group were prepared for RNA sequencing, and the heat map showed the difference in gene expression between the VC group and the control group (Fig. 3A and Supplementary Data). The volcano figure showed that 3736 upregulated genes and 94 downregulated genes were significant compared with the control group (Fig. 3B). KEGG showed that DEGs were enriched in the cellular processes, such as apoptosis and lysosome, and in the classic signaling pathways, such as PI3K-Akt and MAPK (Fig. 3C). RT-qPCR verified the upregulated and downregulated genes in the rat RNA sequencing data. The expression of DEGs in the VC testis was the same as in RNA sequencing (Fig. 3D, E).
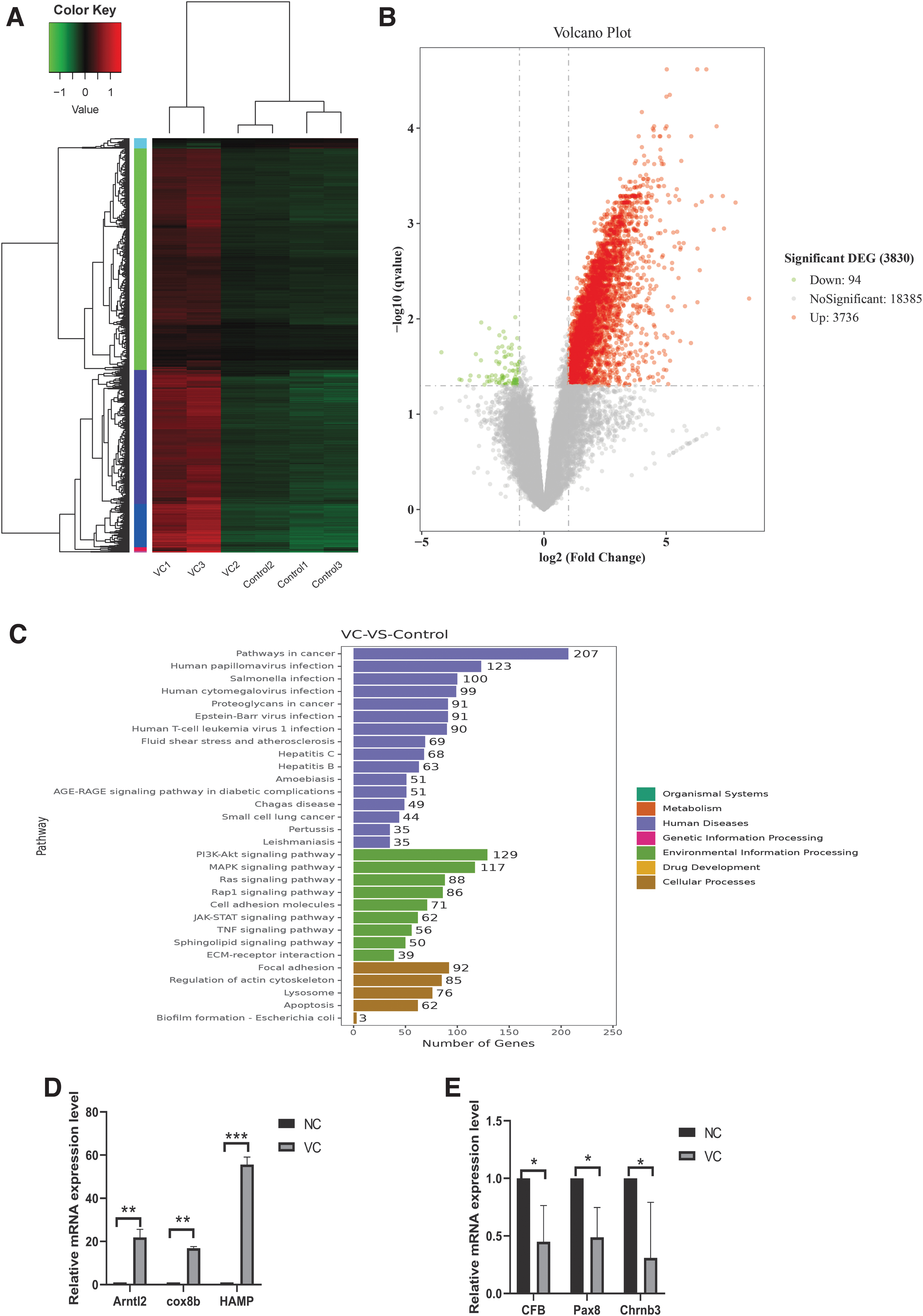
RNA sequencing and differential gene expression analysis and validation in VC rats.
Parthanatos occurred in GC-2 cells with the treatment of hydrogen peroxide for 30 min
GC-2 cells were treated with 0–400 μM hydrogen peroxide for 30 min, and cell proliferation was detected using the CCK8 method. No statistical differences were observed in the decrease of cell viability in the 200 and 300 μM groups, while the decrease in the 400 μM group was statistically significant (Fig. 4A). WB results showed that the expression levels of PARP1, PAR, and γ-H2AX began to increase significantly in the 300 μM group (Fig. 4B), and the immunofluorescence results were consistent with the WB results. The expression of PAR and γ-H2AX increased in the 300 μM group and was concentrated in the nuclear region (Fig. 4C, D).

Parthanatos occurring in GC-2 after 30 min with the treatment of hydrogen peroxide.
Apoptosis occurred in GC-2 cells with the treatment of hydrogen peroxide for 24 h
GC-2 cells were treated with 0–500 μM of hydrogen peroxide for 24 h, and the number of cells decreased significantly (Fig. 5A). Cell proliferation was detected using the CCK8 method. The decrease in cell proliferation was positively correlated with the hydrogen peroxide concentration (Fig. 5B). The apoptosis rate of GC-2 cells was consistent with the increase in hydrogen peroxide concentration by flow cytometry (Fig. 5C). However, PAR and γ-H2AX were not obviously nucleated in the 24-h group compared with the 30-min group (Supplementary Fig. S1).

Apoptosis occurring in GC-2 cells after 24 h with the treatment of hydrogen peroxide.
Discussion
A number of studies have shown that oxidative stress is an important factor affecting sperm quality because the balance between oxidative stress and antioxidant activity in the testis is disrupted (Sanford et al, 2020). Compared with normal fertile patients, the ROS content in the semen of VC patients is significantly increased. GSH, SOD, and other antioxidant enzymes in the testis are significantly decreased, and LPO, Malondialdehyd, and other peroxide products are significantly increased (Anand et al, 2012). All of these indicators can be associated with a decrease in sperm concentration, sperm motility, and normal sperm forms in men. In our study, our results in rats are consistent with previous data in rats showing a decrease in sperm count and a disturbance of spermatogenic epithelial cells (Razi et al, 2021). This is accompanied by decreased levels of GSH and an increased activity of LPO in testicular tissue, indicating that oxidative stress occurs in VC.
Spermatogenesis is the process by which diploid gamete cells produce haploid sperm (Green et al, 2018). During spermatogenesis, diploid male primordial germ cells develop into haploid sperm through meiosis and differentiation and pass on genetic material to the next generation (Tabuchi et al, 2018). It is regulated by a complex network of interactions between testicular cells. Sertoli cells and peritubular myoid cells produce several substances that control and support germ cell growth and spermatogenesis through the production of androgens by mesenchymal cells (Tang et al, 2016). Changes in the microenvironment, such as heat stress and hypoxia, might disrupt spermatogenesis processes and result in lower male fertility (Henriksen et al, 2020; Wang et al, 2022a).
With the metabolic activity of cells, the production of ROS is inevitable, and low levels of ROS are necessary for spermatogenesis and capacitation (Tang et al, 2019). White blood cells, as an important source of ROS production, are significantly increased in patients and rats with VC, and inflammatory factors, such as IL-1 and IL-16 and NLRP3, are significantly increased, suggesting that inflammation may play a crucial role in VC (Antonuccio et al, 2021; Baazm et al, 2020; Fang et al, 2021). Excessive ROS production leads to oxidative damage to tissue genetic material, lipids, and proteins, and ultimately to decreased fertility (Tang et al, 2019). We used a hydrogen peroxide-induced GC-2 cell oxidative stress model to simulate the damage to spermatocytes caused by oxidative stress in VC rats. Hydrogen peroxide induces cell damage and affects cell proliferation by inhibiting the expression level of antioxidant enzymes and producing excessive ROS.
Some studies have shown that the upregulation of antioxidant system and dysfunctional oxidative phosphorylation in unilateral VC may lead to a shift in the body's redox balance to a more reductive state, leading to reductive stress (Swain et al, 2020). In our study, it was found that the redox-related genes (NRF-2, GPX1, GSR, SOD3, and TXNRD1) showed an upregulation trend, except catalase, an enzyme that catalyzes the degradation of hydrogen peroxide, showed a downward trend. The results may show that the break of antioxidant balance in the testis of VC rats resulted in a state of reductive stress in testis (Supplementary Fig. S2).
In recent years, with the development of deep sequencing and bioinformatics, sequencing data sets have been increasingly uploaded to the NCBI website, and RNA deep sequencing has been used in an increasing number of experiments (van Dijk et al, 2018). Changes in RNA expression between samples can be directly expressed, and DEG enrichment in some pathways can help researchers generate new ideas (Petersen et al, 2017). Bioinformatics was used to analyze the RNAseq sequencing data of VC rats to study the potential factors leading to decreased male fertility in VC, and we found that the TNF signaling pathway and apoptosis pathway were enriched. The PI3K-Akt signaling pathway and MAPK signaling pathway are also enriched. We also found and verified some interesting differential genes from the sequencing data of VC rats. For example, the expression of ferroptosis-associated gene HAMP was significantly increased in the RNAseq results (fold change = 7.05, p < 0.0001).
These genes were validated in the rat model and in the GC-2 cell oxidative stress model induced by hydrogen peroxide, and some DEGs were obtained in the animal and cell models.
Parthanatos, as a different apoptosis from classical caspase-dependent apoptosis, plays an important role in ischemia–reperfusion and some brain diseases (Del Re et al, 2019; Dionísio et al, 2021). During the long course of VC, changes in the testicular internal environment may lead to different kinds of tissue damage and induce different apoptosis (Tang et al, 2019). PARP is a posttranslational modification enzyme in most eukaryotic cells and it can be activated by recognizing fragments of DNA and can be regarded as a receptor for discovering DNA damage. PARP1 accounts for more than 90% of intracellular PARP activity and is a DNA repair enzyme that exists mainly in the nucleus of eukaryotes.
When the ROS content in tissue cells increases sharply, DNA damage leads to the aggregation of PAR in the nucleus, and PARP1 is activated as a molecular receptor of DNA damage to repair damaged DNA or induce cell apoptosis (Rose et al, 2020). The PAR level in GC-2 cells was very low, but the expression level of PAR increased significantly after 30 min of hydrogen peroxide stimulation. Although the proliferation ability of the GC-2 cell was not obviously decreased, the expression of PARP1 was significantly increased as a result of the level of PARP1 in VC testis, the expression of PAR was significantly increased and transferred to the nucleus, and the marker of DNA damage γ-H2AX was significantly increased. The results showed that DNA damage was obvious in cells treated with hydrogen peroxide for 30 min, and DNA damage repair was induced by PARP1.
However, in the group treated for 24 h, the expression of PAR or PARP1 did not increase the same as that of the 30-min group. We found that the expression of PARP1 increased under acute oxidative stress, and DNA damage was obvious in our study. These results suggest that PARP1 may be one of the causes of decreased sperm motility in VC.
In our study, VC decreased the ratio of the testes to the epididymis and the number of sperm. H&E showed spermatogenic cell damage, increased LPO content, and decreased GSH content in the testicular tissue homogenate. DEGs screened from RNA sequencing results were verified in the GC-2 oxidative stress model induced by hydrogen peroxide. PARP1 was significantly increased in the VC rat model and the 30-min hydrogen peroxide treatment group. The expression of PAR and γ-H2AX in the GC-2 treatment group was significantly increased and aggregated in the nucleus.
However, DNA damage still existed after 24 h of hydrogen peroxide treatment, but PARP1 expression did not increase significantly, indicating that PARP1 may play a crucial role in DNA damage repair when oxidative stress begins to damage DNA in the short term, but as the damage continues, DNA fracture exceeds the repair ability of PARP1. DNA damage may be repaired by other pathways or induce apoptosis. PARP1 may play a crucial role in male infertility caused by VC. Our study may provide new insight into the mechanism of male infertility induced by oxidative stress. In addition, these data highlight PARP1 as a potential regulatory biomarker in VC, and parthanatos may play an important role in reducing fertility in VC patients.
Footnotes
Authors' Contributions
K.H. and Y.L. guided this work and reviewed the article. K.W. finished the article. K.W., Y.G., and C.W. help to finish the experiment, Y.G., C.W., and Z.L. collected relevant information. M.L. modified the figures. The final draft was read and approved by all of the writers.
Disclosure Statement
No competing financial interests exist.
Funding Information
This research was funded by the Anhui Provincial Natural Science Foundation of P.R. China (2008085QC111), the Natural Science Foundation of the Higher Education Institutions of Anhui Province (China) (KJ2021A0704), the Anhui Provincial Key Research and Development Project of P.R. China (202104j07020016), the Climbing Plan of Natural Science General Project of Bengbu Medical College (China) (2021bypd003), and the 512 Talent Cultivation Plan for Middle-aged Backbone Teachers of Bengbu Medical College of P.R. China (by51201207).
Supplementary Material
Supplementary Data
Supplementary Figure S1
Supplementary Figure S2
Supplementary Table S1
References
Supplementary Material
Please find the following supplemental material available below.
For Open Access articles published under a Creative Commons License, all supplemental material carries the same license as the article it is associated with.
For non-Open Access articles published, all supplemental material carries a non-exclusive license, and permission requests for re-use of supplemental material or any part of supplemental material shall be sent directly to the copyright owner as specified in the copyright notice associated with the article.