
Abstract
Cornus officinalis is a perennial deciduous tree or shrub. Its mature fruits are extracted and used in Traditional Chinese Medicine, called Shanzhuyu. The characteristic active components of C. officinalis include loganin and morroniside, which belong to iridoid glycosides. 3-Hydroxy-3-methylglutaryl-CoA synthase (HMGS) is a key enzyme in the cytoplasmic mevalonate pathway providing the precursor molecules isopentenyl pyrophosphate and dimethylallyl pyrophosphate for isoprenoid biosynthesis such as sterols, triterpenes, and their derivatives such as iridoid glycosides. Different concentrations of methyl jasmonate (MeJA) and ethephon (ETH) solutions were sprayed on C. officinalis seedlings, and the effect of hormones on CoHMGS gene expression was detected by real-time fluorescence quantitative PCR. The quantitative real-time PCR results showed that 750 mg/L ETH treatment had the most significant induction effect on CoHMGS gene expression. The HPLC analysis of extracts revealed that the treatment could also significantly increase the content of morroniside and loganin in the leaves of C. officinalis. By use of a CoHMGS-green fluorescent protein (GFP) fusion construct for heterologous expression in tobacco, laser scanning confocal microscopy revealed a cytoplasmic localization. This preliminary study of the CoHMGS gene could prepare the ground for more precisely elucidating the synthesis of secondary metabolite in C. officinalis.
Introduction
C
Among them, iridoid glycosides show high biological activity, which can protect the nervous system and effectively improve diabetes (Peng et al, 2021; Wang et al, 2018). Cornus iridoid glycosides are rich in medically active compounds, including morroniside, loganin, and corniside. The Pharmacopoeia of the People's Republic of China stipulates that morroniside and loganin are the active ingredients of Cornus (Commission, 2020; Dong et al, 2018). Iridoid glycosides are monoterpene derivatives (Kim and Choi, 2021).
Plants contain a variety of terpenoids, which are produced by complex biosynthetic pathways (Bergman et al, 2019). Terpenoids, also called isoprenoids, are produced by subsequent reactions of a starter C5 molecule dimethylallyl pyrophosphate (DMAPP) to which C5 isopentenyl pyrophosphate (IPP) units are fused (Clomburg et al, 2019). There are two mechanisms to generate IPP and DMAPP in plants, through the cytoplasmic mevalonic acid (MVA) pathway and the plastidial methylerythritol phosphate (MEP) pathway (Tetali, 2019).
IPP and DMAPP C5 units formed by the MVA pathway mostly generate sesquiterpenes, triterpenes, and sterols, whereas these C5 units coming from the MEP pathway lead mostly to monoterpenes, diterpenes, tetraterpenes, etc. (Pu et al, 2021). There is some “cross-talk” between the two pathways and the IPP and DMAPP units they generate can be exchanged (Liao et al, 2016). Such a communication between the MVA and MEP pathways has for instance been validated in Spearmint (Li et al, 2020) and spike lavender (Mendoza-Poudereux et al, 2015).
3-Hydroxy-3-methylglutaryl-CoA synthase (HMGS) catalyzes the condensation of acetoacetyl-CoA and acetyl-CoA to form 3-hydroxy-3-methylglutaryl-CoA (HMG-CoA). It is the second step of the MVA pathway. HMGS plays an important role in the biosynthesis of plant terpenoids and sterols, and is, therefore, essential for plant growth and development (Liao et al, 2014). As a key enzyme in the MVA pathway, HMGS has attracted much attention, and more and more medicinal plant HMGS genes have been cloned and studied. The HMGS gene was cloned from Centella asiatica and its functional activity was verified (Afroz et al, 2022).
The sandalwood HMGS gene was cloned, and the experimental results found that the gene could respond to methyl jasmonate (MeJA) treatment (Niu et al, 2021). A differential expression of the HMGS gene significantly affected the production of active substances in Tripterygium wilfordii (Tong et al, 2019). HMGS gene from Chamaemelum nobile L. was induced by MeJA and salicylic acid (Cheng et al, 2016). The experimental results in Ginkgo biloba showed that GbHMGS could be induced by abiotic stress (ultraviolet B and cold) and hormonal treatment (salicylic acid, MeJA, and ethephon [ETH]) (Meng et al, 2017).
Apparently, exogenous hormones can induce the expression of HMGS genes and thereby affect the content of active components in plants. However, the research on the HMGS gene of C. officinalis stands at the beginning. This study explored the subcellular localization of CoHMGS protein in tobacco leaves, detected the expression level of CoHMGS and the content of morroniside and loganin under exogenous hormone treatment. This study should pave the way for some in-depth analysis of the molecular pathways leading to the biomedically active components in C. officinalis.
Materials and Methods
Hormone sprayed on C. officinalis seedlings
We prepared 100, 200, and 400 μM MeJA solutions (Sigma), and 250, 500, and 750 mg/L ETH solutions (Yuanye, Shanghai). The 2-year-old seedlings of C. officinalis were sprayed with hormone solution, and the control group was sprayed with ddH2O. The leaves of C. officinalis were collected at 1, 3, 5, and 7 days after treatment, and quick frozen in liquid nitrogen for use.
Quantitative reverse transcription-PCR analysis of CoHMGS expression levels
Total RNA was extracted from the leaves of C. officinalis according to the kit steps (Nobelab, Beijing), and cDNA was prepared according to the protocol provided by Aidlab (Beijing). Primer Premier 5.0 software was used to design primers to detect the expression level of CoHMGS, the reference gene was GAPDH, and the primer sequences are given in Table 1.
Primers Used for Quantitative Real-Time PCR in This Study
The relative expression levels of target genes were detected by using the SYBR Green reagent (Yeasen, Shanghai) and with the aid of a Roche Light Cycle 96 fluorescence quantitative PCR instrument. Three replicates were set for each treatment, the data processing method was 2−ΔΔCt, data for quantitative real-time-PCR are shown as mean ± SE (n = 3).
Determination of morroniside and loganin content
Eighty percent methanol was used to extract the active substances from the leaves of C. officinalis, and the reference substances of morroniside and loganin were purchased from Macklin company, and each analysis was repeated thrice. The content of morroniside and loganin was determined by the use of an Agilent HPLC system under the following conditions: VWD UV detector, mobile phase methanol and water (25:75), detection wavelength at 240 nm, column oven at 30°C, injection volume was 10 μL, the flow rate was set to 1 mL/min, and the detection time of each test sample was set to 40 min.
Subcellular localization of CoHMGS
The expression vector used for subcellular localization experiments was pCAMBIA 1300-green fluorescent protein (GFP). The stop codon of the open reading frame of CoHMGS was removed and Primer Premier 5.0 software was used to design primers as given in Table 2. Kpn I-CoHMGS-Xba I and pCAMBIA 1300-GFP were digested with Kpn I and Xba I restriction enzymes (TaKaRa), and then ligated with T4 ligase to obtain the pCAMBIA 1300-CoHMGS-GFP fusion expression vector.
Primers Used for Subcellular Localization in This Study
Note: The underlined places refer to the sequences of the enzyme cleavage sites added in the PCR primers.
The recombinant vector was transformed into Agrobacterium GV3101, the Agrobacterium liquid was used to infect the lower epidermis of tobacco leaves, and cultured in the dark for 2 days. The subcellular localization of CoHMGS-GFP was observed by using a confocal laser microscope (Olympus FV 3000, Japan) with selected excitation light source at 488 nm.
Results
CoHMGS transcript level changes in C. officinalis under the induction of MeJA, ETH
The expression levels of CoHMGS in C. officinalis leaves treated with different concentrations of MeJA and ETH for different days were detected by quantitative real-time PCR (qPCR), as shown in Figure 1. The results showed that the induction effect of 750 mg/L ETH on the expression level of CoHMGS continued and was the most significant. The expression level reached the maximum value on the third day, which was 3.6-fold that of the control group. The expression level decreased in the remaining 2 days, but was always higher than that of the control group.
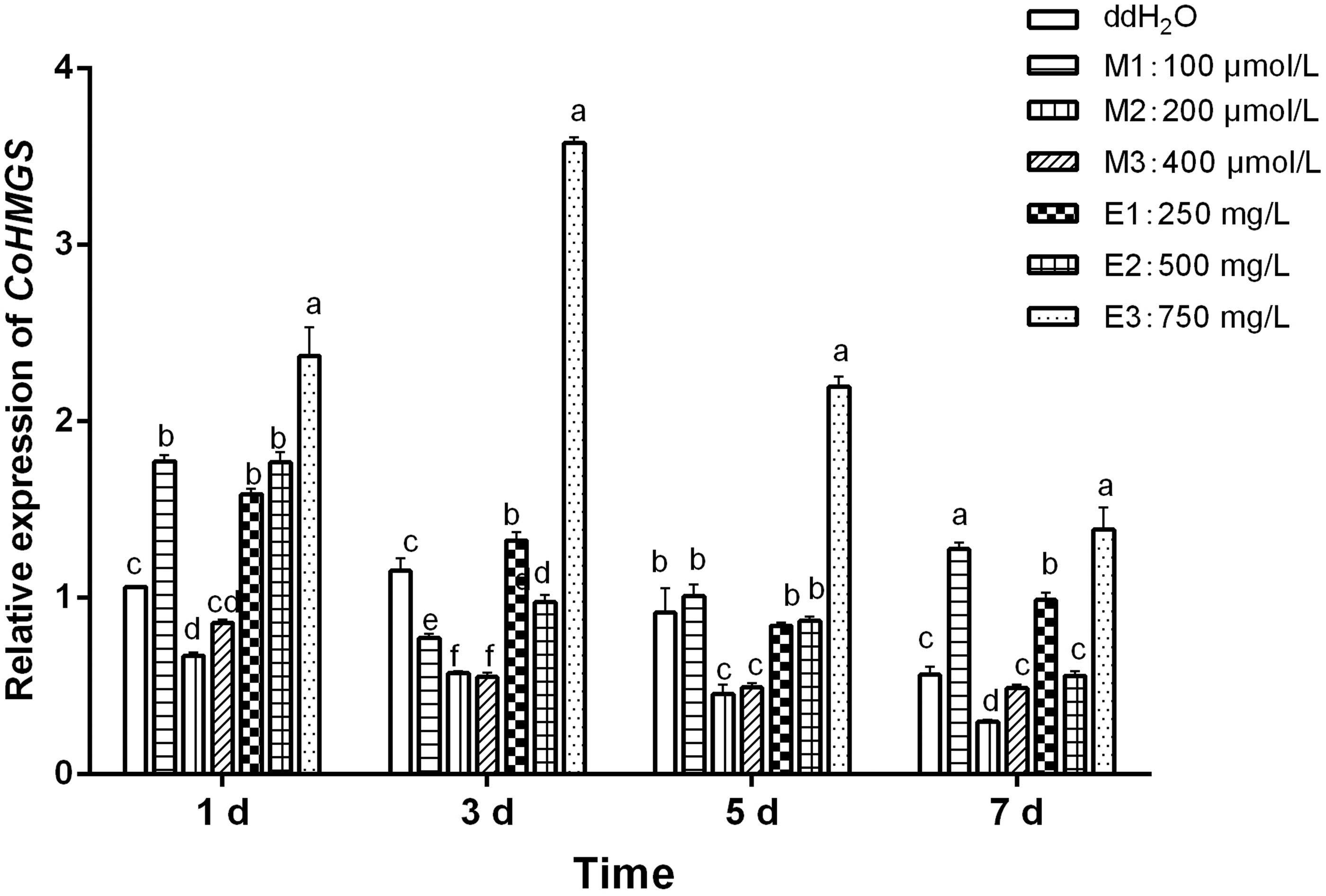
Analysis of expression levels of the CoHMGS in Cornus officinalis leaves treated with exogenous methyl jasmonate and ethephon. Different lowercase letters indicate significant differences between treatments (p < 0.05). CK, control check; E, ethephon treatment; HMGS, 3-hydroxy-3-methylglutaryl-CoA synthase; M, methyl jasmonate treatment.
On the first day and the seventh day after 100 μM MeJA treatment, the expression level of CoHMGS gene was significantly higher than that of the control group, but the induction effect did not last. The expression level of CoHMGS gene was significantly decreased within 7 days after 200 and 400 μM MeJA treatment. Most likely, the high concentrations of MeJA inhibited the expression of CoHMGS. In contrast, the expression of CoHMGS was significantly increased after a treatment with 250 mg/L ETH, although this increase remained limited.
Five hundred milligrams per liter ETH treatment did not exert any obvious effect on the induction of CoHMGS expression. Apparently, different concentrations of exogenous hormone sprays differentially affected the expression level of CoHMGS. Clearly, applying of concentration gradients is a mandatory to identify the optimum treatment.
Morroniside and loganin content changes in C. officinalis under the induction of MeJA, ETH
After the qPCR results, samples with the most significant induction of CoHMGS gene expression were selected for subsequent experiments. Morronin and loganin in C. officinalis leaves treated with 750 mg/L ETH were quantified by HPLC analysis, and the result is shown in Figure 2. Figure 2A shows that 750 mg/L ETH treatment increased the content of morroniside in the leaves of C. officinalis on the first and fifth days. Figure 2B shows that the induction effect of the treatment on loganin content was most obvious on the fifth day. The mentioned results prove that 750 mg/L ETH could promote the content of morroniside and loganin in leaves of C. officinalis.

HPLC detection of active components in the leaves of C. officinalis.
Subcellular localization analysis of CoHMGS protein
The recombinant expression vector pCAMBIA 1300-CoHMGS-GFP was constructed and transformed into Agrobacterium tumefaciens, and the A. tumefaciens, and its liquid was used to infect the lower epidermis of tobacco leaves. Tobacco was treated with pCAMBIA 1300-GFP and pCAMBIA 1300-CoHMGS-GFP Agrobacterium solution, respectively, and the observation results are shown in Figure 3. Taking the pCAMBIA 1300-GFP vector as a control, it was observed that the luminescence position of the pCAMBIA 1300-CoHMGS-GFP vector was in the cytoplasm of tobacco, indicating that the expression position of CoHMGS protein is in the cytoplasm, which is consistent with the prediction result of the software.
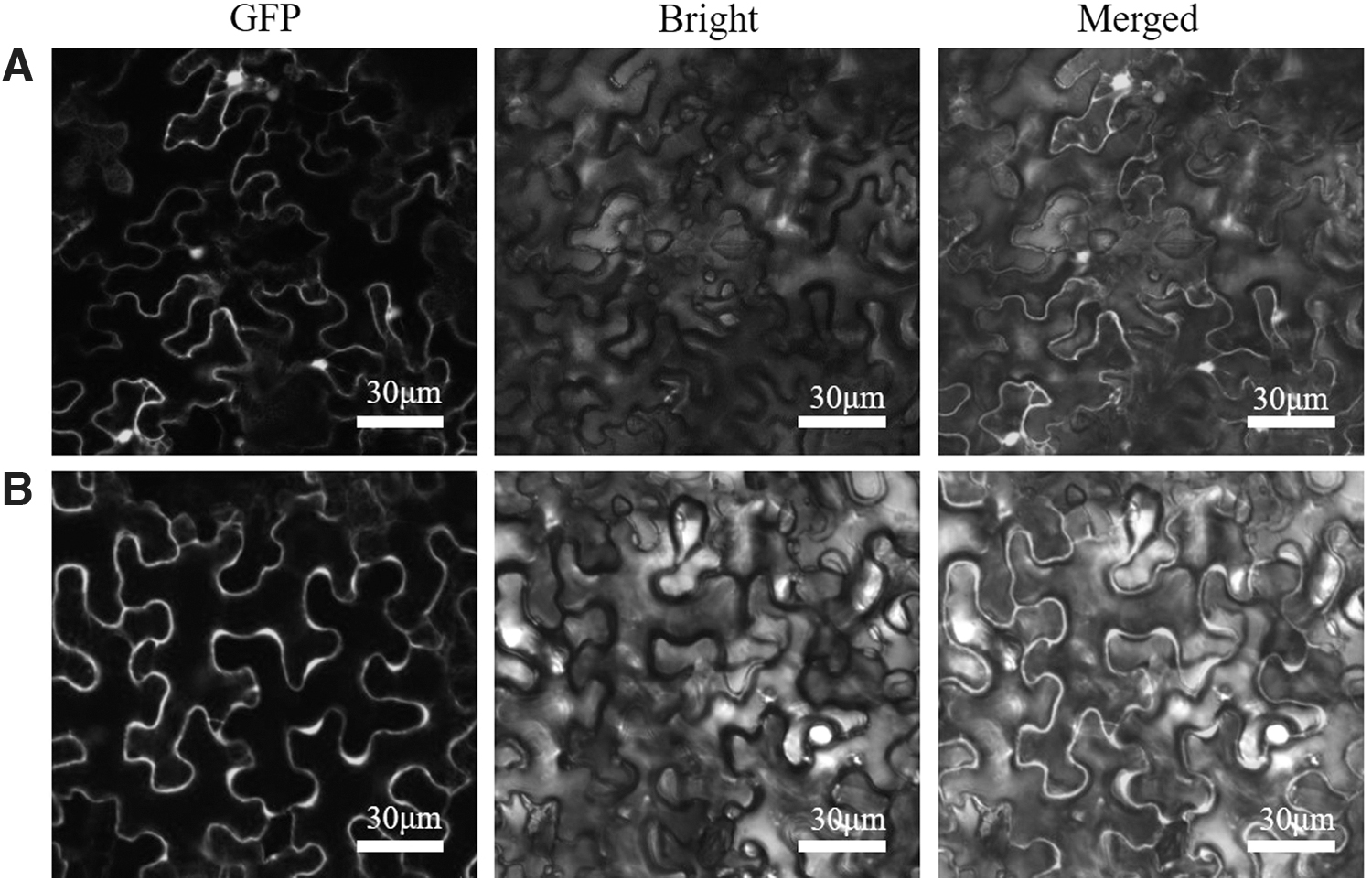
Subcellular localization of CoHMGS-GFP protein in tobacco leaves.
Discussion
The importance of the HMGS gene has been validated in a variety of plant species. It was found that Arabidopsis mutants lacking the gene encoding HMG-CoA synthase resulted in male gametophyte lethal during pollen tube elongation (Ishiguro et al, 2010).
Overexpression of the Brassica juncea HMGS1 gene in Arabidopsis thaliana increased the expression levels of some stress resistance genes, and increased the sterol content and stress tolerance of plants (Alex et al, 2000; Wang et al, 2012). All subsequent experiments used B. juncea HMGS1 (Nagegowda et al, 2004; Pojer et al, 2006). In Ganoderma lucidum, the HMGS gene plays an important role in the biosynthesis of ganoderic acids (Meng et al, 2019).
HMGS is an important enzyme in the MVA pathway, and studying the localization of this protein in cells is the key to understanding the function of this gene. The CoHMGS protein was predicted to be located in the cytoplasm by online software (Cell-Ploc 2.0). The green fluorescence emitted by the CoHMGS protein was observed under the laser confocal microscope, showing uneven green at the cell edge, indicating that CoHMGS is expressed in the cytoplasm of transformed tobacco cells. This result is consistent with the expression of B. juncea HMGS1 protein in onion and tobacco cells (Nagegowda et al, 2005).
Previous reports showed that when ETH was added to the culture medium of G. lucidum, the growth of mycelium in the treatment group was significantly better than that in the control group, and the content of G. lucidum acid was also increased (Zhang et al, 2017). G. biloba seedlings were treated with MeJA and ETH solution, respectively, and it was found that the expression level of GbHMGS1 and the content of total terpene trilactones were increased within 8 days after treatment (Meng et al, 2017).
In this study, it was found that 100 μM MeJA and three concentrations of ETH significantly increased the expression level of CoHMGS on the first day after treatment. Over time, only 750 mg/L ETH had the best positive induction effect on CoHMGS. The HPLC-based analyses demonstrated that 750 mg/L ETH significantly increased the contents of morroniside and loganin in the leaves of C. officinalis These results are basically consistent with the phenomena observed in other plants.
Owing to different experimental environments and materials, the optimum hormone concentration that potentially increased the expression of key genes in the biosynthesis of active ingredients needs to be determined. Furthermore, the effect on the content of compounds of interest may also vary, which requires always an adopted experimental strategy.
Footnotes
Disclosure Statement
No competing financial interests exist.
Funding Information
This research was supported by the National Natural Science Foundation of China (U1404829), key project at central government level. The ability of establishment of sustainable use for valuable Chinese Medicine resources (2060302) and Natural Science Foundation of Henan Province (202300410151). This research was also supported by a special fund for the construction of technical systems of the Traditional Chinese Medicine industry in Henan Province (Yucaike 2022-24). Henan Province Chinese Medicine industry science and technology commissioner service group (Yuke 2022-47).