
Abstract
Alternative splicing (AS) can generate distinct transcripts and subsequent isoforms that play differential functions from the same pre-mRNA. Recently, increasing numbers of studies have emerged, unmasking the association between AS and cancer. In this review, we arranged AS events that are closely related to cancer progression and presented promising treatments based on AS for cancer therapy. Obtaining proliferative capacity, acquiring invasive properties, gaining angiogenic features, shifting metabolic ability, and getting immune escape inclination are all splicing events involved in biological processes. Spliceosome-targeted and antisense oligonucleotide technologies are two novel strategies that are hopeful in tumor therapy. In addition, bioinformatics applications based on AS were summarized for better prediction and elucidation of regulatory routines mingled in. Together, we aimed to provide a better understanding of complicated AS events associated with cancer biology and reveal AS a promising target of cancer treatment in the future.
Introduction
Alternative splicing (AS) is a complicated, but intensively guided process at the pre-mRNA level (Baralle and Giudice, 2017). As first unveiled in the 1970s, AS is characterized by conferring one ancestral gene the ability to generate distinct junior transcripts and isoforms (Flavell et al., 1979; Choi et al., 1980; Chen and Manley, 2009). Genome-wide analyses based on next-generation sequencing technology have shown that AS regulation is innate in excess of 90% of human genes (Mortazavi et al., 2008; Martinez-Montiel et al., 2018). Due to the large and orderly internal regulation, AS can be preserved in the process of evolution.
The AS process launches inseparably with a compound spliceosome, which is composed of five small nuclear ribonucleoprotein (snRNP) particles, U1, U2, U4, U5, and U6 snRNPs, which are subsequently recruited to the splice sites flanking each intron. Around each exon-intron junction, there exists a consensus sequence included in the splice site that is exclusively distinguished by the spliceosome (Saltzman et al., 2011; Lee et al., 2014; Zhou et al., 2019). In addition, other sequences are bound by splicing factors, thus forming enhancers or silencers of AS regulation (De Conti et al., 2013; Ohno et al., 2018). In particular, the RNA-binding protein (RBP) family is a class of leading splicing factors due to their specific RNA-binding ability and ability to exert AS regulation of pre-mRNA.
Moreover, two confrontational families among the RBPs, the serine-arginine-rich protein (SR) and heterogeneous nuclear ribonucleoprotein (hnRNP) families, both exert pivotal effects on AS events (Busch and Hertel, 2012; Geuens et al., 2016; Jeong, 2017; Kedzierska and Piekielko-Witkowska, 2017). Multifarious modes of AS occur, and the major five categories, exon skipping, intron retention, alternative 5′ acceptor sites, alternative 3′ donor sites, and mutually exclusive exons, compose the principal AS style (Blencowe, 2006; Pohl et al., 2013). Then, pre-mRNA that has undergone the AS process produces diverse mature RNAs and contributes to isoforms with differential structure and function (Montes et al., 2012).
The effects of AS regulation on human diseases have been widely studied in recent decades (Kelemen et al., 2013; Montes et al., 2019; Yang et al., 2019; Bhadra et al., 2020; Song et al., 2020; Zhang et al., 2021a). With the rapid emergence of various regulators in AS events, there has been increasingly more attention regarding cancer research. Multiple previously unknown splicing factors or downstream spliced transcripts have also been identified through the analysis of big data using computers and bioinformatics tools (Florea, 2006; Conesa et al., 2016; Song et al., 2019).
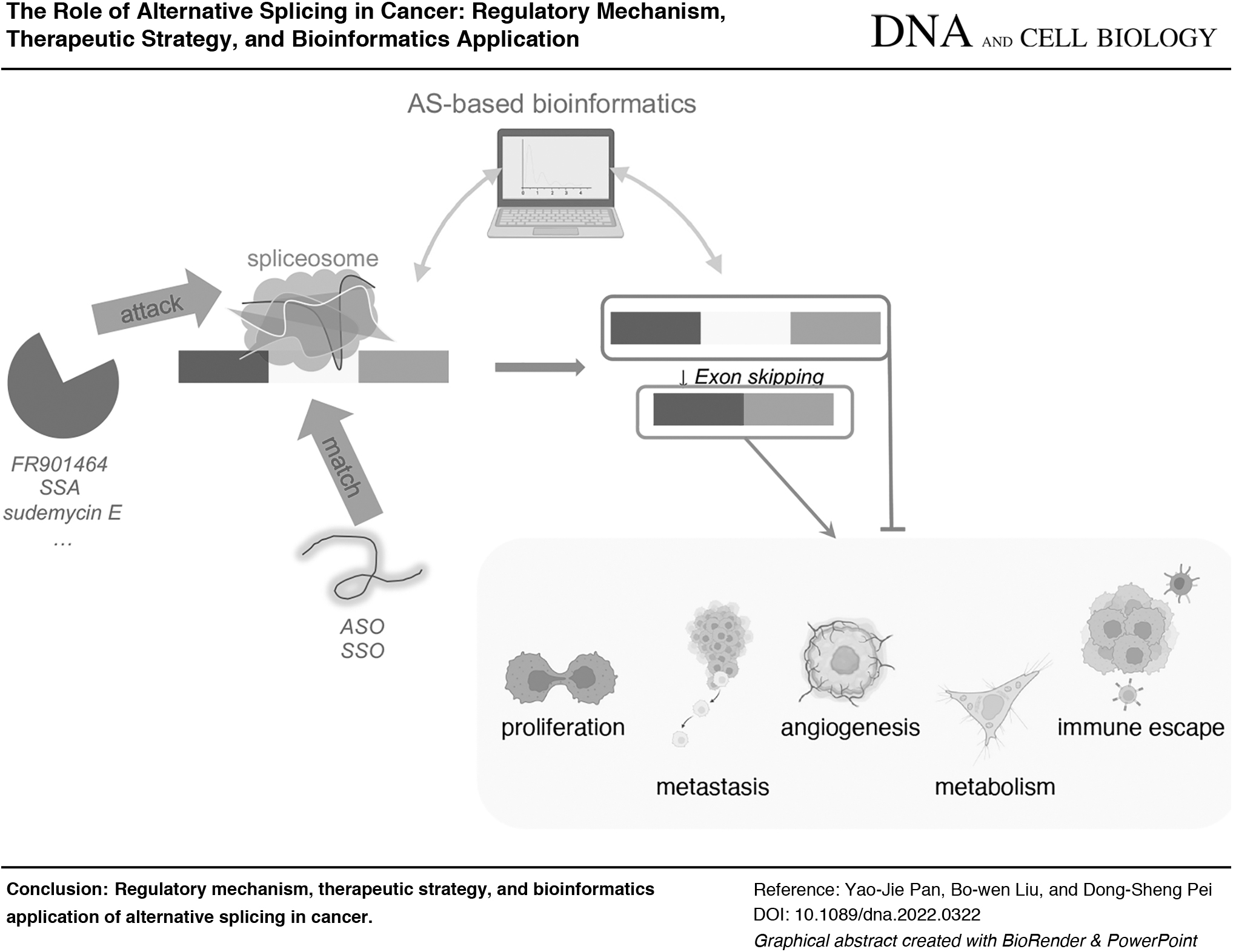
In this study, we collected and sorted out the principal AS activities that occurred during tumor progression. We further unveiled a promising therapeutic strategy targeting AS regulation and displayed bioinformatics applications based on AS to better understand AS-associated networks and support cancer therapy.
The Summary of AS Regulation
The regulation of eukaryotic gene expression is a hierarchical and orderly process (Pope and Medzhitov, 2018; Gil and Ulitsky, 2020). Regarding the transcription process, precursor RNAs (pre-RNAs) are capped, introns are cut off, and poly (A) tails are added to generate mature messenger RNA (Ranish and Hahn, 1996; Proudfoot et al., 2002). During the generation of mature mRNA, all introns should have been cut off theoretically, which is a normal splicing mode named constitutive splicing (Romano et al., 2001; Ding and Elowitz, 2019).
Generally, intronic sequences are excluded, whereas exonic sections are included after the DNA transcript into the pre-mRNA, generating the final mRNA by matching the retained sequences together (Wickramasinghe et al., 2015). Individually, exons being skipped or introns being retained lead to unexpected exon-intron combinations, that is, canonical AS (Ast, 2004; Keren et al., 2010). Importantly, the development of next-generation sequencing technology has uncovered that in excess, 90% of human protein-coding genes experience AS, with ∼80% changes therein altering the protein-coding potential of the transcripts, contributing to protein functional diversity (Mortazavi et al., 2008; Lin, 2015). AS events mainly include five categories: skipped exons, alternative 5′ splice sites, alternative 3′ splice sites, mutually exclusive exons, and retained introns, wherein the decisive roles in determining the occurrence of these events are cis elements and trans-acting factors (Chen and Manley, 2009; Shenasa and Hertel, 2019).
Exon splicing enhancers (ESEs), exon splicing silencers (ESSs), intron splicing enhancers (ISEs), and intron splicing silencers (ISSs) consist of cis elements. These regulatory elements can be recognized by the spliceosome and subsequently assembled by hundreds of proteins and ribonucleoproteins (De Conti et al., 2013; Lee and Rio, 2015; Shih et al., 2019; Zhang et al., 2021b). There is a group of snRNPs, U1, U2, U4/U6, and U5, which interact with the regulatory cis elements that (1) include the 5′ and 3′ SS; (2) correspond to the exon-intron boundaries at each side of the intron; (3) a polypyrimidine tract that lies upstream of the 3′ SS; and (4) an adjacent element known as the branch point sequence.
In particular, the cis elements on pre-RNAs are ready to recruit trans-acting factors, including SR and hnRNP families, which influence splicing through various mechanisms (Kralovicova et al., 2004; Kralovicova and Vorechovsky, 2006; Brant et al., 2019). Due to context-dependent activities, SR proteins could either weaken or strengthen the identification of a peculiar sequence, establishing their identity as both enhancers and splicing silencers. Generally, SR proteins target splicing enhancers, whereas hnRNPs bind to silencers. Unexpectedly, the role of cis- and trans-regulators could inversely affect exon-intron sizes, cellular conditions, and developmental status (Martinez-Contreras et al., 2007; Zhong et al., 2009; Zhou and Fu, 2013; Kedzierska and Piekielko-Witkowska, 2017). For example, a compound of SR proteins SR100 and SRRM4 binds to ISE upstream of the 3′ splice site rather than ESE in microexons, thus exerting the opposite AS function (Chen and Manley, 2009; Raj et al., 2014).
In addition, when SR proteins bind to intronic sequences downstream of target exons, they might unusually exhibit repressive splicing functions (Erkelenz et al., 2013). Except for cis elements and trans factors, the process of AS might be influenced by transcription initiation and elongation, since most splicing activities are performed cotranscriptionally in pre-mRNAs (Naftelberg et al., 2015; Godoy Herz and Kornblihtt, 2019; Li et al., 2019; Giono and Kornblihtt, 2020). If there is an end in the process of AS, the whole process can be evaluated by the concept of a global ratio, the transcription rate, which is responsible for the reading of splicing code inside cells (Stepankiw et al., 2015; Aslanzadeh et al., 2018). Overall, the regulation of AS is complex, but methodical, with in-depth mechanisms to be elucidated.
Aberrant Regulation of AS in Cancers
On the grounds of AS regulation, distinct transcripts or subsequent isoforms were generated to produce multifarious functions (Kelemen et al., 2013). Various reports have determined that the aberrant regulation of AS events is ubiquitous in different developmental processes, including tumorigenesis (Baralle and Giudice, 2017; Climente-Gonzalez et al., 2017; Cherry and Lynch, 2020; Sciarrillo et al., 2020). Especially in cancer, AS-regulated biological processes predominantly embroil cell proliferation, metastasis, angiogenesis, metabolism, and immune escape (Xu and Lee, 2003; Urbanski et al., 2018; Bessa et al., 2020; Bonnal et al., 2020) (Table 1).
Alternative Splicing Events Related to Tumor Process
FGFR2, fibroblast growth factor receptor 2; HBV, hepatitis B virus; HIF-α, hypoxia-inducible factor-α; JMJD6, JuMonJi domain-containing protein-6; KLF4, Krüppel-like factor 4; PD-L1, programmed death ligand 1; PSF, PTB-associated splicing factor; PTBP1, polypyrimidine tract-binding protein 1; RBM, RNA-binding motif; WTAP, Wilms' tumor 1-associating protein.
Cell proliferation
A massive number of studies have mined out crucial splicing factors regulating AS of target pre-mRNA critical to cell proliferation, mainly by modulating the formation of individual AS isoforms or interacting with other splicing factors (Kelemen et al., 2013; Wang et al., 2020a).
As initially discovered as crucial regulators of AS activities, SR and hnRNP family proteins have been extensively reported to alter cell proliferation by AS (Li et al., 2015; Shi et al., 2018; Chen et al., 2019; Wang et al., 2021b; Wang and Jiang, 2021). For example, SCASC4 was alternatively spliced in SRSF1-overexpressing cell lines, resulting in a longer isoform, CASC-4-FL, which contributed to increased proliferative ability (Anczukow et al., 2015). SRSF2 could alternatively splice GCH1 and STK39 into longer isoforms to promote tumor cell proliferation (Luo et al., 2017).
In regard to SRSF3, the regulation of cell proliferation was reported to coordinate with TDP43 in breast cancer (BC) cell lines. The SRSF3/TDP43 complex manipulates exclusive splicing events of exon 12 from both NUMB and PAR3 pre-mRNAs (Ke et al., 2018). Regarding hnRNP, miR-206 overexpression in CRC induces AS switching from PKM2 to PKM1 by targeting hnRNPA1 (David et al., 2010; Fu et al., 2020). DAZAP1, an RBP, antagonizes the splicing regulation of hnRNPA1 with MEK/Erk-mediated cell proliferation (Choudhury et al., 2014).
Another hnRNP, hnRNPK, confers an overwhelming ratio of the exon 3-containing isoform of MRPL33 to the shorter isoform, contributing to the intense proliferative ability and weak apoptotic event in cancer cell lines (Liu et al., 2018a). Mechanistically, the spliceosome machine that propels splicing activities is essential to all splicing processes since these are crucial components of snRNPs (Wahl et al., 2009). In particular, there exists a protein complex named SF3B consisting of seven particles, which is critical to splicing reactions (Teng et al., 2017). Inhibition of SF3B1 was found to hinder cell proliferative ability and promote cell apoptosis, resulting in the arrest of the cell cycle by regulating HOXA10 splicing in gastric cancer (GC) (Zhang et al., 2020b). Another composition, SF3B3, was reported to stimulate the inclusion of EZH2 exon 14 and play a proproliferative role in renal cell carcinoma (RCC) (Chen et al., 2017).
In addition to SR and hnRNP proteins, numerous RBPs also alter cell proliferation by binding to AS target RNAs. Polypyrimidine tract-binding protein 1 (PTBP1) is a multifunctional RBP that is abnormally upregulated in tumors and has a vital role during AS reactions (Takahashi et al., 2015). Most of its AS regulation of tumor cell proliferation is mainly involved in gliomas. Knockout of PTBP1 significantly enhanced the RTN4 variant with the inclusion of exon 3, and amplification of the longer isoform containing exon 3 shrunk cell proliferative activity (Cheung et al., 2009). In addition, PTBP1 increases the truncated isoform of ADAR1 to maintain glioma proliferation by an IRES-like element (Yang et al., 2015). In addition, PTBP1 affects apoptotic and proliferative abilities through AS of a member of the antiapoptotic BCL2 family, Mcl-1, which is a critical regulator of intrinsic apoptosis activity (Cui and Placzek, 2016).
In addition, RBMs (RNA-binding motifs) are a cluster of motifs that can also bind to RNA and are involved in AS regulation. Several RBMs are capable of antagonizing the AS effects exerted by SR factors, thus altering the cell proliferation ability. NUMB has been determined to be an AS target of RBM5/6 and RBM10 and influences the regulation of cell proliferation (Lu et al., 2015). In contrast, RBM5/6 binds to the exonic regions adjacent to splice sites, whereas RBM10 binds to the intronic splice sites, thus exerting the opposite role in recruiting splicing components and assisting recognition (Bechara et al., 2013; Hernandez et al., 2016). Including or losing exon 9 results in a difference among the NUMB proline-rich domains, thus interfering with the interaction between isoforms that regulate cell proliferation (Verdi et al., 1999; Toriya et al., 2006).
Loss of RBM10 in lung adenocarcinoma enhances the expression of EIF4H-L, rather than the shorter isoform EIF4H-S, which contributes to cell proliferation (Zhang et al., 2020c). RBM4 was found to antagonize the splicing of SRSF1 by competitively binding to target pre-mRNAs including BCL-X. Consequently, the upregulated ratio of proapoptotic BCL-XS to antiapoptotic BCL-XL leads to enhanced cell proliferative ability (Markus et al., 2016). Although most reports laid the proliferative foundation of RBM4, a later study supplemented that RBM4 might also suppress cell proliferative ability by regulating the AS of TEAD4 pre-mRNA to generate a truncated isoform, TEAD4-S (Qi et al., 2016).
Other proteins have been found to play the role of splicing factors, including QKI, ZNF326, and PRMT5. QKI, one kind of splicing factor, was reported to regulate the AS of macroH2A1, resulting in upregulated levels of macroH2A1.1 rather than macroH2A1.2 and suppressing tumor cell proliferation (Novikov et al., 2011). In addition, QKI could limit cell proliferation by AS of NUMA1 exon 16. Antagonistically, MBNL1 facilitates the proliferative ability by inducing the exon skipping of NUMA1 exon 16 (Sebestyen et al., 2016). ZNF326, a Zn-finger protein, is involved in the AS regulation of a subset of genes, such as REPIN1/AP4, ST3GAL6, TRNAU1AP, and PFKM. Mechanistically, the PRMT5/WDR77 complex methylates ZNF326 and modulates its splicing function (Rengasamy et al., 2017). Furthermore, knockdown of PRMT5 confers AS regulation on MDM4, the crucial regulator of p53, thus bringing about the inhibition of cell proliferation (Gerhart et al., 2018).
Cooperatively, the DHX9-mediated interaction between NONO and SFPQ regulates the AS of BIN1 and affects the proliferation and migration of hepatocellular carcinoma. The formation of the NONO/SFPQ complex shifts the production of BIN1-S to BIN1-L, while transforming a tumor suppressor to an oncogene (Hu et al., 2020a). Interestingly, a newly discovered protein, WTAP (Wilms' tumor 1-associating protein), is considered indispensable for regulating the cell cycle and proliferation since it autoregulates AS of its own pre-mRNA by binding to SR proteins (Horiuchi et al., 2013).
Although the interaction between splicing factors and splice isoforms has been extensively studied in the past few decades, important isoforms converging on cell proliferation have not yet been linked with splicing factors. Ki-67 is a well-known marker in proliferation-related research (Menon et al., 2019). Five variants were generated from the precursor to the mature RNA of Ki-67, including Ki-67-α, Ki-67-β, Ki-67-γ, Ki-67-δ, and Ki-67-ɛ, and these variants are common in tumor tissues and cell lines. Distinct splicing variants obtain differential functions, in which the α-variant functions as the molecular switch for regulating cancer cell proliferation (Chierico et al., 2017). Mechanistically, exon 7, which distinguishes the α-variant from other isoforms, plays an antiproliferative role by inducing a G1 to G0 transition of the cell cycle. Losing this exon results in the permanent cell cycling of cancer cells and enhanced proliferative ability (Schmidt et al., 2004).
In addition, the shorter variant of somatostatin receptor subtype-5, SST5TMD4, was reported to enhance proliferation in most cancer cell types and blunt the antiproliferative response to some medicines. Therein, the truncated variant catalyzed the PI3K/AKT and MEK/ERK signaling pathways to regulate certain cancer-associated genes, such as MMPs, MKI67, ACTR2/3, and CD24/44 (Del Rio-Moreno et al., 2019). Another gene, Krüppel-like factor 4 (KLF4), was reported to exert a bidirectional role in tumorigenesis until the underlying AS regulation was unveiled. The enhancement of the KLF4α/KLF4 (full length) ratio is positively correlated with the proliferative ability of cancer cell lines (Ferralli et al., 2016). However, the upstream regulators that participate in these splicing events remain to be unveiled.
Some drugs are also implicated in the reactions of AS affecting cancer cell proliferative ability. Paclitaxel is a commonly used antitumor drug in the clinic and was recently found to participate in diverse cancer-associated AS reactions involving the DNA damage response and repair and transcription (Liu et al., 2017). In particular, paclitaxel exerts splicing of ECT2 pre-mRNA, a fundamental factor concerted in cytokinesis activities. Concisely, paclitaxel upholds the generation of the truncated ECT2-S isoform, thus suppressing the cell proliferative ability in cancer (Zhu et al., 2018a).
In addition, synergistic treatment with epigallocatechin-3-gallate (EGCG) and ibuprofen was reported to restrain prostate cancer (PC) cell proliferation. EGCG+ibuprofen treatment in PC cell lines brought about an altered proportion of the spliced isoforms of BCL-X and MCL-1, diminishing the levels of antiapoptotic BCL-XL and MCL-1 L mRNAs with a parallel rise of proapoptotic BCL-XS and MCL-1S mRNAs. Further experiments found that cells treated with only EGCG or ibuprofen failed to generate different splicing variants. This might be attributed to protein phosphatase 1 (PP1), which is activated in the case of the combination of these two drugs and subsequently regulates the phosphorylation of certain SR factors, thus influencing AS reactions (Kim, 2008).
Cell metastasis
Deteriorated metastasis causes up to 90% of malignancy-related deaths, so it turns out gradually as one of the widely accepted hallmarks of aggressive tumors (Suhail et al., 2019). Metastasis entails a vital process named epithelial-mesenchymal transition (EMT). Characterized by transdifferentiation from an epithelial to mesenchymal phenotype, the EMT process was verified to confer deteriorated cell metastasis and aggravate distant invasion (Pastushenko and Blanpain, 2019). Recently, this transdifferentiation process has been determined to be involved in AS regulation (Roy Burman et al., 2021). Various studies have determined several significant splicing factors involved in the EMT process, including SRSF, ESRP, RBFOX2, and MNBL1 (Klingbeil and Isacke, 2011; Biamonti et al., 2012; Pradella et al., 2017).
The first member of the SR family, SRSF1, was explored to play a role in EMT progression soon after its promotion of tumorigenesis comes to light (Gout et al., 2012). Because of the continuous serine and arginine in its RS domain, SRSF1 is always activated at the post-transcriptional level, in which kinases such as SRPK1 phosphorylate and upregulate it (Gout et al., 2012; Goncalves et al., 2014). Subsequently, SRSF1 localizes to the nucleus, where it exerts AS regulation of multifarious genes associated with tumor metastasis (Nowak et al., 2010; Climente-Gonzalez et al., 2017). Among these regulated target pre-mRNAs, skipping of exon 11 of RON pre-mRNA was the first AS event found to be involved in EMT activation (Ghigna et al., 2005, 2010).
In addition, SRSF1 also increases the generation of Rac1b by promoting cassette exon inclusion in Rac pre-mRNA. Rac1b was found to affect the EMT process by enhancing reactive oxygen species (ROS) and increasing the expression levels of transcription factors of EMT, such as SNAIL (Radisky et al., 2005; Melzer et al., 2019). Another SR member, SRSF3, loss of which was linked with increased expression of the A-isoform of the insulin receptor, promoted the process of EMT in hepatocellular carcinoma (Sen et al., 2015).
The key splicing factor of epithelial identity, ESRP, was decoded to exhibit a position-dependent effect and regulate exon inclusion in the presence of UGG-rich motifs in pre-mRNA targets (Warzecha et al., 2009a, 2010; Yang et al., 2016). One of these targets is fibroblast growth factor receptor 2 (FGFR2), two exons of which were mutually excluded, thus generating two isoforms, IIIb and IIIc. Specifically, ESRP inhibits exon IIIc and promotes the inclusion of exon IIIb, contributing to the production of the epithelial-specific FGFR2-IIIb isoform, which is linked with the EMT process (Warzecha et al., 2009a, 2010; Zhao et al., 2013). ESRP could also cooperate with other RBPs, such as PTBP1, to regulate AS of FGFR2 pre-mRNA (Carstens et al., 2000).
Therefore, the final AS regulation of EMT in one cellular condition depends on the comprehensive repertoire of all splicing factors involved. Similarly, the same splicing factor could also exhibit distinct functions in EMT. For example, an epithelial-specific exon of NUMB pre-mRNA was modulated by ESRP proteins, and skipping of different exons resulted in distinct effects on cell–cell adhesion (Wang et al., 2009; Wang and Li, 2010; Warzecha et al., 2010).
Furthermore, ESRPs could also modulate the EMT process by regulating AS events of pre-RNAs involved in cell polarity or perceived environmental signals, such as SCRIB, KTTLG, and CD46 (Isken and Maquat, 2008; Warzecha et al., 2009a; Warzecha et al., 2009a; Gottgens et al., 2016). Strikingly, certain pre-mRNA targets regulated by ESRP generate isoforms that can collaborate reciprocally. Collectively, these results indicate that ESRPs virtually manipulate a network of EMT regulators with complicated AS regulation contributing to the promotion or inhibition of EMT progression.
Except for SRSF1 and ESRP proteins, there are other splicing factors contributing to EMT-relevant AS events, such as RBFOX2 and MNBL1, both of which are salient regulators of the mesenchymal splicing signal (Venables et al., 2013). RBFOX2 was reported to exert AS regulation of a polarity protein (PAR3) and an organizer of actin cytoskeleton (Cortactin), thus modulating the EMT process (Shapiro et al., 2011; Braeutigam et al., 2014). In particular, RBFOX2 could promote the generation of epithelial-specific isoforms as well as mesenchymal-specific isoforms under different conditions (Hall et al., 2013; Yang et al., 2016; Quentmeier et al., 2018). This seemingly contradictory scenario shapes an intricate network among AS events and splicing factors therein.
Although a series of splicing factors involved in EMT have been unveiled, various spliced transcripts or isoforms unspecified for upstream splicing factors, including CD44, remain. It dates back to the 1990s, when the earliest AS variants were reported in BC cells, opening the door of AS research associated with tumors (Tanabe et al., 1993). CD44 is a multistructural and functional transmembrane molecule that is programmed by a 19 exon gene in humans (Chen et al., 2018). Generally, the pre-mRNAs of CD44 contribute to two categories of isoforms, namely, the standard (CD44s) and variant (CD44v) isoforms. CD44s is integrated by exons 1–5 and 16–20, while CD44v alternatively includes exons 6–15 (Zhang et al., 2019a).
As verified as an adhesion-related molecule, CD44v is mainly expressed in epithelial cells, while CD44s is distributed in mesenchymal cells, which links the enhancement of CD44s with the promotion of EMT. HNRNPLL, an endorsed antimetastatic gene, has been found to bind to CD44 and exert an AS reaction to curb the production of CD44 with variable exon 6 (CD44v6), contributing to the inhibition of cancer metastasis (Sakuma et al., 2018). A-Kinase Anchor Protein (AKAP8) antagonizes the splicing activity of hnRNPM, dampening its inhibition of CD44v8 exon inclusion and resulting in cancer metastasis (Hu et al., 2020a). The growth factor TGFβ was depicted to degrade one kind of RBP, PCBP1, to regulate AS and the expression of CD44 (Chen et al., 2021).
In particular, TGFβ upregulates CD44s and downregulates ESRP1 (epithelial splicing regulatory protein 1), one of significant AS regulatory factors in epithelial circumstances, contributing to the induction of EMT (Bhattacharya et al., 2018). Regarding the regulation of CD44v, downregulated ESRP1 is capable of switching the CD44v to CD44s isoform, restraining distant lung metastasis (Yae et al., 2012). Further research indicated that it might be the GU-rich element in CD44 pre-mRNA that attracted and induced the ESRP1 interaction (Larsen et al., 2016). In BC, WNT5A exhibits an antimetastatic role by AS crucial genes associated with cell movement, including CD44 (Jiang et al., 2013). Since the expression level of CD44 is elevated in CSCs (cancer stem cells), it has recently been well accepted as a promising CSC marker (Yan et al., 2015; Gomez et al., 2020).
In addition to the ESRP1 mentioned above, the level of CD44s was positively correlated with the mesenchymal stemness marker ALDH1, which is parallel to the basal phenotype of BC (Inoue and Fry, 2015). A further study based on a mouse model of BC demonstrated that the CD44s isoform could regulate the PDFGRβ/STAT3 signaling pathway to enhance CSC characteristics (Zhang et al., 2019b). In addition, upregulated CD44s contributed to the reduction of metabolic activity of ovarian cancer cells by influencing other CSC traits, such as extracellular acidification and oxygen consumption rates (Bhattacharya et al., 2018).
Other spliced pre-mRNAs involved in the EMT process are as follows: heparanase is an endoglycosidase that degrades and remodels the extracellular matrix and participates in essential biological events in tumorigenesis. The truncated variant of human heparanase lacking exon 5 has been unveiled, but its regulator and effects on tumor cell carcinoma have not been unveiled (Nasser et al., 2007). Aurora-A kinase is a serine/threonine kinase that has a significant function in the mitotic process (Dutertre et al., 2002). Aberrant Aurora-A levels can provoke mitotic faults and generate aneuploidy, thus igniting tumorigenesis.
A study found that Aurora-A containing exon 2 confers CRC more aggressive metastasis (Lai et al., 2020). CDYL2 (Chromodomain Y-like 2) belongs to the CDY family modulating tumor metastasis. The CDYL2a transcript was found to be highly expressed and proinvasive in BC tissues, as well as cells. Deeper research determined that the colocalization of CDYL2a and nuclear speckles contributed to the skipped exon events of numerous target genes, such as FIP1L1, NKTR, and ADD3 (Yang et al., 2020). The prevailing immunotherapy has achieved great success in recent years, especially anti-programmed death-1 (PD-1)/programmed death ligand 1 (PD-L1) immunotherapy. The underlying mechanism is involved in AS reactions. PD-L1 is alternatively spliced and generates different isoforms a, b, and c, in which the c isoform promotes tumor metastasis and becomes a promising therapeutic target for CRC (Wang et al., 2021).
In addition, an open-source database utilizing PSI and DEXSeq procedures identified AS events between metastatic and normal tissues with high frequency. Further crosscheck of these events with another database from TCGA (The Cancer Genome Atlas) revealed two alternatively spliced targets, SERPINA1 and CALD1. The bioinformatics algorithm distinguished that SERPINA1 exon 1 switched between the upstream section and the downstream section differentially, and CALD1 experienced exon skipping of exon 6 in metastatic samples (Lian et al., 2020).
Angiogenesis
Angiogenesis is a crucial process that is characterized by changing from pre-existing vasculature to the formation of new blood vessels (Folkman, 2006). Several studies have revealed that angiogenesis is also controlled by the AS process in multiple fields, such as diabetes, rheumatoid arthritis, and cancer (Otrock et al., 2007; Elshabrawy et al., 2015; Viallard and Larrivee, 2017). The reported pre-mRNA targets participating in AS regulation of angiogenesis are vascular endothelial growth factor (VEGF), neuropilins (NRPs), vasohibins (VASH), and hypoxia-inducible factor-α (HIF-α).
The major stimulatory signal of angiogenesis starts with the binding of VEGF-A to VEGF receptor 2 (VEGFR2) (Gacche and Meshram, 2014). The VEGF-A pre-mRNA contains eight exons, AS of which leads to distinct isoforms, including VEGF-A121/A165/A189 (Peach et al., 2018). As the most abundant isoform, VEGF-A165 was determined to be the most forceful initiator of angiogenesis, while the A189 isoform possessed weakened angiogenic activity (Cheng et al., 1997).
The A165 isoform has been shown to increase the phosphorylation of VEGFR2 as well as the PI3K/AKT and MAPK/ERK signaling pathways (Woolard et al., 2004; Cebe Suarez et al., 2006). Subsequently, a transcriptional coactivator for steroid receptors, CAPER-α, has been reported to exert AS regulation of VEGF-A (Huang et al., 2012). It enhanced angiogenesis by promoting the expression ratio of the A121 to A189 isoform in BC cell lines and A189/A165 in Ewing carcinoma cell lines (Dowhan et al., 2005; Huang et al., 2012). Although the mechanism of differential isoform generation remains unknown, the CA-rich element (CARE) of VEGF-A might shed light on a molecular basis. Previous studies found that hnRNPL binds to CARE, thus preventing the interaction between some miRNAs and their pre-mRNA and subsequently influencing the AS of VEGF-A (Jafarifar et al., 2011).
Therefore, it has remained a significant challenge to dissect the network around CARE and the distinct isoforms of VEGF-A. Another key role for angiogenesis is known as a “decoy” receptor to VEGF, variants of which include i13, i14, e15a, and e15b to date (Ferrara and Davis-Smyth, 1997; Wu et al., 2010; Abou Faycal et al., 2019). It was reported that hnRNPD changed the ratio of differential VEFGR1 isoforms, thereby altering angiogenesis ability, although the AS effects were uncovered to be cell-type specific (Ikeda et al., 2011; Fellows et al., 2013). The clear AS regulation of VEGFR1 falls on JuMonJi domain containing protein-6 (JMJD6), NOTCH1, and SRSF2, with the latter acting as a promoter of the i13 isoform and the other two being inhibitory regulators of the isoform (Boeckel et al., 2011; Kangsamaksin et al., 2015; Abou Faycal et al., 2019).
In addition to VEGF, three other genes, NRPs, VASH, and HIF-α, are involved in the AS regulation of angiogenesis. NRPs are transmembrane coreceptors that play roles in VEGF signaling and are implicated in angiogenesis progression (Bowler and Oltean, 2019). The first family member, NRP1, skipping exon 7, was determined to attenuate angiogenesis (Hendricks et al., 2016). Although the other family member NRP2 was reported to function in lymphangiogenesis, the underlying mechanism of its AS regulation remains unmasked (Yuan et al., 2002). VASH is an inhibitor of angiogenesis that can be regulated by VEGF-A (Sato and Sonoda, 2007). VASH-1 is one of the families and has two isoforms, namely, VASH1A and VASH1B (Li et al., 2020a).
An in vitro study indicated that the VASH1B isoform could inhibit angiogenesis, so we postulated that the generation of the VASH1B isoform is associated with VEGF-A (Horie et al., 2016). Since hypoxia-induced AS has been marked as the 11th hallmark of cancer, hypoxia has recently emerged as a key circumstance in studying AS regulation and indeed aggravates angiogenesis (Farina et al., 2020). One variant of HIF-α, HIF-3α, which is generated by the inclusion of intron 7, prevents the binding of HIF to target elements and inhibits its transcription, weakening the angiogenesis ability (Maynard et al., 2007; Ando et al., 2013).
Metabolism
The metabolic pathways conferring proliferative cells with the required energy in cancer have emerged as indispensable hallmarks in the past few decades (Vander Heiden and Deberardinis, 2017). Many reports have shown that AS regulation has an impact on metabolism in carcinoma cell lines and influences tumorigenesis (Kozlovski et al., 2017). To date, three key enzymes have been reported to be associated with AS regulation in neoplastic metabolism, including pyruvate kinase, glutaminase (GA), and ketohexokinase (KHK) pre-mRNAs.
PKM is one of the isoforms encoded by pyruvate kinase genes, and PKM1 and PKM2 are two splice variants that originate from it (Christofk et al., 2008). The difference between these two lies in exons 9 or 10, which were incorporated in PKM1 and PKM2, respectively. Although both isoforms perform the same catalytic cascade, PKM1 seems to be mainly responsible for pyruvate production, and PKM2 is widely expressed in cancer cell lines (Noguchi et al., 1986). A study indicated that PKM2 is indispensable for aerobic glycolysis, which provides an acquired growth benefit for tumorigenesis (Christofk et al., 2008). Later research found that PKM2 interacts with and transactivates HIF-α to promote oxidative metabolism to glycolysis (Luo et al., 2011).
GA is the initiator starting the hydrolysis of glutamine to glutamate, variants of which exist in two forms, GAC and kidney type GA (KGA) (Cassago et al., 2012; Mates et al., 2019; Masisi et al., 2020). The C isoform is highly expressed in various tumor types, such as CRC, BC, and glioma (Gross et al., 2014; Zhu et al., 2017; Ascencao et al., 2018; Han et al., 2018). Its mitochondrial localization confers higher catalytic activity than KGA, leaving the concrete mechanism unveiled.
The initiator enzyme phosphorylating fructose is performed by KHK, which has two AS variants, KHK-A and KHK-C (Han et al., 2018). A study found that KHK-C facilitates fructose metabolism and decreases intracellular phosphate and ATP levels, producing higher ROS concentrations than glucose metabolism (Ishimoto et al., 2012; Softic et al., 2016). Although the underlying mechanism remains uncovered, we postulate that c-Myc might be involved in the regulatory network since both isoforms are targets of c-Myc and that c-Myc depletion indeed switches the KHK-A to KHK-C isoform (Li et al., 2016a).
Immune escape
Malignant cells have inherited various mechanisms to contribute to boosting immune escape. It is clearly established that the immune system can affect the cancer response to therapy (Gajewski et al., 2013). However, tumor cells are born with an immune escape mechanism to prevent themselves from immune system attack (Vanichapol et al., 2018; Lei et al., 2020). In this respect, AS is increasingly described to affect the immune system. The following are the genes reported thus far to affect tumor development by modulating immune escape.
Notably, the AS of CD45 pre-mRNA might activate T cells by mediating T cell receptor signaling (Trowbridge and Thomas, 1994; Mustelin et al., 2003). The AS events contain the variable inclusion of exons 4, 5, and 6, among which the generated CD45RO isoform promotes termination of the T cell response (Xu and Weiss, 2002). Mechanistically, phosphorylation of the PTB-associated splicing factor (PSF) separates it from CD45 exon 4 and hence brings about the inclusion of exon 4 (Melton et al., 2007). The activation of T cells attenuates this process and stimulates the generation of the CD45RO isoform (Heyd and Lynch, 2010).
The 10th member of the SR protein family, SRSF10, mediates IL1RAP pre-mRNA AS arising from the membrane form of the IL1RAP (mIL1RAP) protein. Furthermore, mIL1RAP induced the “don't bite me” signal CD47 to facilitate immune escape by promoting NF-κB (Liu et al., 2018b).
Distinct from the serine- and arginine-abundant SR members, the SFPQ splicing factor is characterized by rich proline and glutamine, while exerting equally crucial AS regulation on pre-mRNA targets. A study determined that SFPQ switches exon 7 skipping of one transcription factor, IRF1, impairing the tumor microenvironment and deteriorating tumor development (Bernard et al., 2021).
Another AS-related gene, survivin, generates distinct transcripts as follows: survivin, survivin-ΔEx3, survivin-2B, survivin-3B, and survivin-2α (Mahotka et al., 1999; Badran et al., 2004; Caldas et al., 2005). Among them, 3B was found to be a cancer-specific isoform leading to tumor immune escape and subsequent tumor formation and treatment resistance (Li, 2005; Vegran and Boidot, 2013; Vegran et al., 2013).
In addition to the genes mentioned above, hepatitis B virus (HBV) was prominently linked to immune escape in tumor cell lines. HBV RNA can undergo AS and be attributed to HBV pregenomic RNA-interacting proteins; nearly 20% were determined to be tightly associated with the splicing machinery (Suzuki et al., 1989; Wu et al., 1991). Nine splicing factors were uncovered to participate in AS regulation on pregenomic HBV, including hnRNPAB, SF1, La, PSF, and SRSF1/4/5/6/7. As a result, downstream HBV splicing-generated proteins decrease the expression level of CCL2 and affect tumor immune escape (Duriez et al., 2017).
Therapeutic Targeting of AS in Cancer
Targeting spliceosome in AS
As a fundamental role in AS regulation, targeting the spliceosome seems to be a feasible technical direction. As a crucial component of U2 snRNP in the spliceosome, SF3B participates in spliceosome assembly, thus assisting global AS progression (Kaida et al., 2007; Sun, 2020 ). A set of small molecules has been synthesized or found to efficiently target SF3B to modulate the AS process.
FR901464 is the first natural compound thought to inhibit AS and tumorigenesis by targeting SF3B (Nakajima et al., 1996a, 1996b; Kaida et al., 2007; Osman et al., 2011). The derivative of FR901464, spliceostatin A (SSA), has also been found to play an antitumor role by attenuating cell proliferation through a mechanism that affects the AS of cell cycle regulators such as Cyclin A2 (Motoyoshi et al., 2004; Corrionero et al., 2011). Similarly, AS regulation by SSA also targets SF3B to limit spliceosome assembly (Ghosh and Chen, 2013).
Further research unveiled the major participants therein, namely, functional cis elements, trans-acting factors U1 and U2 snRNPs supported by abundant ATP (Kaida et al., 2007; Lo et al., 2007; Jurica, 2008; Roybal and Jurica, 2010). Another two derivatives of FR901464, meayamycin, and sudemycin E, were also determined to target SF3B and interfere with spliceosome formation, both of which showed an antiproliferative function in various cancer cell lines (Albert et al., 2009; Gao and Koide, 2013; Convertini et al., 2014; Kashyap et al., 2015).
Notably, a derivative from sudemycin E, sudemycin D6, can alter the site selection of AS without splicing inhibition ability (Convertini et al., 2014). In particular, it also functions by binding to SF3B and performing AS regulation on MDM2 (Shi et al., 2015). Another derivative of sudemycin E, sudemycin K, was reported to promote MCL1-exon 2 skipping in cervical carcinoma cell lines (Makowski et al., 2017). In addition, pladienolide B (PB) and its analog FD-895 are found to inhibit splicing by interacting with SF3B and exert an antiproliferative ability in cancer cell lines (Seki-Asano et al., 1994; Mizui et al., 2004; Kashyap et al., 2015).
Except for SF3B, other spliceosome-associated proteins, such as FBP21, were reported to be targets of small molecules. Borrelidin is a natural polyketide that limits angiogenesis and metastasis by targeting FBP21 (Wakabayashi et al., 1997; Habibi et al., 2012). Mechanistically, borrelidin switches the AS of VEGF pre-mRNA to generate a high ratio of antiangiogenic isoforms in retinal cells (Woolard et al., 2011). Moreover, there are other small molecules targeting the formation of the spliceosome, such as Madrasin. In cervical cancer cells, Madrasin promotes the exon skipping of cell cycle-related mRNAs to adjust cell cycle arrest, which contributes to the inhibition of cancer development (Pawellek et al., 2014). Casein kinase 2 (CK2) is generally highly expressed in tumors, and a recent clinical trial for cancer treatment indicated that an inhibitor of CK2, CX-4945, exhibits antiproliferative effects by AS of CK2 pre-mRNA and subsequently CK2-modulated PI3K/Akt signaling pathway (Chon et al., 2015).
Antisense oligonucleotide technology
ASOs (antisense oligonucleotides) are a class of short-length base oligonucleotides that bind to the target sequence in a complementary manner (Seki-Asano et al., 199). Theoretically, ASOs can be artificially applied to fine-tune the AS process toward antiproliferative antimetastatic isoforms in cancer therapy. Moreover, ASOs could be optimized to directly target the regions close to certain splice sites due to their high specificity to targets, which turned out as splice-switching oligonucleotides (SSOs). SSOs specifically target cis elements on the pre-mRNA, thus competing with other factors and exerting splicing functions to affect exon and intron definition (Urbanski et al., 2018; Jbara et al., 2021; Raguraman et al., 2021). SSOs were applied to treat cancer by targeting crucial cancer-related genes such as BCL-X.
The BCL-X SSO reduces BCL-XL and enhances BCL-XS levels, thus inducing cell apoptosis and inhibiting tumorigenesis (Li et al., 2016b; Dou et al., 2021). The other cancer-related genes applied in SSO technology include FGFR, HER2, BRCA1, SMN2, and ATM (Pickles, 2001; Wan et al., 2009; Kralovicova et al., 2016; Farrelly-Rosch et al., 2017; Smith et al., 2017). In particular, targeting SR and hnRNP proteins resulted in two specific SSO technologies, namely, ESSENCE (exon-specific splicing enhancement by small chimeric effectors) and TOSS (targeted oligonucleotide silencers of splicing). The ESSENCE application exploits an oligonucleotide specifically bound to the SR proteins in their RS domains to modulate AS events.
The TOSS strategy supplies an oligonucleotide recognized by hnRNP proteins, thus inhibiting splicing events (Sanford et al., 2005). Because of the high possibility of degradation of exogenous oligonucleotides by RNase H, modifications to protect ASOs from degradation have emerged in recent decades, including phosphorodiamidate morpholino oligos and peptide nucleic acids (Sazani and Kole, 2003; Gupta et al., 2016; Smith and Zain, 2019). Although ASO and SSO technologies are cutting-edge, there remain great challenges in how to transport them to specific locations. After solving these problems, the designated SSOs for cancer treatment seem plausible in the future.
AS-Based Applications
Regarding bioinformatics research on AS, many algorithms and tools based on meta-data analysis have been derived, which makes the monitoring and prediction of AS events more convenient (Table 2).
Descriptions of Alternative Splicing-Based Applications
AS, alternative splicing; circRNA, circular RNA; ESE, exon splicing enhancer; SNP, single nucleotide polymorphism; SR, serine-arginine-rich protein.
In 2004, a database named BASD, predicting variants from RefSeq, was unveiled (Hui et al., 2004). Four years later, a French team analyzed the level of disorder of distinct transcript isoforms by evaluating uncertainty with Shannon's entropy. The rule formulated as those genes gaining high entropy appear more frequently in splicing factors (Ritchie et al., 2008). In 2014, a team from Sweden developed a tool named SpliceVista used for splice variant identification and visualization in shotgun proteomics data (Zhu et al., 2014). Another algorithm based on validated data from different platforms to identify splicing events was integrated as a novel algorithm called ESLiM a few months later (Risueno et al., 2014).
In 2016, a new toolbox SplAdder and an innovative junction-centric process jSplice enabled de novo mining of AS reactions from RNA-sequencing data with high credibility, efficiency, and accuracy (Christinat et al., 2016; Kahles et al., 2016). Then, a team proposed a novel differential transcript expression analytic algorithm called SDEAP that directly evaluates the frequency of AS activities among all input examples utilizing a Dirichlet mixture and graphic decomposition algorithm (Yang et al., 2016; Yang and Jiang, 2016). A network-based strategy, DrAS-Net, was developed to explore in excess of 2.5 million variants in tumors and connect somatic mutations with cancer-specific AS activities (Li et al., 2017). Whippet, a convenient RNA-seq evaluation approach that is compatible with a laptop model, was exploited to quantify AS events with high accuracy regardless of any complication (Sterne-Weiler et al., 2018).
In addition to the algorithms designed for detecting AS events, a pancancer library of certified natural and mutated AS processes was shown publicly, which focuses on mRNA splicing mutations (Shirley et al., 2018). Furthermore, a de novo algorithm that determines AS events in circular RNA (circRNA) was developed and became the first informatics algorithm that distinguishes AS events special to cancer-related circRNAs (Feng et al., 2019). Tumor heterogeneity has recently received increasing attention due to its potent role in modulating molecular classifications through specific indicators. Previous studies have utilized gene expression to distinguish cell types. Two years ago, one computational approach-linked AS and cancer heterogeneity to exploit single-cell RNA sequencing data was unveiled (Manipur et al., 2019).
A user-friendly database named CancerSplicingQTL was later constructed to more deeply depict the prospective functional roles of single nucleotide polymorphism that rule the regulation of both transcripts and isoforms in human malignancy (Tian et al., 2019). Considering the differential expression levels of isoforms between normal and tumor tissues, researchers have focused on identifying higher degree isoforms (HDIs) from multi-isoform genes in cancer by performing a meta-analytic structure to evaluate the levels of different isoforms. Then, these HDIs were compared with isoforms identified by proteomic information, thus showing potential as prognostic predictors or therapeutic targets for cancer treatment (Ma et al., 2019).
In addition to HDIs, a portal named GESUR was developed, which also illustrates the detailed prognostic characteristics of transcripts associated with patient prognosis and other isoforms (Tang et al., 2019). The latest analysis of changes in genome-wide patterns of AS and its functional consequences was extended as an R language pack named IsoformSwitchAnalyzeR (Vitting-Seerup and Sandelin, 2019). Another R language pack named VALERIE inspected AS events at the single-cell resolution level (Wen et al., 2020).
In addition, some novel omics approaches have been exploited to coordinate with RNA-based algorithms to identify AS events. In 2013, a peptidomics methodology for proteomics groups to analyze, recognize, and distinguish AS isoforms from mass spectrometry-based proteomics studies was developed with more attention and special emphasis on AS. The methodology assists in deducing potential hypotheses on precancerous risk factors, hence contributing to the distinction of latent AS reactions and the identification of early tumor biomarkers (Zhang et al., 2013). In 2016, a novel peptidomics method was presented for monitoring isoform-specific peptides for clinical cancerous proteomics usage from liquid chromatography tandem mass spectrometry/mass spectrometry (LC–MS/MS) (Zhang and Chen, 2016).
In particular, visual interactions to identify AS regulation during the EMT process have also been constructed. This informatics analysis revealed the intricacy underlying the regulation of RBPs and discovered previously unreported RBPs that are involved in AS regulation of EMT-related activities (Qiu et al., 2020).
Furthermore, networks between AS events and corresponding prognosis infiltrating different cancer types have been described successively, including ovarian cancer, prostatic cancer, endometrial carcinoma, esophageal cancer, thyroid carcinoma, head and neck tumor, BC, melanoma, leukemia, pancreatic cancer, lung cancer, GC, colorectal carcinoma, and RCC (He et al., 2018; Huang et al., 2018; Zhu et al., 2018b; Gao et al., 2019; Lin et al., 2019; Mao et al., 2019; Xing et al., 2019; Zhang et al., 2019b, 2020c; Zuo et al., 2019; Cai et al., 2020; Chen et al., 2020a, 2020b, 2020c; Cheng et al., 2020; Li et al., 2020b; Liu et al., 2020; Lu et al., 2020; Qu et al., 2020; Wan et al., 2020; Wang et al., 2020b; Xie et al., 2020; Yu et al., 2020; Zhao et al., 2020a, 2020b; Ye et al., 2021).
In addition to global AS events, further research focused on splice junctions was unmasked in the past year. The RjunBase database helps uncover novel AS targets and offers calculations to distinguish and evaluate possible neoepitopes for cancer therapy (Li et al., 2021). Furthermore, by utilizing TCGA SpliceSeq data, association analysis was implemented to identify the possible link between solid tumor-specific splicing events and DNA methylation (Sun et al., 2020 ).
Furthermore, exploration of cis-elements through methods of machine self-learning is capable of deriving a “splicing code” that guides splice site selectivity. For example, the intelligent web-based resource ESEfinder (
Conclusions
AS plays a crucial role in tumorigenesis, conferring malignant progression, including abnormal proliferation, endless metastasis, and preventing immune attack. Although genome-wide studies have unveiled the general implications of AS events in several aspects of cancer biology, future efforts are still needed to reveal the potential connections underlying the regulation of gene expression and function. To this end, we reviewed AS activities involved in the hallmarks of cancer cells in this article. We also summarized therapies such as ASO technology that are promising in clinical treatment. In addition, we collected applications related to AS and wished to better understand internal regulation networks.
Footnotes
Authors' Contributions
All authors contributed to the review conception and design. Data collection was performed by B.-W.L. The first draft of the article was written by Y.-J.P. and D.-S.P. All authors read and approved the final article.
Disclosure Statement
The authors declare no conflict of interests.
Funding Information
This work was supported by the National Natural Science Foundation of China (No. 81872080).