
Abstract
Improving livestock and poultry growth rates and increasing meat production are urgently needed worldwide. Previously, we produced a myostatin (MSTN) and fibroblast growth factor 5 (FGF5) double-knockout (MF−/−) sheep by CRISPR Cas9 system to improve meat production, and also wool production. Both MF−/− sheep and the F1 generation (MF+/−) sheep showed an obvious “double-muscle” phenotype. In this study, we identified the expression profiles of long noncoding RNAs (lncRNAs) in wild-type and MF+/− sheep, then screened out the key candidate lncRNAs that can regulate myogenic differentiation and skeletal muscle development. These key candidate lncRNAs can serve as critical gatekeepers for muscle contraction, calcium ion transport and skeletal muscle cell differentiation, apoptosis, autophagy, and skeletal muscle inflammation, further revealing that lncRNAs play crucial roles in regulating muscle phenotype in MF+/− sheep. In conclusion, our newly identified lncRNAs may emerge as novel molecules for muscle development or muscle disease and provide a new reference for MSTN-mediated regulation of skeletal muscle development.
Introduction
Meat products serve as important sources of protein for many humans by providing various nutrients. However, the worldwide demand for meat products is increasing dramatically from explosive population growth and the ongoing coronavirus disease 2019 (COVID-19) pandemic. Therefore, improving the livestock and poultry growth rates and increasing meat production are urgently needed. Myogenesis is the process by which skeletal muscle cells develop to form muscle. Skeletal muscle is a muscle group attached to the skeleton, which is composed of a large number of muscle fibers, and each muscle fiber is composed of a large number of myofibrils. The sarcolemma and the basement membrane surround the myofibril bundles from the inside to the outside, and there are a large number of skeletal muscle satellite cells between these two membranes.
Under normal physiological conditions, skeletal muscle satellite cells are in a quiescent state. When activated by physiological or pathological stimuli, the quiescent skeletal muscle satellites are activated. A portion of them return to the quiescent state, while another portion will form contractible and beating muscle tubes through proliferation, migration, nuclear rearrangement, differentiation, and fusion. These processes are crucial for the proper development of skeletal muscle. In fact, myogenesis is an extremely complex and delicate process that is regulated by numerous regulatory factors, such as the paired box families (Pax3/7), myogenic regulatory factors (Myogenin, MyoD, Myf5, and MRF4/6), myocyte enhancer factor 2 (MEF2) family proteins, and myostatin (MSTN). Together, these factors are synergistically involved in regulating the expression of muscle-specific genes and further regulating the development of skeletal muscle, thereby significantly affecting meat production of livestock and poultry (Buckingham, 2006).
MSTN has been well known as a negative regulator of muscle growth and development by limiting the number and size of skeletal muscle fibers after birth. MSTN is highly conserved in mammals, and natural or artificial genetic mutations can result in increased skeletal muscle weight and an obvious “double-muscle” phenotype. This phenomenon has been reported in many species, including cattle, sheep, and pigs, rabbits, and humans (Chen et al, 2021b; Fan et al, 2022). Among these, the most famous are Belgian blue cattle, Piedmontese cattle, Marchigiana cattle, Norwegian white sheep, and Texel sheep. Those evidence suggests that MSTN has immeasurable commercial value for improving livestock meat production and helping address the human food crisis (Chen et al, 2021b; Fan et al, 2022).
It is currently accepted that MSTN requires a complex series of crosstalk reactions for signal transduction to the nucleus. First, the MSTN dimer binds to ActRIIB and then to ALK4/5 to form a complex (Chen et al, 2021b). The Smad pathway is the classical signaling pathway for MSTN, where Smad2/3/4 enters the nucleus to regulate the expression of target genes, and different transcription factors can bind to the Smad2/3/4 complexes, leading to various functions of the Smad signaling pathway (Chen et al, 2021b). In addition, MSTN can also regulate the expression of downstream target genes through the PI3K-AKT-mTOR, Ras-MEK-ERK, p38 MAPK, and JNK signaling pathways (Chen et al, 2021b).
However, the molecular mechanism of MSTN-mediated negative regulation of skeletal muscle growth and development still remains controversial. It has been reported that MSTN negatively regulates the proliferation (Taylor et al, 2001; Thomas et al, 2000) and differentiation (Langley et al, 2002) of skeletal muscle satellite cells. The overexpressed MSTN inhibits the protein expression levels of MyoD and Myogenin to inhibit the activation of myoblast differentiation (Joulia et al, 2003). Interestingly, MSTN is a downstream target of MyoD, which may trigger myoblasts to exit the cell cycle by regulating the expression of MSTN (Spiller et al, 2002). In addition, MSTN decreases the diameter of myotubes (Trendelenburg et al, 2009). The dynamic turnover process of skeletal muscle protein synthesis and degradation is essential for muscle growth and development.
Recent studies have revealed that Smad2 and Smad3 serve as transcription factors that mediate the effect of MSTN on muscle mass, suggesting the effect of the MSTN pathway on muscle protein degradation (Sartori et al, 2009). Further studies demonstrated that the inhibitory effect of the MSTN-Smad2/3 axis on the IGF1-Akt-mTOR pathway, suggesting that MSTN, FoxOs, and Smads synergize to regulate muscle mass by affecting muscle protein synthesis and degradation (Amirouche et al, 2009; Sartori et al, 2009).
In addition to MSTN and myogenic regulators, long noncoding RNAs (lncRNAs) are also widely involved in the process of myogenesis (Li et al, 2018b; Simionescu-Bankston and Kumar, 2016). lncRNAs are a class of RNA transcripts with a length more than 200 nt and have no protein coding potential. With tissue and spatiotemporal specificity, lncRNAs are ubiquitous in organisms with poor evolutionary conservation among different species (Chen et al, 2019). With the mysterious functions of lncRNAs gradually be shed light, accumulating evidence has suggested that lncRNAs may be a gatekeeper of skeletal muscle growth and development, because of its undeniable role in various processes during myogenesis, including quiescence, activation, proliferation, fusion, and differentiation.
Current studies have shown that lncRNAs can regulate skeletal muscle development in a variety of ways. For example, lncRNAs can act as molecular sponges to competitively bind microRNAs or proteins, and further regulating the expression or activity of target genes (Sun et al, 2016). lncRNAs can also mediate epigenetic modifications of target genes (Wang et al, 2015). Furthermore, lncRNAs can regulate the splicing, stability, and abundance of target mRNAs (Blin-Wakkach et al, 2001; Wang et al, 2016). However, most studies of lncRNAs focused on humans and mice (Cabili et al, 2011; Chen et al, 2021a), although more and more lncRNAs in livestock species have been identified. However, few lncRNAs have been uncovered in sheep, and the NONCODE database lacks information on sheep lncRNAs (Zhao et al, 2016).
Yet, a study on sheep longissimus dorsi muscle at 60, 90, and 120 days of gestation, the day of birth, and 360 days after birth identified a total of 694 muscle development-associated differentially expressed lncRNAs (DELs) (Yuan et al, 2020). Similarly, a study on lncRNAs in the gastrocnemius muscle from fetal (days 85 and 120), newborn, and adult of Texel and Ujumqin sheep identified 967 DELs, which were considered to be regulators of muscle differences observed between these two sheep breeds (Li et al, 2018a). In addition, by comparing skeletal muscle lncRNAs among Dorper sheep, Qianhua Mutton Merino sheep, and Small-tailed Han sheep, a total of 29 lncRNAs were screened and related to cell proliferation, muscle development and metabolism, and apoptosis (Chao et al, 2018).
In fact, the currently identified functional lncRNAs that can regulate skeletal muscle development in sheep are very limited. For example, lnc-SEMT modulates IGF2 expression by sponging miR-125b to promote sheep muscle development and growth (Wei et al, 2018). In addition, lncRNA CTTN-IT1 can elevate skeletal muscle satellite cell proliferation and differentiation by acting as a competing endogenous RNA for YAP1 by absorbing miR-29a in Hu sheep (Wu et al, 2020). Overall, identifying muscle development-related lncRNAs may provide new targets references for meat sheep breeding.
To improve both meat and wool production, we previously produced a MSTN and fibroblast growth factor 5 (FGF5) double-knockout sheep using the CRISPR Cas9 system, which generated a MSTN and FGF5 dual-gene biallelic homozygous (MF−/−) mutant (Zhao et al, 2022). Both the MF−/− sheep and the MF+/− sheep, the F1 generation of MF−/− sheep, showed an obvious “double-muscle” phenotype. In this study, we identified the expression profile of lncRNAs in gluteus medius of MF+/− sheep, screened four crucial candidate lncRNAs that can regulate myogenic differentiation and skeletal muscle development, and recognized them as key gatekeepers of muscle phenotype in MF+/− sheep, which will provide a new reference for MSTN regulation of skeletal muscle development.
Materials and Methods
Animals and muscle tissue sample collection
A total of six 12-month-old female Dorper sheep were used in this study. They were divided into two groups with three replicates in each group, including three wild-type (WT) sheep and three MF+/− sheep. All sheep were raised in the experimental base of Institute of Animal Husbandry and Veterinary Medicine, Tianjin Academy of Agricultural Sciences. All sheep are raised in accordance with the national feeding standard NT/T815-2004. All procedures performed in this study were consistent with the National Research Council Guide for the Care and Use of Laboratory Animals. All experimental animal protocols in this study were approved and performed in accordance with the requirements of the Animal Care and Use Committee at China Agricultural University (AW02012202-1-2). The gluteus medius was harvested from WT and MF+/− sheep. All samples were immediately frozen in liquid nitrogen and then stored at −80°C until analysis.
Total RNA isolation, cDNA library construction, and RNA-sequencing
Total RNA was isolated from tissues and cells using TRIzol reagent (Sangon Biotech, Shanghai, China). Specifically, 1 mL TRIzol was added to each sample, then lysed at room temperature for 5 min. Next, 200 μL trichloromethane was added and the EP tubes were vortexed for 15 s, and then incubated for 3 min at room temperature. Following a 15 min centrifugation at 4°C and 12,000 rpm, each supernatant was carefully removed and added to a fresh tube. Then, an equal volume of isopropanol was added and the precipitated RNA at room temperature for 20 min and then centrifuged at 4°C 12,000 rpm for 10 min to collect the precipitate. Next, 1 mL ice-cold 75% ethanol was added to wash the precipitate, which was then collected and centrifuged at 4°C 12,000 rpm for 3 min. After evaporating the ethanol, 45 μL RNase-free water was added to dissolve the RNA.
Next, the Agilent 4200 instrument was used to accurately assess the purity and concentration of the RNA samples. The rRNA was removed from all samples with RINe > 8.0 and no less than 2 μg of total yield. RNA purification, cDNA library construction, and RNA sequencing were conducted by Berry Genomics company. Specifically, the NEBNext® UltraTM Directional RNA Library Prep Kit was used for Illumina® to make rRNA-free RNA sequencing libraries. Then, the first-strand cDNA was synthesized using random hexamer primers and M-MuLV reverse transcriptase (RNaseH-), and cDNA second-strand synthesis was performed using DNA polymerase I and RNase H. Each library was paired-end sequenced (2 × 150 bp) on an Illumina Novaseq6000 platform.
Quality control, lncRNAs identification, and differential expression analysis
The stringent stepwise filtering pipeline was used to identify lncRNAs (Fig. 1A). Raw data were transformed into clean reads by removing reads containing adapter, reads containing poly-N, and low-quality reads from raw data using FastQC v0.11.8 and Trimmomatic v0.39 (Bakhtiarizadeh and Salami, 2019; Bolger et al, 2014). In addition, the Q20, Q30, GC-content and sequence duplication level of the clean data were calculated (Li et al, 2020). High-quality clean reads were used for subsequent analysis. The genome index was constructed using Hisat2 v2.1.0 software and the paired end clean reads were mapped to the sheep reference genome (
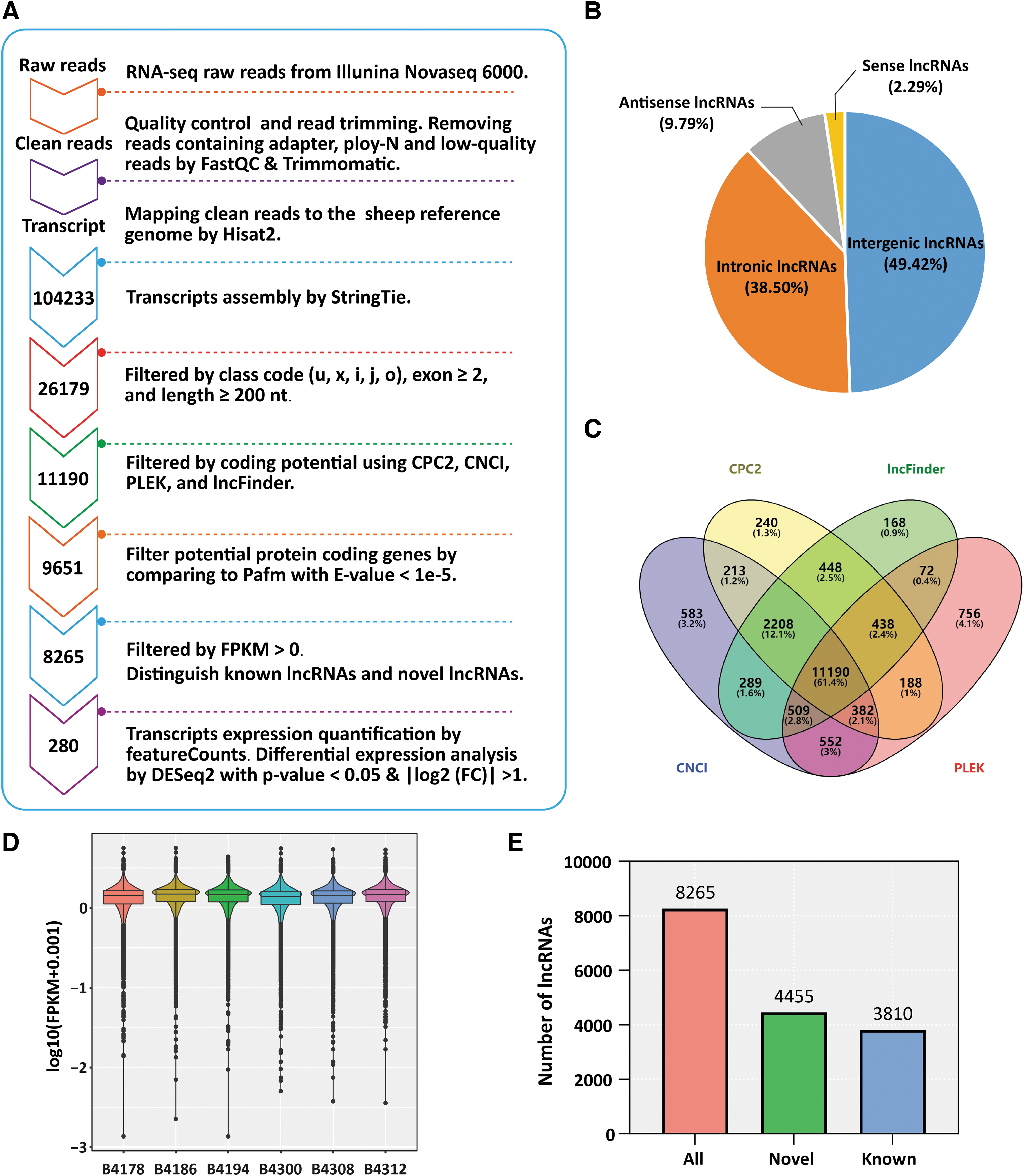
LncRNA filtering pipeline and characteristics.
Further, the gffcompare v0.11.5 was used to identify transcripts in the “u,” “x,” “i,” “j,” and “o” classes, and transcripts overlapped with annotation genes were detected based on the reference transcriptome. Among them, the transcripts with the number of exons ≥2 and the length ≥200 nt were retained for further analysis. Subsequently, the coding capacity of these transcripts was predicted by CPC2 (v1.0.1), CNCI (v2.0), PLEK (v1.2), and lncFinder, and their intersection was recognized as potential lncRNAs. To further determine the noncoding properties of potential lncRNAs, the Pfam database with e-value <1e-5 was used for filtering possible protein coding genes.
Furthermore, the transcripts with FPKM >0 were retained as lncRNAs with typical characteristics. To distinguish known lncRNAs, the potential lncRNAs transcripts were matched with the Ensembl sheep reference annotation GTF file (release 103) (Bakhtiarizadeh and Salami, 2019). With e-value ≤1e-10, min-identity = 80%, and min-coverage = 50%, all transcripts that grouped “gene_biotype = lncRNA” in the GTF file were recognized as known lncRNAs. Next, the featureCounts v2.0.1 software was used for transcripts expression quantification (Liao et al, 2014). In addition, the DESeq2 1.36.0 software was used for differential expression analysis using p-value <0.05 and | log2 Fold Change |>1.
Prediction of potential lncRNA target genes and functional enrichment analysis
LncRNAs function mainly through cis- and trans-regulation of target genes. Thus, the bedtools software was used to extract cis-target genes within 100 kb upstream and downstream of lncRNAs, and the nearest genes were considered as cis-target genes (Guenzl and Barlow, 2012). The trans roles of lncRNAs are not related to their positional relationships with protein coding genes, but to the protein coding genes that are coexpressed with lncRNAs. Therefore, a custom R script was used to calculate the expression correlation between lncRNAs and the differentially expressed genes (DEGs) to analyze the trans-target of lncRNAs. The DEGs with |correlation| > 0.95 and p-value <0.001 were recognized as potential trans-target genes of lncRNAs (Li et al, 2020).
Finally, the cis-target genes and trans-target genes were combined as potential target genes (PTGs) of lncRNAs. Functional annotation of the lncRNAs was performed using the functional enrichment analysis of their related PTGs. The DAVID 2021 was used to perform functional enrichment analysis for the PTGs for gene ontology (GO) and Kyoto Encyclopedia of Genes and Genomes (KEGG) pathway categories. Only terms with p-value <0.05 were considered to be significantly enriched.
Cell isolation, culture, transfection, and induced differentiation
Sheep skeletal muscle satellite cells were isolated as we previously described (Chen et al, 2021a). The resuscitated skeletal muscle satellite cells were resuspended in growth medium (GM) containing 20% fetal bovine serum (Gibco, Grand Island, NY) and 1% penicillin/streptomycin solution (Gibco) in DMEM/F12 (Gibco) and incubated at 37°C in a 5% CO2 incubator. After the cells reached 70% confluence, the differentiation medium (DM) was changed to induce differentiation. The DM contained DMEM high glucose (Gibco), 2% horse serum (Gibco), and 1% penicillin/streptomycin solution. For cell transfection, the cells were transfected with siRNA using Lipofectamine 3000 (Invitrogen) when they were cultured to 60–70% confluence in 12-well, plates. In all cell culture plates, the final concentrations used for siRNA were 100 nM. All cells were collected for detection after transfection 24 h.
Real-time quantitative PCR
The first-strand cDNA was prepared using PrimeScript™ RT reagent Kit with gDNA Eraser (Takara, Beijing, China). qPCR was performed using 2 × SYBR Green qPCR Mix (Low ROX) (Aidlab Biotechnologies, Beijing, China) using a Stratagene Mx3000P (Agilent Technologies, Santa Clara, CA). GAPDH mRNA was used as the endogenous control, and the 2−ΔΔCt method was used to calculate the relative expression levels of lncRNAs. All primers are listed in Supplementary Table S1.
Immunofluorescence staining
The immunofluorescence staining was performed as described previously (Chen et al, 2019). In brief, the cells were fixed in 4% paraformaldehyde for 30 min. Then permeabilized in 0.1% Triton X-100 for 20 min if required. In general, proteins on cell membranes are not required to be permeable. Subsequently, the fixed and permeabilized cells were blocked with 5% normal goat serum for 30 min at room temperature, and then incubated with primary antibody at 4°C overnight. The next day, the cells were incubated with the fluorescent labeled secondary antibody at 37°C for 1 h in the dark. Then, the nuclear was counterstained with DAPI and the immunofluorescence images of five visual fields were randomly collected. All pirmary antibody information is as follows: anti-Pax7 (DSHB, Mouse), anti-MyoD1 (Affinity, AF7733, Rabbit), anti-MyoG (DSHB, F5D, Mouse), and anti-Myosin VIIa (anti-MyHC) (abcam, Ab3481, Rabbit).
Statistical analysis
All results are presented as the mean ± standard error of the mean. Statistical analyses of differences between groups were performed using two-tailed Student's t-test. *p < 0.05, **p < 0.01, and ***p < 0.001.
Results
Summary statistics of RNA-seq data
Previously, we prepared a MSTN and FGF5 double-knockout sheep, which is a MSTN and FGF5 dual-gene biallelic homozygous (MF−/−) mutant (Zhao et al, 2022). Both the MF−/− sheep and the MF+/− sheep, the F1 generation of MF−/− sheep, showed an obvious “double-muscle” phenotype. In this study, we aimed to identify potential functional lncRNAs that can affect this phenotype. Therefore, we constructed a cDNA library from six sheep gluteus medius samples, in which B4178, B4186, and B4194 were from WT sheep, and B4300, B4312, and B4316 were from MF+/− sheep (Supplementary Table S2). Raw data were transformed into clean data by removing reads containing adapter, poly-N, and low-quality reads. The Q20 and Q30 were more than 90% (Supplementary Table S2), and the proportion of rRNA was less than 4% (Supplementary Table S2), indicating that the RNA-seq data were suitable for subsequent analysis.
Subsequently, Hisat2 software was used to map clean reads to the sheep reference genome (Oar rambouillet v1.0), and the unique mapping rates were greater than 86%, and total mapping rates were greater than 98% (Supplementary Table S2). The raw sequence data reported in this article have been deposited in the Genome Sequence Archive (Genomics, Proteomics & Bioinformatics 2021) in National Genomics Data Center (Nucleic Acids Res 2022), China National Center for Bioinformation/Beijing Institute of Genomics, Chinese Academy of Sciences (GSA: CRA008539) that are publicly accessible at
Identification of lncRNAs in gluteus medius samples
To identify lncRNAs, we developed a stringent filtering pipeline to screen transcripts with lncRNA characteristics (Fig. 1A). According to these characteristics of lncRNAs, 26179 transcripts were filtered by assembling and screening with samtools, StringTie and gffcompare (Fig. 1A). Among these transcripts, 49.42% were from intergenic regions, 38.5% were from intronic regions, and 2.29% and 9.79% were sense and antisense lncRNAs, respectively (Fig. 1B). Subsequently, the coding capacity of these transcripts was calculated by CPC2, CNCI, PLEK, and lncFinder, and a total of 11,190 potential lncRNAs transcripts were strictly filtered (Fig. 1A, C). To further determine the noncoding properties of these potential lncRNAs, we also matched them to the Pfam database with a strict standard of e-value <1e-5, and filtered for possible protein coding genes, and a total of 9651 transcripts were identified as potential lncRNAs (Fig. 1A).
Finally, after removing low-expression transcripts with FPKM >0, a total of 8265 transcripts were recognized as lncRNAs with typical characteristics (Fig. 1A), and their expression levels in each sample also corresponded to known lncRNA characteristics (Fig. 1D). In addition, to distinguish the known lncRNAs, the 8265 lncRNAs were matched with the Ensembl sheep reference annotation GTF file (release 103). With e-value ≤1e-10, min-identity = 80%, and min-coverage = 50%, all transcripts that grouped as “gene_biotype = lncRNA” in the GTF file were recognized as known lncRNAs. The results showed that 4455 transcripts were recognized as novel lncRNAs, and 3810 transcripts were known lncRNAs (Fig. 1E).
The MSTN and FGF5 double-knockout causes differential expression of lncRNAs
To further identify sheep muscle development-related lncRNAs and reveal the molecular mechanism by which MSTN and FGF5 double-knockout causes the “double-muscle” phenotype, the differential expression of lncRNAs in the gluteus medius of WT and MF+/− sheep was performed. The principal component analysis (PCA) of all samples showed relatively good repeatability within sample groups and good discrimination between WT and MF+/− samples (Fig. 2A). Quantitative expression of transcripts was analyzed using featureCounts. Subsequently, DESeq2 was used for differential expression analysis. With p-value <0.05 and | log2 Fold Change |>1 as the screening criteria, a total of 280 DELs were screened, including 78 upregulated DELs and 202 downregulated DELs (Fig. 2B). The cluster analysis of top 30 DELs showed that the samples were clustered into WT groups and MF+/− groups, with obvious differences between the groups (Fig. 2C).

Differential expression analysis of lncRNAs and functional analysis of their target genes.
Prediction of target genes and characterization of potential functions of the lncRNAs
To assess the potential functions of these lncRNAs in regulatory processes, the cis- and trans-regulated target genes of lncRNAs were predicated. The bedtools software was used to extract cis-target genes within 100 kb upstream and downstream of the lncRNAs, and then we performed differential expression analysis on the 12,047 extracted PTGs of lncRNAs. With |log2 Fold Change| > 1 and p-value <0.05, a total of 150 differentially expressed cis-target genes were screened (Fig. 2D).
In addition, our previous study identified 295 DEGs in the gluteus medius of WT and MF+/− sheep (data not published). Here, the Pearson correlation analysis was used to analyze the expression correlation between DELs and DEGs among samples, and the DEGs with |correlation| > 0.95 and p-value <0.001 were recognized as potential trans-target genes of DELs. The results showed that 216 DELs were significantly positively or negatively correlated with 192 DEGs (Fig. 2D). Finally, we combined the cis-target genes with the trans-target genes and identified a total of 256 PTGs of the lncRNAs (Fig. 2D).
To further characterize the potential functions of the lncRNAs, DAVID 2021 was used to perform functional enrichment analysis on the 256 PTGs. GO enrichment analysis showed that the 256 PTGs were significantly enriched in positive regulation of calcium ion import, regulation of muscle contraction, cardiac muscle contraction, muscle contraction, skeletal muscle cell differentiation, and regulation of the force of heart contraction and so on in biological process (BP); in cellular component, these PTGs mainly belonged to extracellular region, cytoplasm, I band, external side of plasma membrane, neuron projection, membrane, perinuclear region of cytoplasm, and extracellular matrix; and molecular function (MF) mainly included protein homodimerization activity, protein binding, nucleotide binding, receptor binding, ATP binding, heat shock protein binding, and CXCR3 chemokine receptor binding (Fig. 2E), indicating that DELs are significantly closely related to muscle contraction, calcium transport, and skeletal muscle cell differentiation.
In addition, KEGG enrichment analysis showed that the 256 PTGs were significantly enriched in cytokine–cytokine receptor interaction, the IL-17 signaling pathway, viral protein interaction with cytokine and cytokine receptors, the TNF signaling pathway, and the p53 signaling pathway (Fig. 2F), suggesting that the DELs are significantly associated with skeletal muscle inflammation, cell apoptosis, and autophagy. In a word, these results suggest the potential roles of the lncRNAs in the “double-muscle” phenotype caused by MSTN and FGF5 double-knockout.
Candidate lncRNAs may be the potential gatekeepers of the muscle phenotype in MSTN and FGF5 double-knockout sheep
To verify the reliability of the RNA-seq data and identify the potential role of these lncRNAs in skeletal muscle development, three upregulated DELs and eight downregulated DELs that were closely related to skeletal muscle development were selected as candidate lncRNAs based on enrichment analysis and the functions of target genes, and the real-time quantitative PCR (RT-qPCR) was performed for verification. The results showed that the expression of three upregulated DELs (Fig. 3A) and eight downregulated DELs (Fig. 3B) at the gluteus medius tissue level were completely consistent with the RNA-seq data, suggesting that the RNA-seq data were reliable. Given that myogenic differentiation of skeletal muscle satellite cells is a critical step in skeletal muscle development, we also validated expression of the 11 candidate lncRNAs at the cellular level.
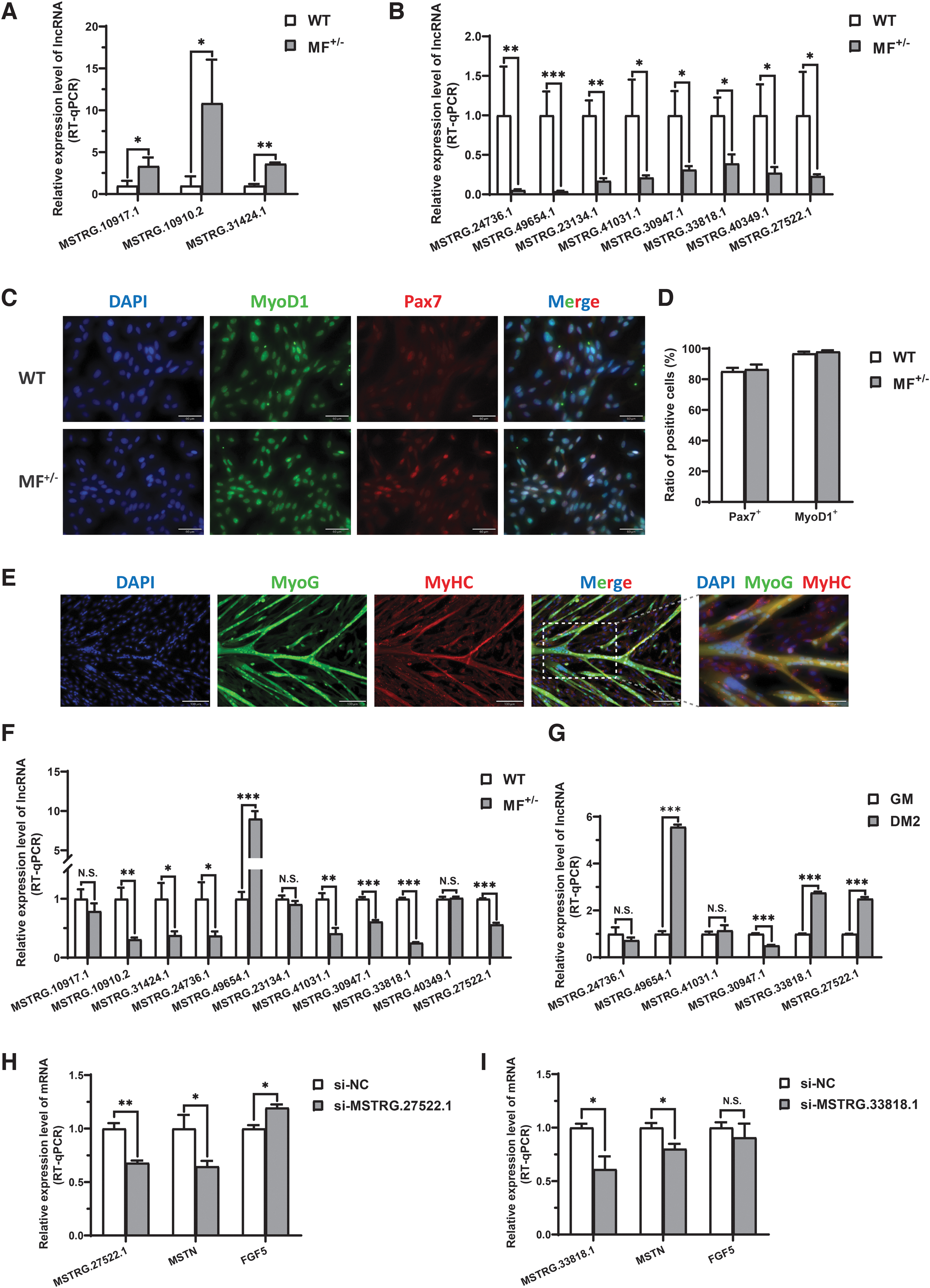
Validation of candidate lncRNAs expression at the tissue and cellular levels.
First, we isolated skeletal muscle satellite cells, and immunofluorescence staining for their markers Pax7 and MyoD1 showed that the ratio of Pax7-positive (Pax7+) cells and MyoD1-positive (MyoD1+) cells in WT and MF+/− cells were more than 85.3% and 96.6%, respectively (Fig. 3C, D), indicating a very high purity of isolated skeletal muscle satellite cells. In addition, the isolated skeletal muscle satellite cells isolated from WT sheep were induced for differentiation in vitro, and followed by the immunofluorescence staining for myogenic differentiation markers MyoG and MyHC. The results showed that the skeletal muscle satellite cells could fuse to form a large number of myotubes after induced differentiation in differentiation medium for 2 days (DM2) (Fig. 3E), which further indicated that the isolated skeletal muscle satellite cells could be fully used to the establishment of subsequent myogenic differentiation models in vitro.
Subsequently, the RT-qPCR results of the 11 candidate lncRNAs in skeletal muscle satellite cells showed that the expression of 8 candidate lncRNAs were significantly different between WT and MF+/− skeletal muscle satellite cells in GM (Fig. 3F).
To further determine their role in myogenic differentiation, we also validated six candidate lncRNAs before and after induced differentiation. The results showed that the expression levels of MSTRG.49654.1, MSTRG.33818.1, and MSTRG.27522.1 were highly significantly (p < 0.01) increased after induced differentiation, while the expression levels of MSTRG.30947.1 were highly significantly (p < 0.01) decreased after induced differentiation (Fig. 3G). To further reveal the potential relationship between candidate lncRNAs and the MSTN and/or FGF5 genes, the lncRNA interference experiments were performed. Our results showed that knockdown of MSTRG.27522.1 significantly (p < 0.05) inhibited the expression of MSTN gene and promoted the expression of FGF5 gene (Fig. 3H). Downregulated MSTRG.33818.1 also significantly (p < 0.05) inhibited the expression of MSTN gene, but had no significant (p > 0.05) effect on FGF5 gene (Fig. 3I). Taken together, the candidate lncRNAs were recognized as potential gatekeepers for the muscle phenotype of MF+/− sheep.
Discussion
lncRNAs are widely involved in many important BPs, which makes it attract much attention (Kopp and Mendell, 2018; Weikard et al, 2017; Zhao et al, 2019). It seems that lncRNAs appear to be emerging as gatekeepers of skeletal muscle development. However, most studies of lncRNAs have focused on humans and mice (Cabili et al, 2011). In fact, although numerous lncRNAs have been identified in sheep skeletal muscle (Chao et al, 2018; Li et al, 2019;Li et al, 2018a; Ren et al, 2017), only a rare of lncRNAs have been implicated as a molecular switch in sheep muscle development (Bakhtiarizadeh et al, 2016), especially in MSTN gene-edited sheep. For example, the NONCODE database is dedicated to ncRNAs of 17 species, including humans, mice, bovines, rats, chickens, and pigs.
However, this database lacks information on sheep lncRNAs (Zhao et al, 2016). In the present study, we comprehensively characterized skeletal muscle related-lncRNAs between WT and MF+/− sheep, as well as their potential regulatory roles in muscle development.
Using a strict filtering pipeline, we identified a total of 8265 lncRNAs, including 4455 novel lncRNAs transcripts and 3810 known lncRNAs, which expanded the available catalog of lncRNAs in the sheep genome. Subsequently, a total of 280 DELs between WT and MF+/− sheep were identified, which included 78 upregulated DELs and 202 downregulated DELs. However, the functions of a large number of lncRNAs are a mystery, especially those of the novel lncRNAs. In general, the function of lncRNAs is inferred by predicting its target genes. A well-known function of lncRNAs is to regulate gene expression, as they can directly regulate RNA polymerase II or promote transcription factor phosphorylation and affect their DNA-binding activity (Sadallah et al, 2011; Zhang et al, 2018). Numerous studies have shown that lncRNAs preferentially regulate their neighboring protein-coding genes in a cis-acting manner (Jandura and Krause, 2017).
Thus, the cis-target of lncRNAs can be inferred from their physical location relative to mRNAs. In addition, lncRNAs can also regulate protein-coding genes far away from their transcriptional sites, and this position-independent regulation is referred to as trans-acting, which can be predicted by analyzing mRNAs that are coexpressed with the lncRNAs. In this study, a total of 150 differentially expressed cis-target mRNAs were identified, all of which were less than 100 kb away from the lncRNAs. In addition, the coexpression analysis of DELs and DEGs identified a total of 196 differentially expressed trans-target mRNAs. By combining the cis- and trans-targets, a total of 256 PTGs were identified, of which 86 PTGs displayed both cis and trans interactions with lncRNAs.
Functional enrichment revealed that DELs were significantly associated with muscle contraction, calcium ion transport, and skeletal muscle cell differentiation, as well as apoptosis, autophagy, and skeletal muscle inflammation. Among these PTGs, we focused on enrichment entries that are closely related to muscle development, and BTG2, MYC, ATF3, CXCL10, MSTN, and ANKRD2 were significantly enriched for myogenic differentiation. The BTG2 and its homolog BTG3 were mapped to porcine chromosomes, and their expression levels in the longissimus dorci muscle were different between the Landrace and Chinese Tongcheng pig breeds (Feng et al, 2007). Further investigation showed that both BTG2 and BTG3 were induced in differentiated C2C12 cells, suggesting a role for them in myogenic differentiation (Feng et al, 2007).
A recent study also demonstrated that miR-222-3p can regulate the proliferation and differentiation of C2C12 myoblasts by targeting BTG2 (Yang et al, 2019). ATF3 plays a role in promoting the survival of motor neurons and maintaining muscle innervation, thereby improving the motor ability of ALS (Seijffers et al, 2014). CXCL10, a chemokine of the CXC subfamily, promotes the proliferation of muscle satellite cells via CXCR3 (Zhang et al, 2020). Recombinant CXCL10 treatment rejuvenates muscle satellite cell functions and restores muscle regeneration in aged mice (Zhang et al, 2020). In addition, the muscle-specific putative myogenic regulator ANKRD2 can be controlled by MyoD during myogenic differentiation (Bean et al, 2005). These results suggest that lncRNAs targeting these PTGs may be important regulators of myogenic differentiation and muscle formation. The information of these lncRNAs and their PTGs are listed in Table 1.
Summary of Key Interesting Candidate Long Noncoding RNAs and Their Potential Target Genes Closely Related to Muscle Development
DEL, differentially expressed lncRNA.
In addition, the differentially expressed PTGs AVPR1A, HES6, and ZFP36 had aroused our great interest. Arginine vasopressin receptor 1A (AVPR1A) is the most abundant receptor in all arterial vasculature (Liu et al, 2020), and vasopressin increases blood pressure by occupying AVPR1A receptors on vascular smooth muscle (Russell and Walley, 2010), these evidence highlights the important role of AVPR1A in myocardial contraction. HES6 is an important regulator of myogenesis, the constitutive expression of HES6 in myoblasts can inhibit the expression of myogenesis repressor MyoR and induced myogenic differentiation (Gao et al, 2001). Reciprocally, blocking endogenous HES6 function by using a WRPW-deleted dominant negative HES6 mutant resulted in a increased MyoR levels and completely blocked the muscle development program (Gao et al, 2001).
More interestingly, the mRNA decay activating protein ZFP36 plays a key role in maintaining the quiescence of skeletal muscle satellite cells by promoting ARE-mediated mRNA decay of myogenic differentiation MyoD1 mRNA (Bye et al, 2018; Hausburg et al, 2015). In addition, ZFP36 also regulates the early adipogenesis of preadipocytes by promoting the mRNA decay of ARE mediated immediate early genes (Lin et al, 2012). Altogether, the lncRNAs MSTRG.49654.1, MSTRG.33818.1, MSTRG.27522.1, and MSTRG.30947.1 were recognized as potential key gatekeepers of muscle phenotype in MF+/− sheep due to the role of candidate lncRNAs PTGs in myogenic differentiation and skeletal muscle development and the significant differential expression at the tissue and cellular levels, especially before and after myogenic differentiation. The information related to these lncRNAs and their PTGs are listed in Table 1.
Conclusion
In this study, we identified the expression profiles of lncRNAs in WT and MF+/− sheep, and then screened out the key candidate lncRNAs that can regulate myogenic differentiation and skeletal muscle development. These key candidate lncRNAs can serve as critical gatekeepers for muscle contraction, calcium ion transport, skeletal muscle cell differentiation, apoptosis, autophagy, and skeletal muscle inflammation, further revealing that lncRNAs play crucial roles in regulating muscle phenotype in MF+/− sheep. In conclusion, our newly identified lncRNAs may emerge as novel molecules for muscle development and provide a new reference for MSTN to regulate skeletal muscle development.
Data Availability Statement
The datasets presented in this study can be found in online repositories (
Ethics Statement
All experiments were performed in accordance with relevant guidelines and adhere to the ARRIVE guidelines (
Footnotes
Disclosure Statement
No competing financial interests exist.
Funding Information
This work was supported by National Key Research and Development Project of China (2021YFF1000704), Natural Science Foundation of China (32072722), and National Transgenic Creature Breeding Grand Project (2016zx08008-003).
Supplementary Material
Supplementary Table S1
Supplementary Table S2
References
Supplementary Material
Please find the following supplemental material available below.
For Open Access articles published under a Creative Commons License, all supplemental material carries the same license as the article it is associated with.
For non-Open Access articles published, all supplemental material carries a non-exclusive license, and permission requests for re-use of supplemental material or any part of supplemental material shall be sent directly to the copyright owner as specified in the copyright notice associated with the article.