
Abstract
Circular RNA (circRNA) is involved in the occurrence and development of various cancers. To this day, the expression and mechanism of circRNA in osteosarcoma (OS) remain unclear. We previously found that circ_0001060 was highly expressed in OS tumor tissues. In this work, we identified that high level expression of circ_0001060 was significantly associated with late clinical stage, larger tumor volume, higher frequency of metastasis, and poor prognosis in OS patients. Furthermore, we confirmed that silencing circ_0001060 inhibited the proliferation and migration of OS cell. Using bioinformatics analysis, we built three circRNA-miRNA-mRNA regulatory modules (circ_0001060-miR-203a-5p-TRIM21, circ_0001060-miR-208b-5p-MAP3K5, and circ_0001060-miR-203a-5p-PRKX), suggesting that these signaling axes may be involved in the inhibitory effect of circ_0001060 on OS. To sum up, circ_0001060 is a novel tumor biomarker for OS as well as a potential therapeutic target.
Introduction
Osteosarcoma (OS) is the most common primary bone malignancy that arises from transformed mesenchymal cells (Whelan and Davis, 2018). OS is an aggressive cancer that has rapid progression, high rate of relapse, and great lung metastatic potential, which make it one of the most common accounts of death caused by cancer in youngsters and young adults (Noone et al, 2017). After a standard treatment procedure with neoadjuvant chemotherapy, the 5-year survival rate of patients with primary OS is ∼65–70%, but if metastasis or recurrence occurs, this value will sharply drop to less than 20% (Gorlick et al, 2013; Isakoff et al, 2015). Over the past three decades, therapeutic research dedicated to improving the survival rate of OS patients has sustained (such as immunotherapy), even so the outcomes are barely satisfactory (Gill and Gorlick, 2021; Meltzer and Helman, 2021). Thus, it is vital to search for novel druggable targets for OS.
Circular RNAs (circRNAs), which manufactured by back-splicing of exons, are covalently closed, endogenous RNA splicing product with tissue-restricted and cell-specific expression patterns (Kristensen et al, 2019). Acting as noncoding RNAs, circRNAs can regulate protein function by serving as sponges for RNA binding proteins and microRNAs or conduct self-translation to exert biological functions (Chen, 2020). Getting benefit from the technological surmounts in high-throughput sequencing and bioinformatics algorithms, related studies of circRNAs have been greatly motivated (Beermann et al, 2016).
After nearly a decade of preliminary research, circRNAs have become the focus of noncoding RNA research and assume a key role in the process of cancer development through multifarious mechanisms (Kristensen et al, 2022; Liu et al, 2022; Ma et al, 2020; Yu et al, 2022). CircRNAs has potential proven clinical utility, and its structural stability, evolutionary conservation, organ specificity, and detection convenience (which can be detected in liquid samples such as plasma, saliva, and urine) also make it an ideal potential diagnostic and prognostic biomarker (Cui et al, 2018; Meng et al, 2017; Tu et al, 2020). However, the specific mechanism of circRNAs in OS has yet to be elucidated.
Our previous research obtained the differentially expressed circRNAs in tumor tissues and adjacent tissues of OS patients via RNA sequencing technology (Xi et al, 2019). A new circRNA named circ_20403 was found (also known as circBase ID: circ_0001060, Alias: circ_001971), which is highly expressed in OS tumor tissues and may be participated in regulating the occurrence and development of OS. In this research, we find that high level expression of circ_0001060 is significantly associated with late clinical stage, larger tumor volume, higher frequency of metastasis, and poor prognosis in OS patients. In addition, our experimental results confirm that silencing circ_0001060 can inhibit the proliferation and migration of OS cell line. We conclude that circ_0001060 is a novel tumor biomarker for OS as well as potential therapeutic target.
Materials and Methods
Clinical data
Clinical follow-up data were gathered from the archives of the First Affiliated Hospital of Guangxi Medical University. Ethical approval was acquired from the ethics committees of the First Affiliated Hospital of Guangxi Medical University (approval no. 2019KY-E-097). All procedures involving human participants were conducted in accordance with the Declaration of Helsinki. All subjects and their parents signed informed consent for the whole procedures.
Cell culture
Human OS cells, the Saos-2 (China Center for Type Culture Collection [CCTCC]: GDC0075), 143B (CCTCC: GDC0076), MG-63 (CCTCC: GDC0074), HOS (CCTCC: GDC0333), and U2OS (CCTCC: GDC0626), were purchased from CCTCC (Wuhan, China). And, human osteoblast hFOB1.19 (National Collection of Authenticated Cell Cultures [NCACC]: GNHu14) was purchased from the NCACC (Shanghai, China). Human OS cell lines were cultured in Dulbecco's modified Eagle's medium (DMEM) supplemented with 10% fetal bovine serum (Gibco, USA), 100 U/mL penicillin, and 100 U/mL streptomycin (Invitrogen, USA). hFOB1.19 cell line was cultured in DMEM/Nutrient Mixture F-12 supplemented with 10% fetal bovine serum (Gibco) and 0.6 mg/mL G418 (Beyotime, China). All human OS cells were cultured in a 37°C cell incubator with 5% CO2. And, hFOB1.19 cells were cultured in a 34°C incubator with 5% CO2.
Quantitative reverse transcription-PCR assay
Total RNA obtention from human OS cells and hFOB1.19 cells was performed with TRIzol (Invitrogen) reagent and circRNA and mRNA reverse transcribed with a PrimeScript™ RT reagent kit (Takara, Japan), under a guidance of the manufacturer's protocols. And, the reactions were conducted via PowerTrack™ SYBR Green Master Mix (Thermo Fisher Scientific, USA) and subsequently measured by a LightCycler® 96 PCR System (Roche, Switzerland) in the light of the manufacturer's guidances. The relative RNA expression levels were assessed by the 2−△△Ct algorithm, and GAPDH act as an internal reference. And, PCR products were performed with Sanger-sequenced (Sequetech, Mountain View, CA, USA).
Nucleic acid electrophoresis and RNase R treatment
Following RNA collection, cDNA was synthesized by making use of a PrimeScript RT reagent kit (Takara). Genomic DNA (gDNA) was extracted from Saos-2 and HOS cells using the QIAamp DNA kit (Qiagen, Germany). The PCR products of cDNA and gDNA were run on agarose gel electrophoresis with Tris Acetate-EDTA buffer. The gel was run initially for 30 min at 120 V. The DNA Marker (Sangon Biotech, China) was utilized to be DNA marker. The bands were identified on the gels by shadowing with UV irradiation. As for RNase R intervention, 2 μg of RNA were mixed with or without 3 U/μg RNase R (Sigma–Aldrich, USA) for10 min at 37°C.
Cell transfection
Human lentivirus-sh-circ0001060 was purchased from RiboBio (Guangzhou, China). Lentivirus-sh-circ0001060 was ultracentrifuged, concentrated, and put in the Saos-2 and HOS cells culture medium. Following virus infection, cells were screened with puromycin dihydrochloride (Beyotime) for 7 days, and viable cells next acted as stably transfected cells. And, cells were frozen at 80°C for later use.
Cell viability and proliferation assay
Cultivation mode of Saos-2 and HOS cells were conducted as described previously. After cultured for 1, 2, 3, 4, 5, 6, and 7 days, 10 μL of Cell Counting Kit-8 reagent (Beyotime) were put in each well with 2 h of incubation, and then, cell viability was measured at 450 nm by a microplate reader (Thermo Fisher Scientific). The percentage cell viability was estimated contrast with the average absorbance of transfected cells to that of untransfected cells. Cell proliferation level was detected by EdU staining using a BeyoClick™ EdU-488 Cell Proliferation Kit (Beyotime), in the light of the manufacturer's instructions.
Colony formation assay
Saos-2 and HOS cells were inoculated at 600 cells per well into 12-well culture plates and laid aside in a 37°C cell incubator for 8 days. Then, the cells were washed with phosphate buffered saline (PBS) three times, fixed with 4% paraformaldehyde for 30 min, and finally stained with 0.1% crystal violet solution for 20 min. The colonies were tallied and photographed under an invert microscope (Leica, Germany).
Wound healing assay
Saos-2 and HOS cells were seeded in 24-well culture plates (Corning, NY, USA). After reaching 80% density, cell monolayers were scored with a 10 μL sterile pipette tip. Then, each cell culture well was washed with PBS three times and was replaced FBS-free culture medium. At 0 and 48 h following scratching of the monolayers, cell migration in three random areas was viewed and images were captured using a digital camera (Leica). Quantification of the percentage of wound healing was calculated by the ImageJ software.
Migration and invasion assay
After digesting the Saos-2 and HOS cells, they were scoured twice with PBS and suspended with serum-free medium. Take 100 μL of cell suspension and add it to upper chamber of Transwell Permeable Supports (Corning) with (transwell invasion assay) and without (transwell migration assay) Matrigel (BD Biosciences, USA). Five hundred microliters of FBS medium were put in the lower chamber of 24-well plate and cultured for 12 h. Cells in lower side of upper chamber were fastened and then dyed with 0.1% crystal violet. Images were photographed by microscope (Leica), and cell counts were conducted using the ImageJ software.
Cell apoptosis and cell cycle analysis
7-Aminoactinomycin D cell viability assay kit (Beyotime) was applied to discern cell apoptosis analysis, and cell cycle analysis kit (Beyotime) was employed to ascertain cell cycle progression, following the manufacturer's instructions.
Fluorescence in situ hybridization
Fluorescence-labeled circ_0001060 probes were devised and synthesized by RiboBio. The probe image was observed with the Fluorescent In Situ Hybridization (FISH) Kit (RiboBio) following the manufacturer's instructions. Images were photographed via a confocal fluorescence microscope (Leica).
Nucleocytoplasmic separation and quantitative reverse transcription-PCR
The RNA in the nuclei and cytoplasm of Saos-2 and HOS cells was collected by using Cytoplasmic and Nuclear RNA Purification Kit (Norgen Biotek, Canada), following the manufacturer's protocols. Then, quantitative reverse transcription (qRT)-PCRs were carried out to determine the relative RNA expression levels.
Bioinformatics analysis
Candidate miRNAs for circ_0001060 were determined through the crossover of results from circBank databases (
Statistical analysis
Statistical analysis was performed using SPSS 17.0 software (IBM, USA) and GraphPad Prism version 8.0 (GraphPad Software, USA). The results in the graphs are shown as the mean ± SD of three independent experiments. Differences between groups were determined by the Student t-test. p < 0.05 was considered statistically significant. Subject operating characteristic curves were created to assess its diagnostic value.
Results
High circ_0001060 expression level indicates poor prognosis in OS
Our previous research has verified that circ_0001060 is highly expressed in OS tumor tissues (Xi et al, 2019). We next trace the clinical follow-up of the subjects involved, and baseline and follow-up clinical characteristics are listed in Table 1 (see details in Supplementary Table S1). After data collection and analysis, we observed a strong positive correlation between the high expression of circ_0001060 with distant metastasis (p = 0.0490; Fig. 1a) and larger tumor size (p = 0.0149; Fig. 1b). Other factors, including age (p = 0.9064), gender (p = 0.0613), and primary tumor location (p = 0.8403; Fig. 1c), were not found to be correlated with the expression of circ_0001060.
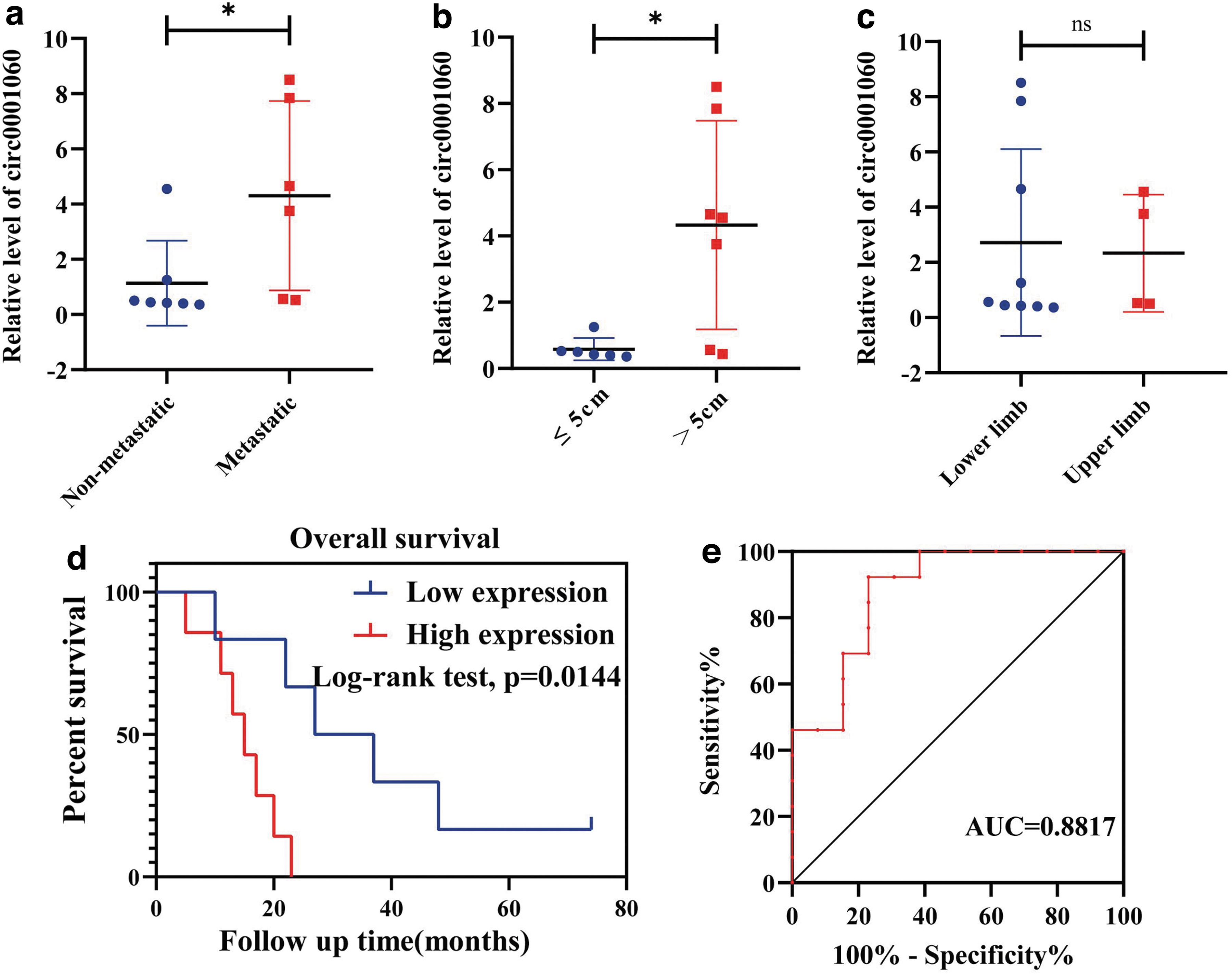
Correlation of circ_0001060 expression with clinical characteristics. Correlation between circ0001060 expression and
Correlations Between circ_0001060 Expression and Follow-Up Clinical Characteristics of Patients with Osteosarcoma
p < 0.05.
The median overall survival between the low and high circ_0001060 expression groups (in accordance with the mean value of circ_0001060 expression level) was appraised by the Kaplan–Meier analysis. And, we found that patients with higher expression level of circ_0001060 demonstrated poorer overall survival rates. In addition, receiver operating characteristic (ROC) analysis was performed for ascertaining the diagnostic value of circ_0001060 based on 13 pairs of tissue samples. As shown in Figure 1e, the area under the ROC curve is 0.8817, which suggests that circ_0001060 is a potential OS diagnostic marker. Thus, we speculate that high circ_0001060 expression is tightly linked to unfavorable prognosis and may be a potential target in OS.
Characterization of circ_0001060
Before functional biological exploration of circ_0001060 in OS, we first validate the circRNA structure and cyclization site. By querying the circBase database (
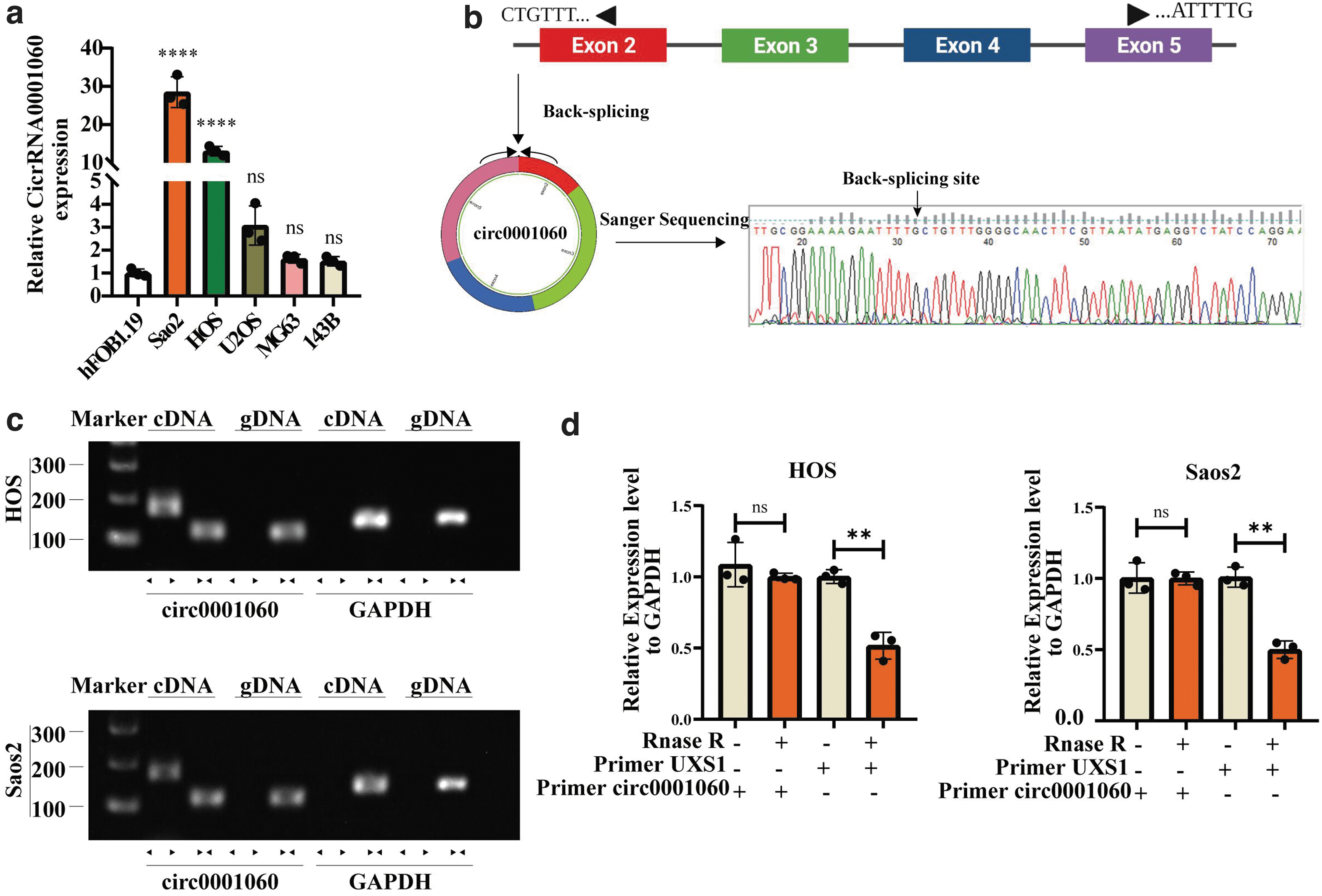
Characterization of circ_0001060 in OS.
We used divergent primer and convergent primer to amplify circ_0001060 and GAPDH (as a negative control) in cDNA and gDNA, respectively. Divergent primers amplified circ_0001060 in cDNA, but not in gDNA, indicating that circ_0001060 is reverse looping (Memczak et al, 2013) (Fig. 2c). CircRNAs possess a circular stable structure, which makes it stabilize under the degradation of RNase enzyme (Chen, 2020). Herein, we performed qRT-PCR after RNase R digestion. The relative expression of linear GAPDH and USX1 in RNase R group decreased significantly, whereas circ_0001060 decreased only slightly, indicating that circ_0001060 is more tolerant to digestion than linear RNA. So far, we have preliminarily determined that circ_0001060 is the further research target.
Tumor suppressor effect of circ_0001060 knockdown in OS cell lines in vitro
For stable circ_0001060 knockdown in OS cell lines, we designed three small hairpin RNAs (shRNAs) with silent sites to specifically target the junction sites of circ_0001060. After transfected shcirc_0001060 into Saos2 and HOS cells, we found that circ_0001060 expression was significantly silenced by shRNA via performing qRT-PCR, and mostly silenced by shcirc_0001060 RNAi 01 (Fig. 3a). Thus, RNAi 01 was selected for further research.
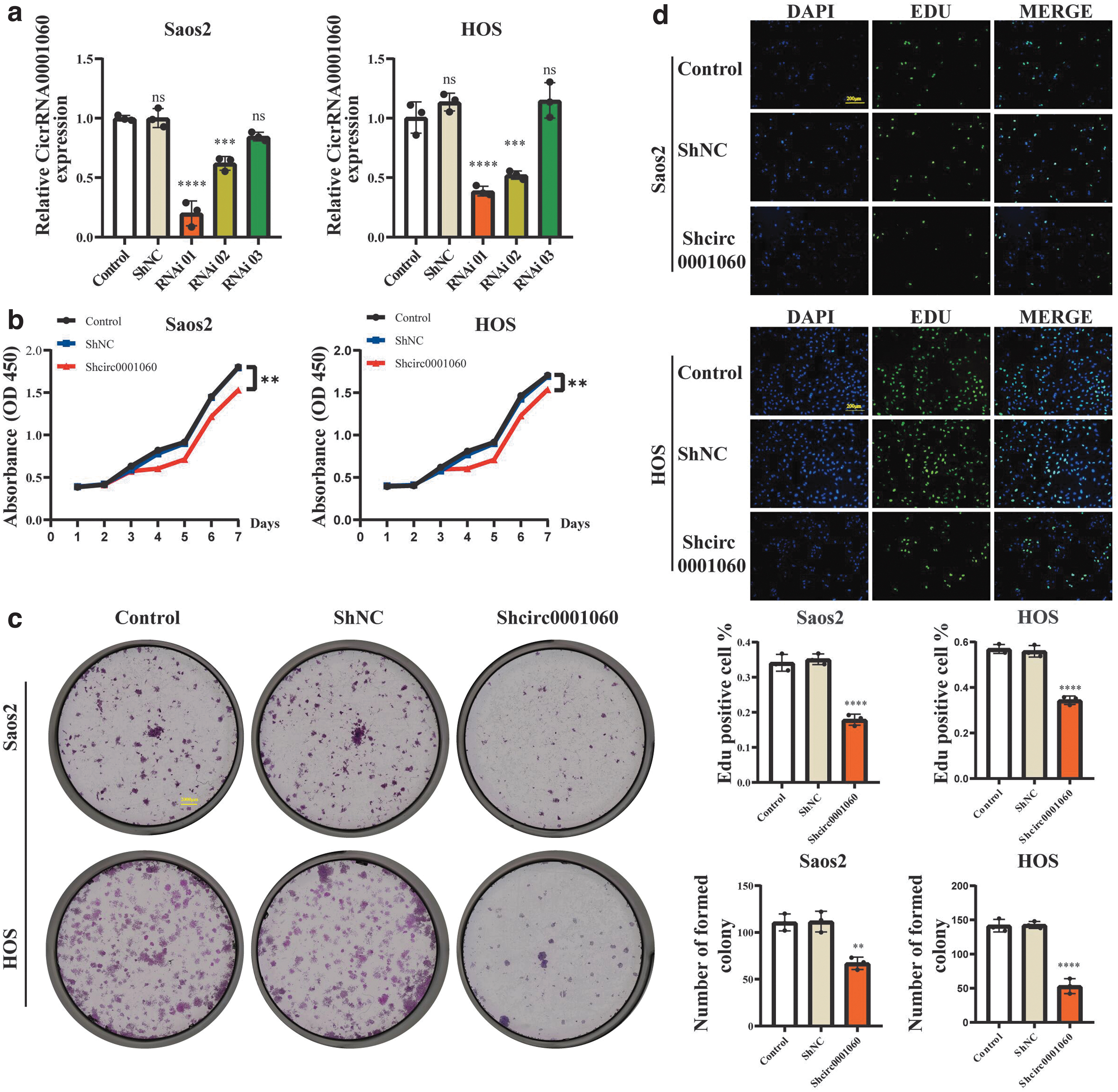
Silencing of circ_0001060 inhibits the proliferation of OS cell lines in vitro.
We observed that proliferation and colony-forming abilities of Saos2 and HOS cells were considerably suppressed after silencing circ_0001060 (Fig. 3b–d). Meanwhile, wound healing assay, migration assay, and invasion assay further revealed that circ_0001060 knockdown inhibited migration and invasion capacities of Saos2 and HOS cells (Fig. 4a, b). Moreover, we found that circ_0001060 knockdown greatly increased the number of apoptotic Saos2 and HOS cells (Fig. 5a) and induced G0/G1 phase arrest (Fig. 5b). Overall, our research results indicated that silencing circ_0001060 can be a “stumbling block” in OS development.
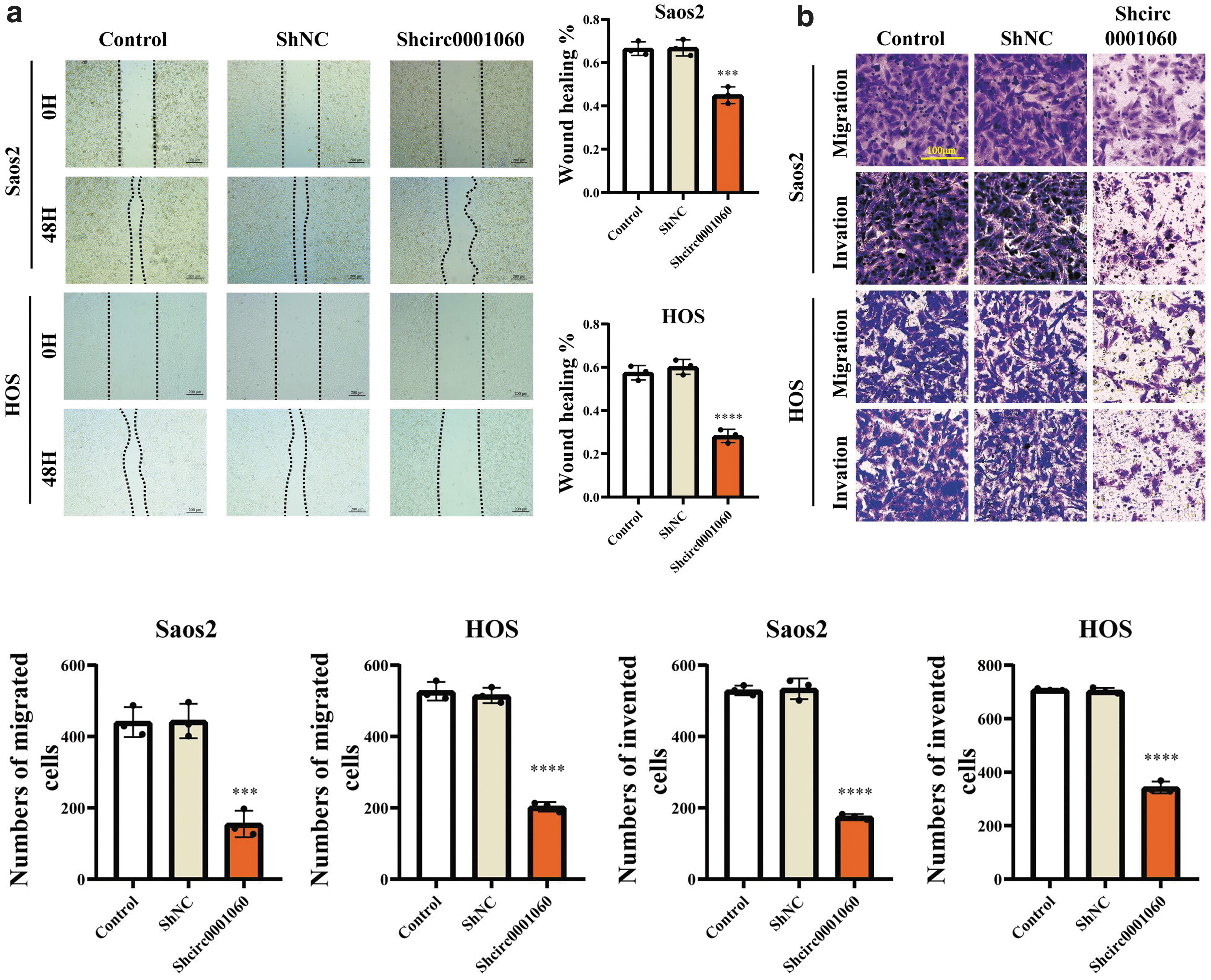
Silencing of circ_0001060 inhibits the migration and invasion of OS cell lines in vitro. Cell migration and invasion abilities of HOS and Saos2 cells transfected with shcirc0001060 or shNC were evaluated by

Silencing of circ_0001060 promotes the apoptosis and inhibits the cell cycle distribution of OS cell lines in vitro. Flow cytometry was used to evaluate the effect of circ0001060 knockdown on the
Cytoplasm localization, downstream target miRNAs, and function prediction of circ_0001060
By acting as a competing endogenous RNAs, circRNAs can compete for miRNAs binding as a miRNA sponge and exist extensively in the cytoplasm (Fontemaggi et al, 2021; Zhong et al, 2018). By performing FISH assay, we observed that circ_0001060 was localized mainly in the cytoplasm under confocal microscope (Fig. 6a). Meanwhile, the result of qRT-PCR after nucleocytoplasmic fractionation intervenes also revealed the similar phenomenon (Fig. 6b). Based on these grounds, we conjectured that circ_0001060 may play a role of miRNA sponge. Following this, we next identified nine candidate miRNAs by ascertaining the intersection of the results from two bioinformatics databases (RNAhybrid and circBank; Fig. 6c). And these nine miRNAs regulate 80 (hsa-miR-627-3p), 23 (hsa-miR-140-5p), 10 (hsa-miR-203a-5p), 25 (hsa-miR-208b-5p), 19 (hsa-miR-29a-5p), 21 (hsa-miR-4762-3p), 93 (hsa-miR-5582-3p), 15 (hsa-miR-6769a-5p), and 149 (hsa-miR-6873-3p) downstream target protein genes, respectively.

Downstream target miRNAs and function prediction of circ_0001060.
In this way, we build a network including circ_0001060, nine above miRNAs and their downstream targets to show their interactions (Fig. 6d). Furthermore, we assessed functional enrichment in the downstream target genes via GO and KEGG annotations. And, the results of our analysis exhibited that downstream target genes were enriched in varied biological processes and signaling pathways, such as the cytoplasm, regulation of transcription from RNA polymerase II promoter, protein binding processes, and pathways in cancer (Fig. 6e). All these revealed the circRNA signatures of circ_0001060 and indicated that it may participate in the tumor occurrence and development.
Potential circRNA-miRNA-mRNA network of circ_0001060 in OS
To find out the hub genes among the target genes, we constructed a PPI network by applying STRING (Szklarczyk et al, 2019) (Fig. 7a). We used the MCODE application in Cytoscape to seek out the vital module and identified 6 vital modules containing 26 hub genes (Fig. 7b). Next, we analyzed the overall survival to ascertain the prognostic value of these hub genes. Finally, we screened out three significantly hub genes, Tripartite Motif Containing 21 (TRIM21), Mitogen-Activated Protein Kinase Kinase Kinase 5 (MAP3K5), and Protein Kinase X-Linked (PRKX).
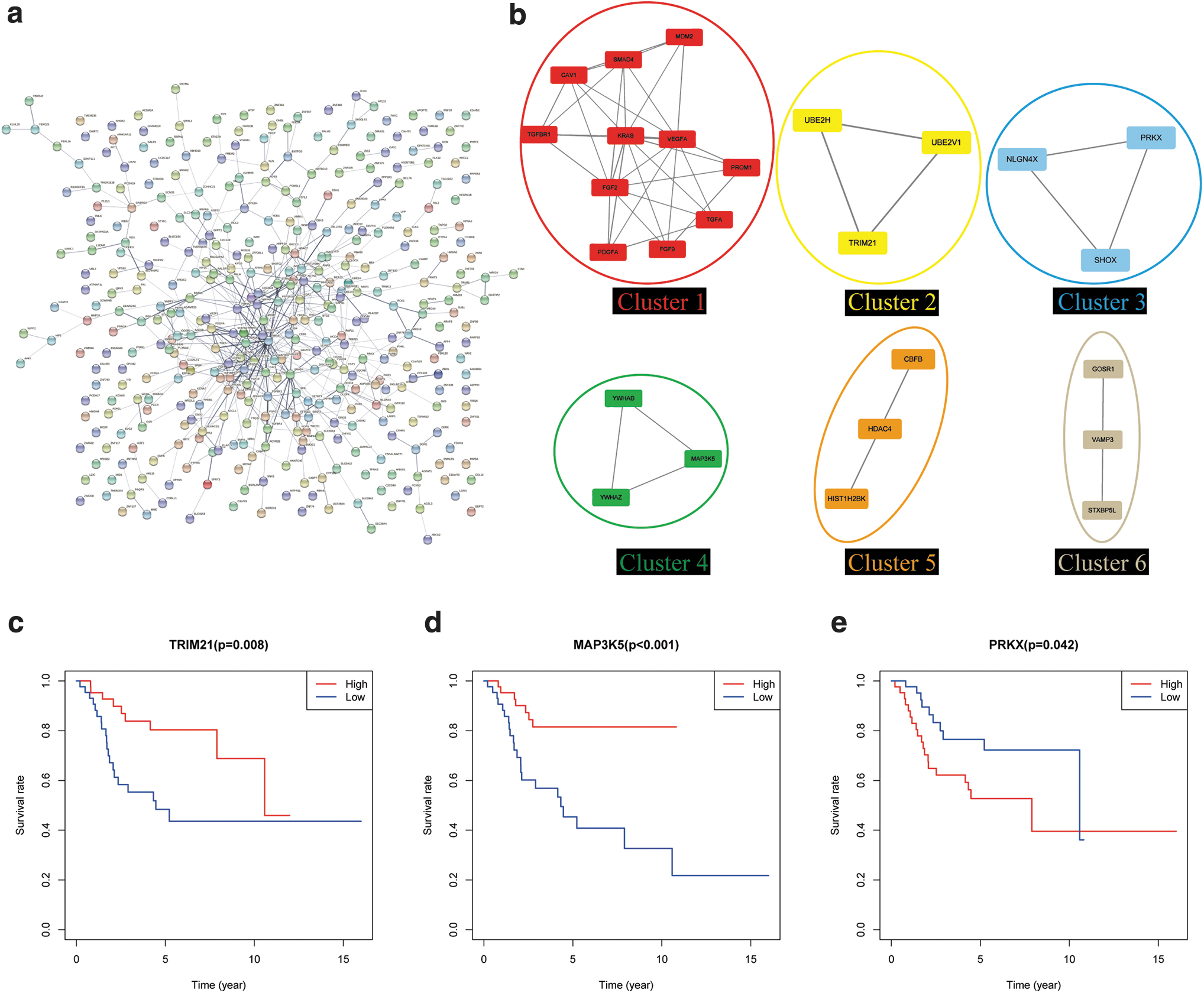
Key genes of circ_0001060 circRNA-miRNA-mRNA network in OS.
The survival curves of TRIM21, MAP3K5, and PRKX (Fig. 7c–e) showed that higher expression of TRIM21 and MAP3K5 was associated with increased overall survival (p = 0.008 and p < 0.001, respectively), and higher expression of PRKX was relevant to worse overall survival in OS patients (p = 0.042). In view of the above, we established three circRNA-miRNA-mRNA regulatory modules (circ_0001060-miR-203a-5p-TRIM21, circ_0001060-miR-208b-5p-MAP3K5, and circ_0001060-miR-203a-5p-PRKX). And these signaling axes may have potential effects on the tumorigenesis of OS.
Discussion
Nowadays, OS remains as one of the thorniest issues that must be solved owing to its highly metastatic and poor survival rate (Gorlick et al, 2013; Isakoff et al, 2015). What is more, a large proportion of patients with OS have already emerged subclinical pulmonary micrometastasis when diagnosed (Cortini et al, 2017). Moreover, the insufficiency of definite biomarkers has greatly slowed down the research progress of improving the clinical results of OS (Burns et al, 2020). These years, circRNAs research in carcinogenesis became a hot trend in the scientific community, for instance, in the progression and metastasis of lung adenocarcinoma (Qiu et al, 2018), gastric cancer (Ma et al, 2020), bladder cancer (Xie et al, 2018), and prostate cancer (Yu et al, 2022). The circRNAs related to OS have been involved in some previous studies (Liu et al, 2017, 2018; Wu et al, 2019). Yet, there remains blind zone in the biological function of circRNAs in OS. Herein, we reveal that circ_0001060 is a potential biomarker for OS and forecasted its downstream biological function.
In our preliminary work, we have screened circ_0001060, which shows high expression in OS by high-throughput sequencing (Xi et al, 2019). circRNA is a kind of circular RNA with stable structure, which can be divided into three categories: exonic circRNAs, exon–intron circRNAs, and intronic circRNAs (Kristensen et al, 2019; Memczak et al, 2013). Our Sanger sequencing result indicated that circ_0001060 is formed by back-splicing of the 3′ and 5′ ends of exons 2–5 of UXS1. UXS1 encodes an enzyme, pitched in the perinuclear Golgi, catalyzes the synthesis of UDP-xylose involved in glycosaminoglycan (GAG) synthesis on proteoglycans. The GAG chains are covalently linked to proteoglycans, which partake cancer signaling and signaling pathways (Gubbiotti et al, 2020; Wiweger et al, 2011). What interests us is that circ_0001060 (also named circ_001971) has a positive effect on colorectal cancer growth, metastasis, and angiogenesis as reported in the previous literature, which suggests the cancer-promoting function of circ_0001060 (Chen et al, 2020).
Our clinical data of 13 patients suggest that circ_0001060 expression is consistent with the positive correlation between distant metastasis and tumor size. More importantly, the Kaplan–Meier analysis and ROC analysis revealed a poorer survival role of circ_0001060 in OS patients, which indicated that circ_0001060 might have a hand in the development of OS. We next determined the stable cyclic structure of circ_0001060, an indispensable characteristic of circRNAs. We further revealed the promotional effect of circ_0001060 in OS proliferation, migration, and invasion. And, we also confirmed that circ_0001060 regulates the apoptosis and cell cycle of OS cells in vitro.
The subcellular localization of circRNAs is closely linked with its function execution (Piwecka et al, 2017). A growing number of evidence show that cytoplasmic circRNAs are miRNAs sponge, which can restrain translation or induce target mRNA degradation after binding to Ago2 protein (Chen, 2020; Liu and Chen, 2022). We found out that circ_0001060 is localized largely to cytosolic compartments of the OS cells via nucleocytoplasmic fractionation and FISH assay. Therefore, we speculated that circ_0001060 is poised for miRNAs sponge functionality. In this research, we obtained nine predicted target miRNAs of circ_0001060 by the bioinformatics databases. Next, bioinformatics analysis of miRNAs downstream targets demonstrated that circ_0001060 may be closely related to tumor and circRNAs function.
Eventually, we built three circRNA-miRNA-mRNA regulatory axis, circ_0001060-miR-203a-5p-TRIM21, circ_0001060-miR-208b-5p-MAP3K5, and circ_0001060-miR-203a-5p-PRKX. Previous studies have identified the tumor-suppressive role of miR-203a-3p in pancreatic ductal adenocarcinoma and OS regulated by different circRNAs (Wu et al, 2019; Zheng et al, 2022). Despite miR-203a-5p and miR-203a-3p share a same stem-loop pre-miRNA, their biological functions may be inconsistent (Kozomara et al, 2019), whereas miR-208b is more involved in the regulation of skeletal muscle and cardiovascular system (Fu et al, 2020; Schulte et al, 2019).
A study indicated a positive role of TRIM21 in OS cells by promoting autophagy (Zhang et al, 2021). MAP3K5, also known as apoptosis signal-regulating kinase 1, has emerged as important players in hepatocyte injury, inflammation, and fibrosis in nonalcoholic steatohepatitis; however, its role in cancer development remains poorly understood (Fujimoto et al, 2021; Harrison et al, 2020; Lan et al, 2022; Yao et al, 2012). And, PRKX, as a novel cAMP-dependent protein kinase member, may play a role in bone carcinogenesis (Huang et al, 2016; Liu et al, 2016). Considering these points, what we obtained need to be further explored in the future.
Conclusions
Collectively, our research identified a new circRNA, circ_0001060. And, circ_0001060 probably act as a new tumor biomarker for OS. Functionally and mechanistically, circ_0001060 promotes the progression of OS, and its possible mechanisms might be acting as a miRNAs sponge.
Footnotes
Authors' Contributions
Y.L. and T.X. designed the research. H.L. and Q.H. performed the experiments. The article was drafted by H.L. and revised by Y.L., T.X., W.F., and B.L. H.T., K.L., Y.Q., and F.L. collected and recorded clinical data and analyzed statistical data. F.T. and J.Z. performed the bioinformatics analysis.
Disclosure Statement
No competing financial interests exist.
Funding Information
This study was supported by the Natural Science Foundation of Guangxi Province (2020GXNSFAA259088), Youth Science and Technology Project of the First Affiliated Hospital of Guangxi Medical University (201903038), and “Medical Excellence Award” Funded by the Creative Research Development Grant from the First Affiliated Hospital of Guangxi Medical University.
Supplementary Material
Supplementary Data
Supplementary Table S1
References
Supplementary Material
Please find the following supplemental material available below.
For Open Access articles published under a Creative Commons License, all supplemental material carries the same license as the article it is associated with.
For non-Open Access articles published, all supplemental material carries a non-exclusive license, and permission requests for re-use of supplemental material or any part of supplemental material shall be sent directly to the copyright owner as specified in the copyright notice associated with the article.