
Abstract
Circular RNAs (circRNAs) are a form of RNAs that lack coding potential. The role of such circRNAs in dental pulp stem cell (DPSC) osteo/odontogenic differentiation remains to be determined. In this study, circRNA expression profiles in DPSC osteo/odontogenic differentiation process were analyzed by RNA-seq. qRT-PCR was used to confirm the differential expression of circ_0005044, miR-296-3p, and FOSL1 in DPSC osteogenic differentiation process. Circ_0005044, miR-296-3p, and FOSL1 were knocked down or overexpressed. Osteoblastic activity and associated mineral activity were monitored via alkaline phosphatase (ALP) and alizarin red S (ARS) staining. Interactions between miR-296-3p, circ_0005044, and FOSL1 were assessed through luciferase reporter assays. Finally, an in vivo system was used to confirm the relevance of circ_0005044 to osteoblastic differentiation. As results, we detected significant circ_0005044 and FOSL1 upregulation in DPSC osteo/odontogenic differentiation process, as well as concomitant miR-296-3p downregulation. When knocking down circ_0005044 or overexpressed miR-296-3p, this significantly inhibited osteogenesis. Luciferase reporter assay confirmed that miR-296-3p was capable of binding to conserved sequences in the wild-type forms of both the circ_0005044 and FOSL1. Furthermore, knocking down circ_0005044 in vivo significantly attenuated bone formation. Therefore, the circ_0005044/miR-2964-3p/FOSL1 axis regulates DPSC osteo/odontogenic differentiation, which may provide potential molecular targets for dental-pulp complex regeneration.
Introduction
Many stem-like precursor cells reside within the dental pulp tissue and are capable of regenerating this tissue in response to injury by migrating to the damaged region, proliferating, undergoing odontoblastic differentiation, and producing reparative dentin (Lambrichts et al, 2017; Wang et al, 2011). When these stem-like cells are isolated and grown in vitro, they are referred to as dental pulp stem cells (DPSCs) (Hilkens et al, 2013). These DPSCs are relatively heterogeneous in composition, containing a range of adipocytes, chondrocytes, and odontoblast-, osteoblast-, and neural-like cells (Lambrichts et al, 2017; Wang et al, 2011). DPSC odontoblastic differentiation is vital to mineralized matrix production during the regeneration of the dental pulp and during the dentinogenic process. Earlier studies have characterized some of the mechanisms regulating DPSC differentiation (Geng et al, 2017; Qin et al, 2012), but further clarification of the underlying molecular mechanisms is still required.
The odontoblastic differentiation of DPSCs has been shown to be regulated by miRNAs (Sun et al, 2014). In one study, for example, the let-7b miRNA was shown to suppress MMP1 expression so as to post-transcriptionally regulate DPSC osteogenesis (Wang et al, 2018b). One prominent theory proposes that such miRNAs are in turn regulated by competing endogenous RNAs (ceRNAs), forming a complex network of coregulatory noncoding and coding RNA molecules (Huang et al, 2015). These ceRNAs are able to bind miRNAs through their competitive miRNA response elements, effectively sequestering them and thereby modulating any normal or disease-associated processes linked to these miRNAs (Song et al, 2017).
Circular RNAs (circRNAs) are a form of ceRNA that were originally thought to be a product of splicing errors (Nigro et al, 1991), but that have more recently been shown to be functional and widespread throughout animal cell types (Memczak et al, 2013). These circRNAs are most often composed of highly conserved exon sequences that are often expressed in patterns that are specific to a given tissue or stage of development (Barrett et al, 2015; Salzman et al, 2013). Owing to their circular nature, circRNAs are extremely stable and can resist the degradative activity of RNases (Memczak et al, 2013; Salzman et al, 2013). This stability makes circRNAs potentially ideal as a diagnostic tool in numerous pathological contexts.
At a functional level, many circRNAs have also been found to modulate intracellular signaling in a number of different cancer types (Qi et al, 2015). Of note, one recent study identified 24 differentially regulated circRNAs that were specifically linked to stages of osteoblastic differentiation, suggesting that these particular circRNAs may play a role in this differentiation process (Dou et al, 2016). The specific function of circRNAs in the context of human DPSC odontogenic differentiation, however, remains unclear. To date, a single study found that 1314 and 1780 circRNAs were up- and downregulated, respectively, during this process (Li and Jiang, 2019), and no studies to date have generated a functional circRNA/miRNA/mRNA network associated with this process.
In this study, we used an RNA-seq approach in an effort to identify circRNAs that were differentially regulated in the context of DPSC osteo/odontogenic differentiation. This led us to identify circ_0005044 as an important regulator of this differentiation process in human DPSCs. Together, our findings offer valuable new insights into the mechanistic basis for DPSC osteo/odontogenic differentiation, potentially highlighting novel therapeutic avenues for future tissue regeneration-based studies.
Materials and Methods
Cell culture
The Ethics Committee of the Stomatological Hospital, Southern Medical University approved all human protocols described herein (Approval No.: 2016006). For this study, we utilized premolars and third molars without any carious lesions that had been extracted from eight healthy donors (five female, three male; age: 12–25 years) in July 2016. These teeth were extracted for orthodontic or impacted third molar reason the Department of Oral and Maxillofacial Surgery at the Stomatological Hospital, Southern Medical Hospital (Guangzhou, China). All patients provided informed consent. Using these samples, we isolated and cultured human DPSCs as in previous analyses (Liu et al, 2020).
HEK-293T cells were obtained from the American Type Culture Collection (ATCC). Both hDPSCs and HEK293T cells were grown in a 5% CO2 humidified incubator using general medium (GM), which is α-MEM (Gibco) containing 15% FBS (Gibco) and penicillin/streptomycin (Sigma-Aldrich). DPSCs were cultured until 70–80% confluent, at which time odontogenic differentiation was induced by culturing cells in osteo/odontogenic induction medium (OM) that had been supplemented with 100 nM dexamethasone, 200 μM
RNA-seq and bioinformatic analyses
DPSCs cultured in GM (control group) or OM (osteo/odontogenic group) for 14 days were used for RNA-seq, each group contained three samples. An Illumina HiSeqseq™ 2000 instrument (Illumina, Inc.) was used to sequence RNA samples. Raw data were initially filtered, after which circRNA predictions were made using the CIRI software (v1.2), and the annotation of these circRNAs based on their source was conducted with circBase (
qRT-PCR
TRIzol (Invitrogen) was used to extract cellular RNA based on provided instructions, after which a SuperScript III®kit (Invitrogen) was used for reverse transcription. All qRT-PCR were conducted with an ABI PRISM 7500 Sequence Detection System (Life Technologies) together with the Power SYBR Green Master Mix (Applied Biosystems). Relative mRNA expression was normalized based on GAPDH levels, whereas U6 was used for circRNA and miRNA normalization. Thermocycler settings were as follows: 95°C for 3 min; 40 cycles of 95°C for 15 s, 60°C for 40 s. Sangon Biotech (Guangzhou, China) synthesized all primers used in this study (Table 1). The 2−resenmethod was used to quantify relative gene expression from triplicate assays (Livak and Schmittgen, 2001).
Primers Used in This Study
RNA oligoribonucleotides
GenePharma Co. (Shanghai, China) was the source of all RNA oligoribonucleotides used in this study, including miRNA mimics, miRNA inhibitors, siRNAs specific for cir_0005044 or FOSL1 (si-FOSL1), and appropriate control constructs (Table 2).
RNA Oligoribonucleotide Sequences Used in This Study
Cellular transfection
Cells were allowed to grow until reaching 70–80% confluence, at which time they were transfected with appropriate constructs (miR-296-3p mimic, miR-296-3p inhibitor, si-circRNA, si-FOSL1, or appropriate controls; 100 nM) using Lipofectamine 2000 (Invitrogen) according to the instructions provided. Cells were then allowed to rest for 48 or 72 h, after which mRNA and protein expression was analyzed.
ARS and ALP staining
DPSCs were initially plated into 24-well plates in a 500 μL volume and were allowed to grow until 70% confluent. Cells were then cultured in OM for either 7 or 14 days for ALP or ARS staining, respectively. Following these culture periods, cells were fixed for 20 min using 4% paraformaldehyde, after which they were stained using 2% ARS or BCIP/NBT ALP Color Development Kit (Beyotime) based on provided directions, the manufacturer's instructions. ALP activity was additionally quantified using a colorimetric assay kit (BioVision). For this approach, cells were washed using PBS before lysis in 1% Triton X-100 (Sigma-Aldrich) and addition to distilled water. Absorbance at 405 nm was then quantified. A BCA kit was used to measure total cellular protein (Thermo Fisher Scientific), with ALP activity being determined based on the ratio of absorbance to protein levels. Mineralized nodule formation was quantified by dissolving the ARS stain for 1 h using 1 mL 10% cetylpyridinium chloride (Sigma-Aldrich), after which absorbance at 570 nm was quantified and normalized to total protein levels.
Western blotting
Protein was extracted from DPSCs via RIPA lysis buffer (Solarbio, Beijing, China). BCA protein assay kit (Pierce, Rockford, IL) was used to identify the total concentration of the proteins. Next, the loading buffer was added with the extracted protein (30 μg), followed by denaturation of the protein in boiling water (for 5 min). Ten percent (w/v) SDS-PAGE was used for the protein separation. Then the protein was transmitted to the PVDF membrane.
For blockage purposes, BSA (5% w/v; Sangon Biotech) was used for 60 min at ∼25°C, followed by membrane probing with anti-RUNX2 (Cell Signaling Technology), anti-FOSL1 (Cell Signaling Technology), or anti-GAPDH (Cell Signaling Technology), and anti-rabbit secondary antibody (Cell Signaling Technology) for 24 h at 4°C. After washing with Tris-buffered saline with Tween-20 (TBST), the incubation of membranes was carried out with the secondary antibodies (Cell Signaling Technology) at ∼25°C for 60 min and these antibodies were conjugated with horseradish peroxidase with 1: 3000 dilution. TBST was used to wash unbound antibodies. The ECL imaging kit (Thermo Fisher Scientific) was used to generate the chemiluminescent signal. The internal control was GAPDH (Supplementary Fig. S1).
Luciferase activity assay
Direct interactions between miR-296-3p and either circ_0005044 or FOSL1 were assessed via a luciferase reporter assay approach. In brief, we synthesized WT or mutant (MU) versions of the miR-296-3p complementary sequences in circ_0005044 or FOSL1 and we inserted these constructs into the pmirGLO luciferase vector (Promega). We then conducted a reporter assay by cotransfecting 293T cells with WT or MU forms of these reporters together with miR-296-3p mimic or control constructs using Lipofectamine 2000 as mentioned previously. After 48 h, a Dual-Luciferase Reporter Assay System (Promega) was used to quantify luciferase activity in triplicate samples based on provided directions.
In vivo bone formation assay
To assess ectopic in vivo osteo/odontogenic bone formation, we mixed together 2 × 106 DPSCs that been transfected using si-circRNA or si-NC constructs together with 40 mg of deproteinized bovine bone tissue (Bio-Oss, Wolhusen, Switzerland). These mixtures were then implanted subcutaneously on the backs of nude mice (6 weeks old). At 8 weeks postimplantation, these samples were collected, fixed, and subjected to demineralization and paraffin embedding, and were cut into 2-μm-thick sections that were subjected to hematoxylin and eosin staining. In addition, RUNX2 expression was assessed by immunohistochemical staining. The Ethics Committee of Stomatological Hospital, Southern Medical University approved this study.
Statistical analysis
SPSS v16.0 (SPSS, Inc.) was used for all statistical testing. Differences between groups were compared via one-way ANOVAs and Student's t-tests as appropriate (*p < 0.05; **p < 0.01).
Results
Differential circRNA expression during DPSC osteo/odontogenic differentiation
We began by identifying circRNAs that were differentially expressed over the course of DPSC osteo/odontogenic differentiation. At 14 days postinduction of differentiation, we conducted the RNA-seq analysis of induced and control noninduced DPSC culture samples. When circRNA expression profiles were compared between these samples, we found that 87 circRNAs were differentially expressed as a function of differentiation status (log2FC ≥1, p < 0.05). Of these, 65 and 22 were up- and downregulated, respectively, in induced DPSCs (Fig. 1A).

RNA-seq results.
We then selected 5 of the identified differentially regulated circRNAs for qRT-PCR validation. We were able to confirm that the expression of these circRNAs changed significantly in response to osteo/odontogenic differentiation, consistent with our RNA-seq findings (Fig. 1B). We then leveraged previously published mRNA and miRNA profiles from DPSCs in the context of osteogenesis (Liu et al, 2020) to conduct a correlation analysis of differentially expressed circRNAs, miRNAs, and mRNAs. Following this analysis, we constructed a coexpression network (Fig. 1C). This network suggests that circ_0005044 has the potential to serve as a ceRNA for miR-296-3p, sequestering it and thereby preventing it from suppressing the expression of its target gene FOSL1. This suggested that the circ_0005044/miR-296-3p/FOSL1 axis may be involved in the osteogenic differentiation of DPSCs.
DPS osteoblastic differentiation is associated with circ_0005044 and FOSL1 upregulation and miR-296-3p downregulation
We next sought to confirm that miR-296-3p and FOSL1 expression were altered during DPSC osteo/odontogenic differentiation as predicted in the above bioinformatic analyses. To that end, we performed a qRT-PCR analysis of DPSCs following culture in OM. These cells exhibited significant upregulation of the osteogenic markers ALP, DSPP, and RUNX2 (Fig. 2A–C), indicating successful osteo/odontogenic induction of DPSCs. We found that circ_0005044 and FOSL1 expression rose over the course of this differentiation process, reaching maximum levels that were approximately six-fold higher than baseline after 14 days. In contrast, miR-296-3p expression fell significantly over this same time period in a manner negatively correlated with the expression of circ_0005044 and FOSL1 (Fig. 2D–F).
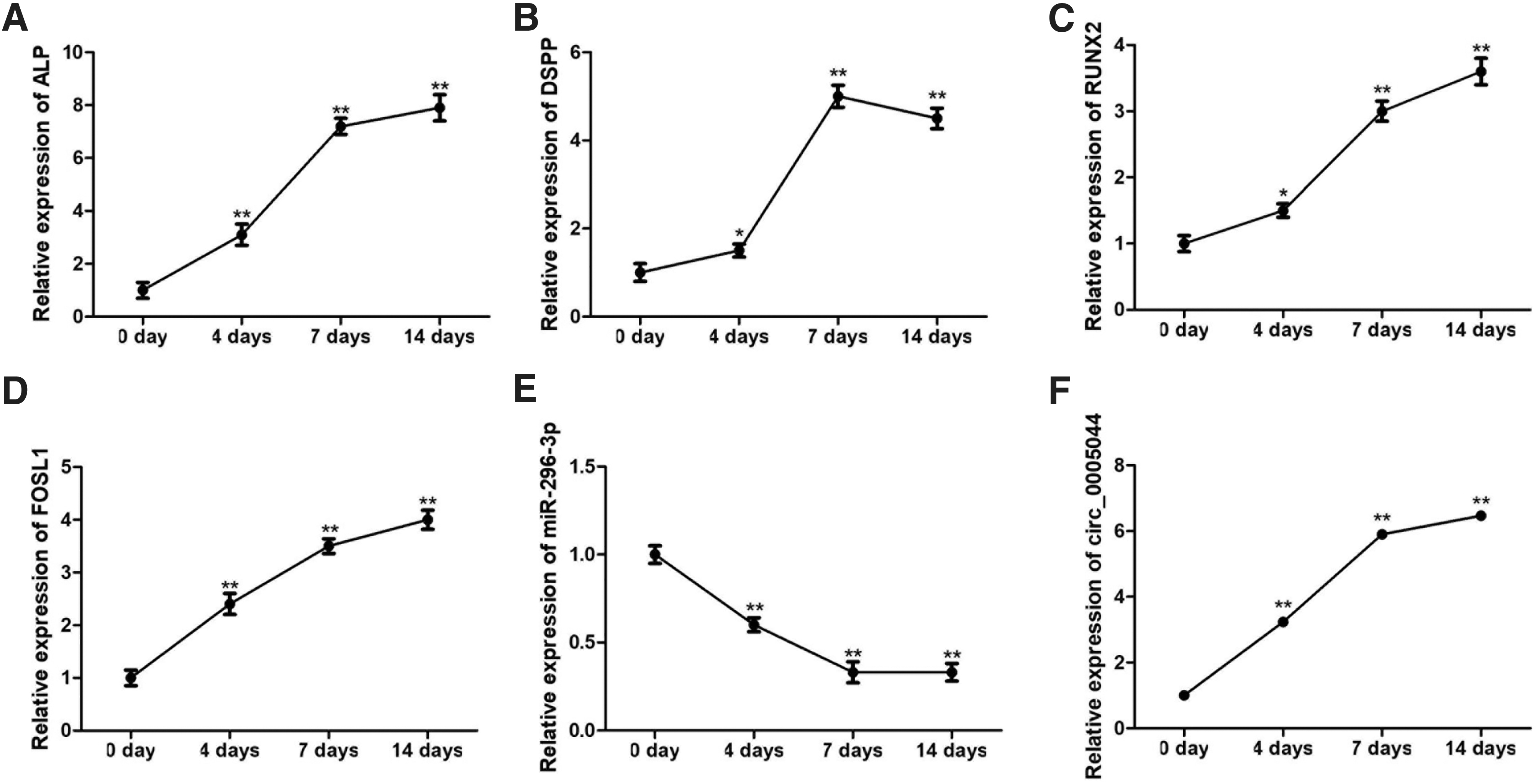
The expression of circ_0005044, miR-296-3p, and FOSL1 during DPSC osteogenic differentiation.
DPSC osteogenic differentiation is inhibited by the knockdown of circ_0005044
We next explored the biological impact of siRNA-mediated circ_0005044 knockdown after confirming the success of this approach via qRT-PCR (Fig. 3C). At 7 days postosteogenic induction, ALP staining was reduced in circ_0005044 knockdown cells (Fig. 3A), with a similar reduction in ARS staining after 14 days consistent with reduced matrix mineralization (Fig. 3B). We further confirmed via Western blotting that circ_0005044 knockdown resulted in reduced RUNX2 expression in DPSCs (Fig. 3D), with similar knockdown-associated reductions in the mRNA level expression of ALP, DSPP, and RUNX2 as well as a concomitant increase in levels of miR-296-3p (Fig. 3C). We confirmed using the RegRNA 2.0 database that circ_0005044 and miR-296-3p share a complementary sequence region. We then conducted a luciferase reporter assay, which revealed that miR-296-3p was able to interact with the WT but not MU version of this circ_0005044 sequence, consistent with the ability of these two nucleic acids to interact with one another within cells (Fig. 3E, F).
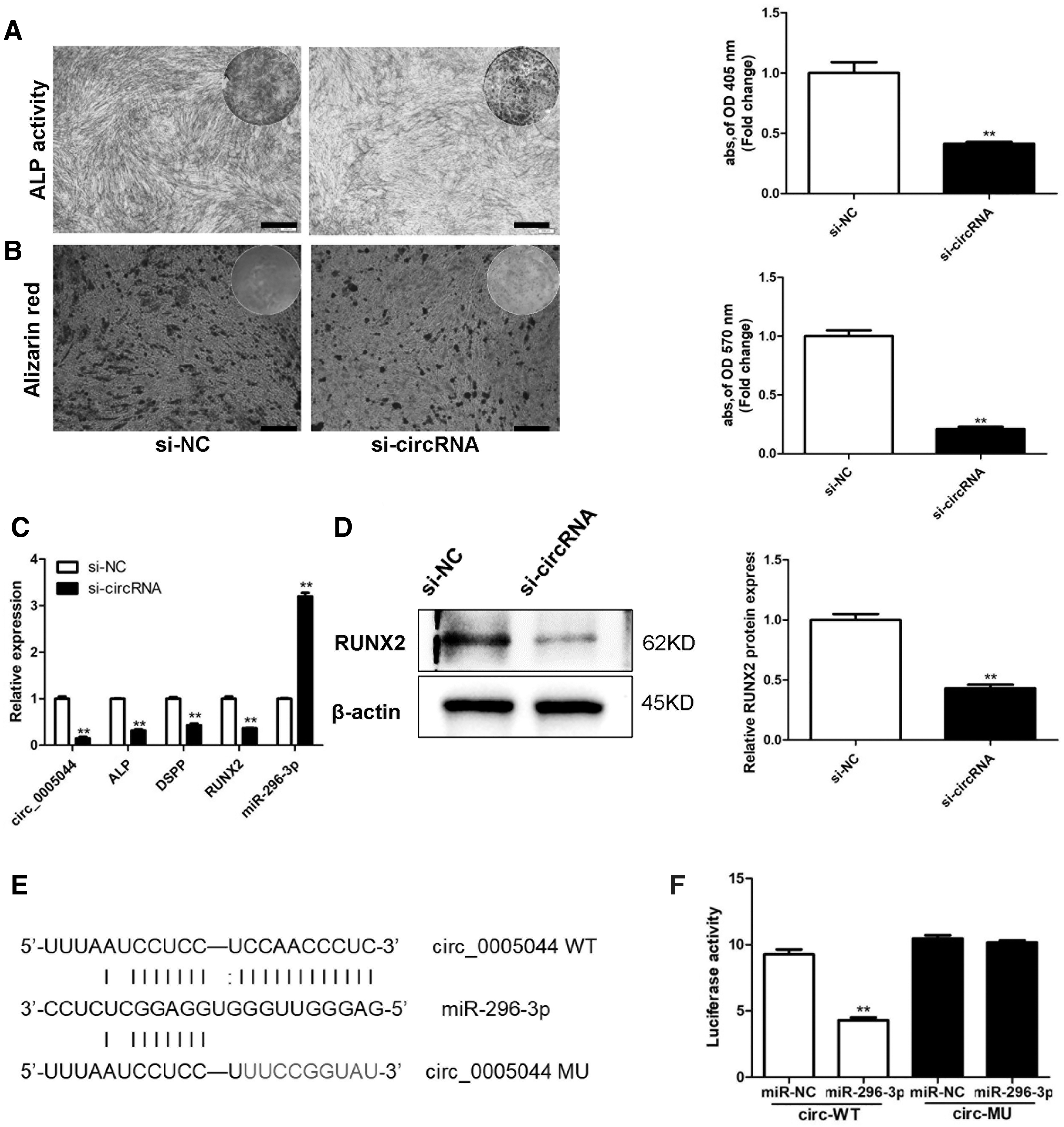
Circ_0005044 sequesters miR-296-3p and thereby regulates DPSC osteogenic differentiation.
miR-296-3p impairs DPSC osteo/odontogenic differentiation
We next sought to evaluate the role played by miR-296-3p in the context of DPSC osteo/odontogenic differentiation by knocking down or overexpressing this miRNA in these cells. Our overexpression approach led to a roughly four-fold increase in the expression of miR-296-3p in transfected cells, whereas knockdown efficiency was confirmed to be ∼70%. When this miRNA was overexpressed we observed reduced ALP staining, whereas the opposite phenotype was detected when it was instead knocked down (Fig. 4A). Comparable trends with respect to ARS staining were also observed at 14 days postosteogenic differentiation in treated cells (Fig. 4B). At the mRNA level, we found that miR-296-3p mimic transfection markedly reduced the expression of FOSL1, ALP, DSPP, and RUNX2, whereas miR-296-3p had the opposite impact (Fig. 4C). This was also consistent with protein level changes in FOSL1 and RUNX2 expression as assessed by Western blotting (Fig. 4D).

miR-296-3p inhibits DPSC osteogenic differentiation.
circ_0005044/miR-296-3p axis regulates FOSL1 expression
Consistent with our above predictive analyses and measurements, we used the MIRANDA database to confirm that miR-296-3p shares sequence complementarity with a portion of the 3′-untranslated region (UTR) of FOSL1 (Fig. 5A). Using a luciferase reporter assay system as mentioned previously, we then verified that miR-296-3p was able to directly bind to the WT but not the MU version of this 3′-UTR sequence, thereby confirming that this miRNA is capable of targeting this gene within cells so as to suppress its expression (Fig. 5B).
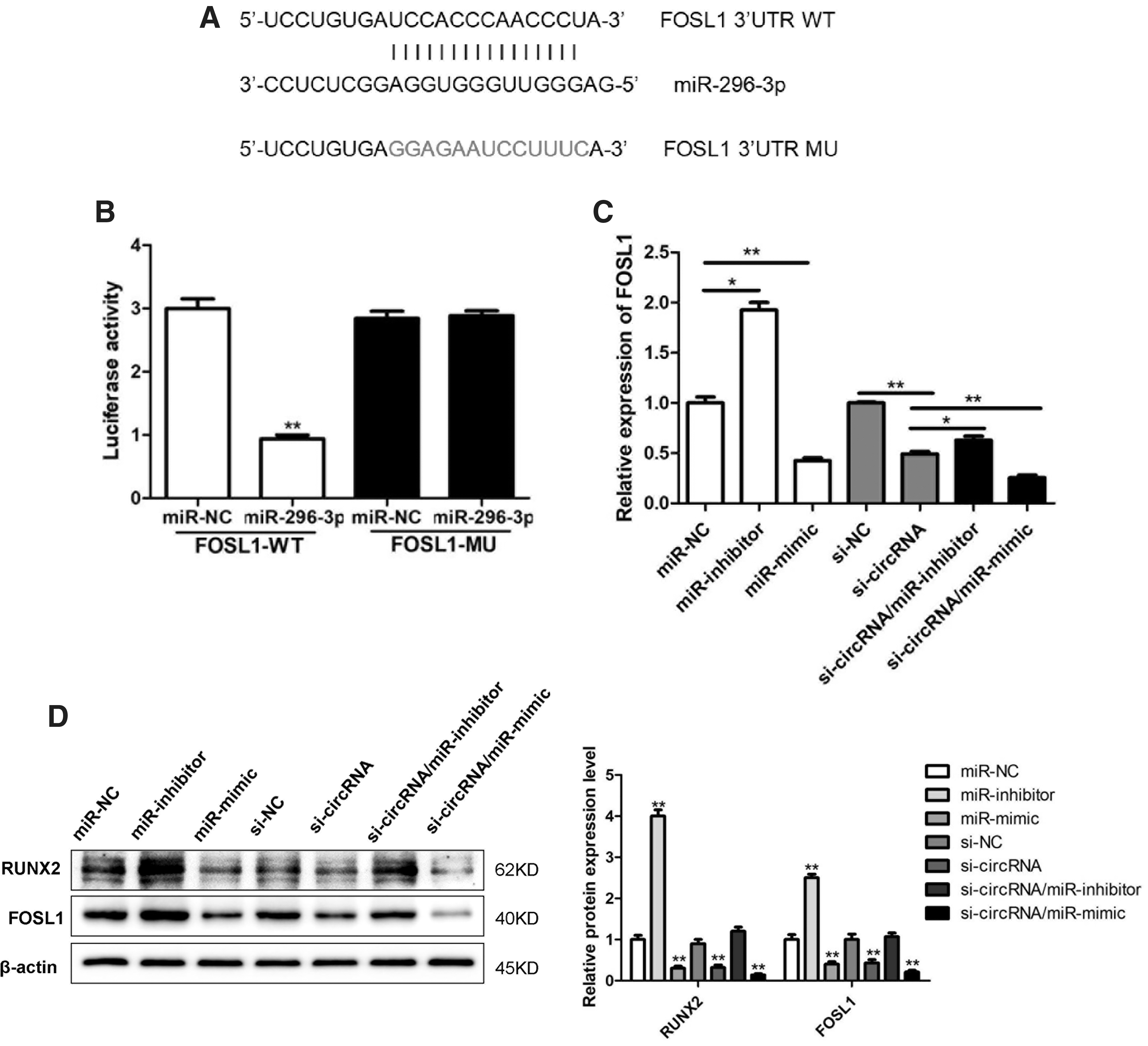
The circ_0005044/miR-296-3p axis controls FOSL1 expression.
To further establish FOSL1 as a target downstream of the circ_0005044/miR-296-3p axis in DPSCs, we next assessed the expression of FOSL1 at the mRNA and protein level in DPSCs that had been transfected with an miR-296-3p mimic, an miR-296-3p inhibitor, an siRNA specific for circ_0005044, or appropriate control constructs. We observed significant reductions in FOSL1 expression in response to circ_0005044 knockdown or miR-296-3p mimic transfection, whereas it was significantly upregulated in response to miR-296-3p inhibitor transfection (Fig. 5C, D). We additionally cotransfected cells with both si-circ_0005044 and miR-296-3p mimics or inhibitors, and we found that compared with cells transfected with only si-circ_0005044, FOSL1 levels were markedly reduced in cells cotransfected with si-circ_0005044 and a miR-296-3p mimic, whereas they were increased in cells cotransfected with si-circ_0005044 and miR-296-3p inhibitor (Fig. 5C, D). Together these findings confirm that circ_0005044 functions upstream of miR-296-3p, thereby interfering with the ability of this miRNA to suppress FOSL1 expression in DPTs during osteogenesis.
FOSL1 knockdown suppresses DPSC osteo/odontogenic differentiation and limits the ability of circ_0005044 to promote this process
To confirm that FOSL1 plays a role in DPSC osteo/odontogenic differentiation, we used an siRNA to knock it down with 70–80% efficiency in these cells. At appropriate time points, we observed significantly impaired ALP and ARS staining in cells in which FOSL1 had been knocked down (Fig. 6A, B). In line with this result, we detected significant decreases in the upregulation of ALP, DSPP, and RUNX2 in these FOSL1 knockdown cells (Fig. 6C). This similarly coincided with protein level reductions in RUNX2 and FOSL1 (Fig. 6D). To verify the relevance of circ_0005044/miR-296-3p-mediated FOSL1 regulation during DPSC osteogenesis, we next cotransfected these cells with both an miR-296-3p inhibitor and an FOSL1-specific siRNA before osteogenic induction. Cells that had also been transfected with the only miR-296-3p inhibitor exhibited increased ALP staining, whereas such staining was significantly reduced if cells had been cotransfected with the FOSL1-specific siRNA. Consistent with this, FOSL1 siRNA cotransfection partially disrupted matrix mineralization in miR-296-3p inhibitor-transfected DPSCs (Fig. 6E–I).

FOSL1 knockdown impaired DPSC osteogenesis and overcomes the impact of miR-296-3p inhibition.
Osteogenesis is inhibited by circ_0005044 knockdown in vivo
Finally, we sought to confirm that circ_0005044 plays a key role in DPSC osteoblastic differentiation in vivo but using a murine model system. After an 8-week period, we found that bone formation was significantly reduced in samples in which circ_0005044 had been knocked down relative to control samples (Fig. 7A). Consistent with this, the RUNX2 staining intensity in circ_0005044 knockdown samples was markedly reduced relative to that in control samples (Fig. 7B).

The role of circ_0005044 in the context of in vivo DPSC differentiation.
Discussion
Several studies to date have provided strong evidence that circRNAs can both regulate gene expression and serve as ideal biomarkers for the diagnosis of particular disease states (Hansen et al, 2013; Li et al, 2015b; Lukiw, 2013). For example, one recent analysis determined that both circ19142 and circ5846 were closely linked to the differentiation of osteoblasts (Qian et al, 2017), whereas BMP2 has been suggested to modulate this same osteogenic process via a circ19142/circ5846-targeted miRNA–mRNA axis. Another analysis aimed at clarifying circRNA expression profiles in periodontal ligament stem cells during osteogenesis revealed that these expression patterns varied in a defined manner over the course of differentiation and subsequent biomineralization, suggesting that these circRNAs play defined roles during this process (Zheng et al, 2017).
Few studies to date, however, have specifically examined the functional role of circRNAs during the osteo/odontogenic differentiation of DPSCs. In this study, we used an RNA-seq-based approach to identify 87 differentially expressed circRNAs (65 upregulated; 22 downregulated) associated with this differentiation process by comparing control cells with those that were actively differentiating. We further identified circ_0005044 as a highlight expressed upregulated circRNA that functionally regulated the DPSC osteo/odontogenic differentiation process. This was confirmed by the fact that circ_0005044 knockdown interfered with ALP and ARS staining, consistent with a disruption in normal DPSC osteoblastic differentiation in vitro. This was further validated through mRNA and protein level analyses of key osteogenesis-related genes. We additionally utilized an in vivo model system to confirm that circ_0005044 regulates osteogenesis in a more physiological environment. Overall, our findings highlight circ_0005044 as a possible regulator of DPSC osteo/odontogenic differentiation.
Within cells, circRNAs can function as ceRNAs capable of binding and sequestering specifying target miRNAs so as to modulate gene expression (Salmena et al, 2011). This functionality has led some authors to refer to circRNAs as miRNA “sponges,” given that they often contain multiple miRNA binding sites and can thus broadly regulate mRNA transcription and protein translation (Li et al, 2015a). In line with such functionality, we were able to demonstrate a strong correlation between circ_0005044 miR-296-3p, and we found that miR-296-3p overexpression led to a marked reduction in the expression of key osteo/odontogenic marker gene expression in DPSCs, whereas inhibition of this miRNA had the opposite effect. Together these results further support a model wherein circ_0005044 promotes DPSC osteo/odontogenic differentiation via the negative regulation of miR-296-3p.
Previous work has also highlighted roles for miR-296-3p in tumor metastasis in a number of different cancer types (Bai et al, 2013; Fu et al, 2017; Liu et al, 2013; Wang et al, 2018a). The specific role of this miRNA appears to be tumor specific, given that it has been shown to favor prostate cancer invasion and metastasis (Liu et al, 2013), but to suppress this same metastatic activity in the case of lung adenocarcinoma (Schodel et al, 2016). The role of this miRNA in osteogenesis has not previously been explored, and as such further research and validation of our results are warranted.
Through further work, we determined that circ_0005044/miR-296-3p regulate the osteo/odontogenic process at least in part via controlling FOSL1 expression levels within DPSCs. This was supported by complementary miR-296-3p binding sites within the FOSL1 3′-UTR, and a direct binding interaction was confirmed through the use of a luciferase reporter assay system. Of importance, we found that overexpressing miR-296-3p and knocking down circ_0005044 both reduced FOSL1 protein levels within DPSCs, whereas these levels were increased when miR-296-3p was knocked down. When cells were cotransfected using both a miR-296-3p mimic and an siRNA specific for circ_0005044, this led to a more profound reduction in FOSL1 levels relative to siRNA transfection alone, whereas cotransfecting cells with both a miR-296-3p inhibitor and si-circ_0005044 partially disrupted the intracellular activity of circ_0005044 knockdown. Together these findings provided strong evidence that circ_0005044 functions upstream of miR-296-3p and thereby indirectly modulates the levels of FOSL1 expressed within cells.
FOSL1 (also known as Fra-1) is an AP-1 family transcription factor protein that is known to be a key regulator of osteogenic processes (Jochum et al, 2000; Yu et al, 2013). In murine model systems, lack of c-Fos expression can result in the development of osteoporosis (Wang et al, 1992), but this phenotype can be reversed when FOSL1 is overexpressed (Fleischmann et al, 2000), as this gene functions downstream of c-Fos in osteoclast progenitor cells (Matsuo et al, 2000). Mice that overexpress FOSL1 constitutively have been shown to suffer from progressive osteosclerosis (Jochum et al, 2000) and severe lipodystrophy (Luther et al, 2011), whereas knockout of this gene in embryonic mice is associated with an osteopenic phenotype (Eferl et al, 2004).
In our analysis, the knockdown of FOSL1 significantly impaired DPSC osteo/odontogenic differentiation in a manner consistent with the phenotypes observed when cells were transfected using either a circ_0005044 siRNA or an miR-296-3p mimic. When we knocked down FOSL1, this partially overcame the ability of miR-296-3p inhibition to promote DPSC osteogenesis. These findings therefore further supported a model wherein the circ_0005044/miR-296-3p pathway regulates DPSC differentiation through FOSL1.
When FOSL1 was inhibited, we observed mRNA and protein level reductions in the expression of RUNX2. The mechanistic basis for this finding, however, remains to be clarified. Prior work, however, has shown that FOSL1 upregulation can lead to changes in PI3K/AKT pathway activation in gastric cancer cells (He et al, 2015). This PI3K/AKT pathway has also been associated with RUNX2 expression levels in osteoblasts (Chen et al, 2019). As such, changes in FOSL1 expression within differentiating DPSCs may lead to changes in RUNX2 expression levels via a mechanism associated with PI3K/AKT signaling. Further studies will be essential to test this hypothesis.
Osteo/odontogenic differentiation of DPSCs is very important in dentin–pulp complex repair process. Our results suggest that hsa_circ_0005044 might be a novel biomarker or therapeutic target for regulating dentin–pulp complex repair. And one challenge is that the limited resources of DPSCs in permanent teeth will retard the application in the future. To solve the question, the DPSCs may be produced with the capabilities of immortalization or increased the number of cell passages. And we believe that DPSCs will be used not only for research but also for the clinical application in the future.
Conclusions
In summary, in this study we provide novel evidence that circ_0005044 is associated with DPSC osteo/odontogenic differentiation. We found that circ_0005044 is capable of regulating miR-296-3p, leading to a consequent increase in the expression of FOSL1. As such, circ_0005044 may be a viable biomarker or therapeutic target in the context of efforts aimed at modulating the osteoblastic differentiation of DPSCs.
Footnotes
Data Availability Statement
The RNA-seq data is openly available in NCBI Sequence Read Archive (
Acknowledgment
Zhongjun Liu and Siwei Li contributed equally to this study.
Disclosure Statement
No competing financial interests exist.
Funding Information
This study was supported by grants from the National Natural Science Foundation of China (81800957), Natural Science Foundation of Xinjiang Uygur Autonomous region (2019D01C00), Guangzhou Municipal Science and Technology Project (201904010068, 202201011515) and Guangdong Medical Research Fund (A2019567, A2017572, B2018065 and B2018121).
Supplementary Material
Supplementary Figure S1
References
Supplementary Material
Please find the following supplemental material available below.
For Open Access articles published under a Creative Commons License, all supplemental material carries the same license as the article it is associated with.
For non-Open Access articles published, all supplemental material carries a non-exclusive license, and permission requests for re-use of supplemental material or any part of supplemental material shall be sent directly to the copyright owner as specified in the copyright notice associated with the article.