
Abstract
This article is focused on the main pathways used by viruses to achieve infection and lysis of unicellular eukaryotes described as pathogenic for multicellular organisms. In light of the recent discussions on how tumor cells exhibit unicellular behavior, highly malignant cells can be considered as another unicellular pathogenic entity, but with endogenous origin. Thus, a comparative panel of viral lysis of exogenous pathogenic unicellular eukaryotes such as Acanthamoeba sp., yeast, and tumors is presented. The important intracellular parasite Leishmania sp is also presented, which, in contrast, has its virulence improved by viral infections. The possible exploitation of viral-mediated eukaryotic cell lysis to overcome infections of Leishmania sp is discussed.
Introduction
Molecular biology started with the study of the relationship between viruses and prokaryotes, namely bacteriophages and the bacterium Escherichia coli, in the early stages of genetics development. Using this experimental model, it was possible to elucidate key features that are shared by other virus–cell interactions: the virus enters the cell and replicates using bacterial cell machinery. Some viruses can exhibit two alternative outcomes: the lytic path, where viral particles are built leading to cell lysis; and the lysogenic path, where the viral genome is integrated into the bacterial genome and it is vertically transmitted to the progeny (Clokie et al., 2011).
It was not long before these characteristics, which were naturally present between viruses and bacteria, offered an opportunity for the development of molecular biology techniques, which allowed the study of genomes through cloning and gene sequencing (van Holde and Zlatanova, 2018). In addition, it has become a therapeutic alternative for the treatment of bacterial infections. Therapy with lytic bacteriophages had its enthusiasm in the early 20th century, with positive results for the treatment of various bacterial infections, including cholera (Cisek et al., 2017).
After the discovery of antibiotics, studies with bacteriophages mostly ceased. With the emergence of bacterial infections resistant to multiple drugs, treatment of these infections has become a challenge. According to a report by the U.S. Centers for Disease Control and Prevention (CDC), more than 2.8 million antibiotic-resistant infections occur in the United States each year, with more than 35,000 deaths as a result (CDC, 2019).
Given these numbers, the scientific community has turned to the use of phages as an alternative for the treatment of multiresistant infections (Abd El-Aziz et al., 2019; Pallavali et al., 2017). Thus, the search for phage strains with strong lytic activity to overcome resistant bacterial infections has received increased attention (Jeon et al., 2019; Yang et al., 2019).
Viruses are part of all life forms, and the relationships established between viruses and eukaryotes also offer opportunities for the development of technologies and methods aimed the treat and reversing therapeutic resistance to chemotherapy in various pathological conditions induced by unicellular eukaryotes. The relationship between viruses and eukaryotes has elements as vast as they are complex, it has been shaped over an evolutionary period and has had a strong impact on the composition and dynamics of modern eukaryotic genomes (Forterre, 2006). Even though several biotechnological tools and treatments have emerged from these studies, the potential adaptation of these techniques was not explored toward neglected diseases, namely those caused by Leishmania species. Herein, we will address some of the most potentially useful virus–eukaryotic cell interactions that are and could be, explored for therapeutic purposes, with an emphasis on Leishmaniasis.
Leishmaniasis and virus–Leishmania interactions
Leishmaniasis
Leishmaniasis is caused by obligate intracellular protozoa from the Leishmania genus. Its life cycle is divided into two phases: Promastigotes, the flagellate form found in the sandfly vector, which is delivered to the skin, and further to the bloodstream of a vertebrate host after the sandfly bite (Nakkash-Chmaisse et al., 2011). Once inside the host, the flagellate form is phagocyted by macrophages, and in structures known as phagolysosomes, promastigotes lose their flagella, transform into amastigote, and proliferate (McConville and Naderer, 2011). Eventually, new amastigotes emerge from lysed macrophages and infect more macrophages. When the infected vertebrate is bitten by a sandfly, the amastigotes are taken up, and inside the gut of the sandfly, it differentiates into infective promastigotes, ready to infect the next vertebrate host (Tsigankov et al., 2014) (Fig. 1A).

Leishmania life cycle and LRV1 propagation.
The clinical manifestations of Leishmaniasis include cutaneous, mucocutaneous, and fatal visceral disease. The disease pathogenesis is derived from strong immunological responses against the parasite, with the production of inflammatory cascade signaling molecules leading to the recruitment of neutrophils, monocytes, and activated T cells, which contributes to lesion formation. Whereas cutaneous Leishmaniasis is the most common, Visceral Leishmaniasis is the deadliest; and mucocutaneous can be disfiguring, causing social stigmatization and patient´ psychological suffering (Kassi et al., 2008).
Treatment of Leishmaniasis involves the use of three main compounds: pentavalent antimonial, Amphotericin B, and Miltefosine, but other drugs with different levels of efficiency are also used. Pentavalent antimonial (Sbv) constitutes the first-line treatment against Leishmaniasis, with sodium stibogluconate and meglumine antimoniate. The several problems regarding antimonial use are long courses of administration, high toxicity, and cost, among other disadvantages, the efficiency of the treatment has been reduced due to drug resistance (Roatt et al., 2020). Pentavalent antimonials have been in use since the 1940s to treat Leishmaniasis (Marsden, 1985).
Used as the second-line treatment for Leishmaniasis, Amphotericin B deoxycholate is a broad-spectrum antifungal that binds to membrane sterols, forming ion channels and increasing membrane permeability (de Carvalho et al., 2013). Side effects such as fever, nausea, and vomiting among others limit its use (Sundar et al., 2007). Miltefosine is an alkyl phosphocholine compound used in the treatment of several unrelated conditions as solid tumors, Schistosomiasis, and visceral Leishmaniasis (Dorlo et al., 2012; Eissa et al., 2015), with side effects such as diarrhea, hepatotoxicity, and thrombophlebitis among others. Furthermore, it is also teratogenic (Sundar and Chakravarty, 2015).
Owing to drug resistance to the available therapies, the strong side effects, and limited use in elderly, children, and pregnant women, alternative approaches have been tested as the development of liposome- and cyclodextrin-based formulations to improve drug bioavailability (Frézard et al., 2009), and nanoformulations using polymeric nanoparticles, among other formulations, has improved drug efficiency in vivo (Akbari et al., 2017; de Carvalho et al., 2013).
With so many approaches under development, Leishmaniasis is still an important world health issue, affecting more than 1 million people, mostly in developing countries (WHO, 2023 ).
Virus–Leishmania interactions
The viral life cycle involves receptor-dependent attachment to the target cells' surface, penetration through membrane fusion (enveloped) and receptor-mediated endocytosis; intracellular trafficking, uncoating, viral gene expression, and genome replication; capsid assembly, and release. Most enveloped viruses are released extracellularly through exocytosis, where the virus progeny is generated and constantly released from the host cell without cell death or lysis resulting in chronic infection, whereas naked viruses are mostly released by cell lysis (Ryu, 2017). Although there are viruses capable to infect new hosts after release to the extracellular medium, others are noninfective, capable of only vertical transmission, and are considered endosymbionts (Lafleur and Olivier, 2022).
Some strains of Leishmania sp are constantly infected with Leishmania RNA Virus (LRV) (Molyneux, 1974), which belongs to the Totiviridae family, and is characterized by nonenveloped, 40 nm icosahedral symmetry organized particles, built from 120 copies of capsid protein with linear 5,283-nucleotide-long double-stranded RNA (dsRNA) genomes (Procházková et al., 2021; Widmer et al., 1989). They possess a capsid-RNA-dependent RNA polymerase, necessary for its replication (Stuart et al., 1992).
After phylogenetic studies, LRV types were divided into LRV1 and LRV2, with LRV1 infecting Leishmania of the New World and LRV2 being detected in Old World Leishmania (Ginouvès et al., 2016; Zabala-Peñafiel et al., 2022). It was extensively documented that some protists are permanently infected with representatives of the Totiviridae, with no external transmission pathway. This is also true for Leishmania species, where the virus is spread through vertical transmission, with no virus-free infection by an extracellular virus (Lafleur and Olivier, 2022).
The presence of the LRV1 in Leishmania species may induce more aggressive disease progression. Some studies have reported that LRV1 acts as a potent innate immunogen, leading to a hyperinflammatory immune response through toll-like receptor 3 (TLR3), leading to enhanced virulence of Leishmania (Ives et al., 2011). However, the frequency of LRV1 present in Leishmania sp in some regions is very low (Macedo et al., 2016), and even, the severity of the disease was not always correlated to the presence of the LRV1 infection (Pereira Lde et al., 2013).
Recently, a stunning study reported a possible pathway for horizontal transmission of the LRV1 through exosome in L. Viannia panamensis and L. mexicana. This study showed that Leishmania exosomes function as viral envelopes, thereby facilitating LRV1 transmission and increasing infectivity in the mammalian host. They conclude that exosome-containing LRV1 is secreted to the extracellular environment and that these exosomes were able to infect virus-free promastigotes in vitro, but this infection was sustained only for L. Viannia panamensis. Furthermore, it was shown that naked viruses that are extruded from the flagellar pocket are unable to infect Leishmania in vitro (Atayde et al., 2019) (Fig. 1B).
The natural infections by LRV1 lead to the release of immunogenic particles containing the virus without impairment of the parasite viability. Several hosts for some viruses can be revealed through in vitro cocultures. Observing other known interactions between viruses and other eukaryotes could provide some candidates.
Virus–pathogenic eukaryotic cell interactions leading to lytic cycle and/or cell death
Cell lysis of malignant human cells (viral oncolysis)
Viral oncolysis is the lysis of a tumor cell-mediated by viruses that infect and replicate inside them. The use of viruses to treat tumors came from the observation that some cancer patients went into temporary remission when naturally infected by viruses (Kelly and Russell, 2007). Initially, human pathogens (Hepatitis B virus) were used to treat Hodgkin's disease, leading to the death of some patients (Hoster et al., 1949). To avoid the virulence of human viruses, animal viruses were also tested, with promising results (Cassel and Garret, 1965), and with the advent of genetic engineering, modified versions of viruses with more specificity toward cancer cells, with mild side effects are under clinical trials (Alemany et al., 2000).
Considering the recent discussions on how tumor cells exhibit unicellular behavior (Fernandes et al., 2012; Solé et al., 2014), highly malignant cells can be regarded as another unicellular pathogenic entity, but with endogenous origin. The growing malignancy leads tumor cells to a unicellular behavior (Chen et al., 2015), where the cell uses several molecular defense mechanisms that are extremely suitable to the unicellular condition, including multidrug resistance, radioresistance, and enhanced mobility, features that allow tumor cells to detach from multicellular structure (Fernandes et al., 2012). Even though several strategies are available to treat tumors, resistance to both conventional and target-directed therapy maintains the high rates of cancer deaths (WHO, 2022).
Among several important features of resistant tumors are the overexpression of genes able to reduce cell sensitivity to drug treatment, leading to multidrug resistance (Emran et al., 2022) and mutation of the targets in target-directed therapy (Braun et al., 2020). Some of these are oncogenes, such as the Kras, a homolog of the Kirsten murine sarcoma virus whose overexpression is involved in the reduction of activity of the antiviral defense system dependent on type I interferon (IFN) response activation (Christian et al., 2012).
These interactions were already elucidated and were shown that overexpression of proteins related to tumor resistance helps to disable some defenses against oncolytic virus infections (Fernandes, 2016). This includes the dsRNA-activated protein kinase (PKR) antiviral defense system. In normal cells, PKR is activated by the presence of dsRNA through activation of type I IFN response activation, leading to phosphorylation of elF2α, responsible to initiate translation of the RNA, impairing cell translation. In contrast, in tumor cells, PKR is inactivated or downregulated by tumor progression factors. This points some viruses toward tumor cells, instead of normal cells (Fernandes et al., 2016).
These observations have launched viral anticancer therapy with promising clinical results for glioma (Chiocca and Lamfers, 2003), liver (Park et al., 2008), and prostate cancer (Taguchi et al., 2017). Reoviruses and adenoviruses (Pesonen et al., 2011) are the most common oncolytic viruses under study, but oncolysis induced by Myxoma virus (Lun et al., 2010), influenza (Muster et al., 2004) and Herpes simplex (HSV) (Fasullo et al., 2009). Among the desirable features of the most successful oncolytic viruses under development is the induction of significant tumor cell lysis, stimulation of antitumor immune response, and mild side effects.
In addition to the use of viral oncolysis as monotherapy, the combination of these viruses with conventional and target therapy has strong improved oncolysis potential (Porosnicu et al., 2022). In 2015, the first Oncolytic Virus Therapy was approved by FDA. Talimogene laherparepvec (TVEC), based on a modified HSV virus (HSV) type 1, was approved to treat metastatic melanoma with strong results in patients (Andtbacka et al., 2015). The association of TVEC with immune checkpoint inhibitors can lead to side effects such as Vitiligo (Iglesias et al., 2019), and Disseminated Herpes Infection (Kimmis et al., 2022), which has rare (Glas et al., 2012) (or no) lethality compared with unresectable melanoma.
The modulation of virulence factors is an important approach to allow the use of some viruses with strong oncolytic activity that are potentially harmful to patients. This is the case of vesicular stomatitis virus (VSV), which needs manipulation due to its natural neurotoxicity (Lin et al., 2020).
Oncolytic therapy is one successful example of how virus–cell natural interactions were biotechnologically explored leading to a prolific field of knowledge and advancement of medicine with clinical results.
Virus-induced cell death of yeast (two-step killer toxins)
The genus Candida constitutes a group of yeasts that cause opportunistic infections in humans, with Candida albicans being the most common species identified in human infections. Candidiasis can affect the skin, gastrointestinal and respiratory mucosa, and urogenital tract. They become pathogenic in patients with compromised immune systems, leading to superficial, invasive, and systemic infection; the latter may lead to patients' death (Wisplinghoff et al., 2004). In addition to the immunocompromised people, risk groups include people with chronic diseases such as diabetes, to patients under corticosteroids and antibiotic treatments, among others (Fukushima et al., 2005).
Candida, so as other yeast, can be susceptible to killer toxins (Giovati et al., 2018). These are cytotoxic proteins that may be encoded by yeast chromosomes, dsDNA plasmids, or dsRNA viruses (Mannazu et al., 2019). These toxins are expressed as an unprocessed precursor and are secreted after post-transcriptional processing (Riffer et al., 2002). Three different killer toxins (K1, K2, and K28) were identified in Saccharomyces cerevisiae, each one encoded by a different killer virus (ScV-M1, ScV-M2, and ScV-M28) from the Totiviridae family (Ramírez et al., 2017).
In this phenomenon, toxins kill susceptible strains in a two-step mode. In addition to the killer virus, a helper virus L-A is necessary to generate the killer phenotype (Schmitt and Breinig, 2006). Both the killer and the helper viruses are dsRNA viruses. The two-step mode involves the activation of a first receptor located on the yeast cell wall and a second receptor located on the plasma membrane. Initially, the killer toxin (e.g., K1) binds to a receptor (β-1,6-
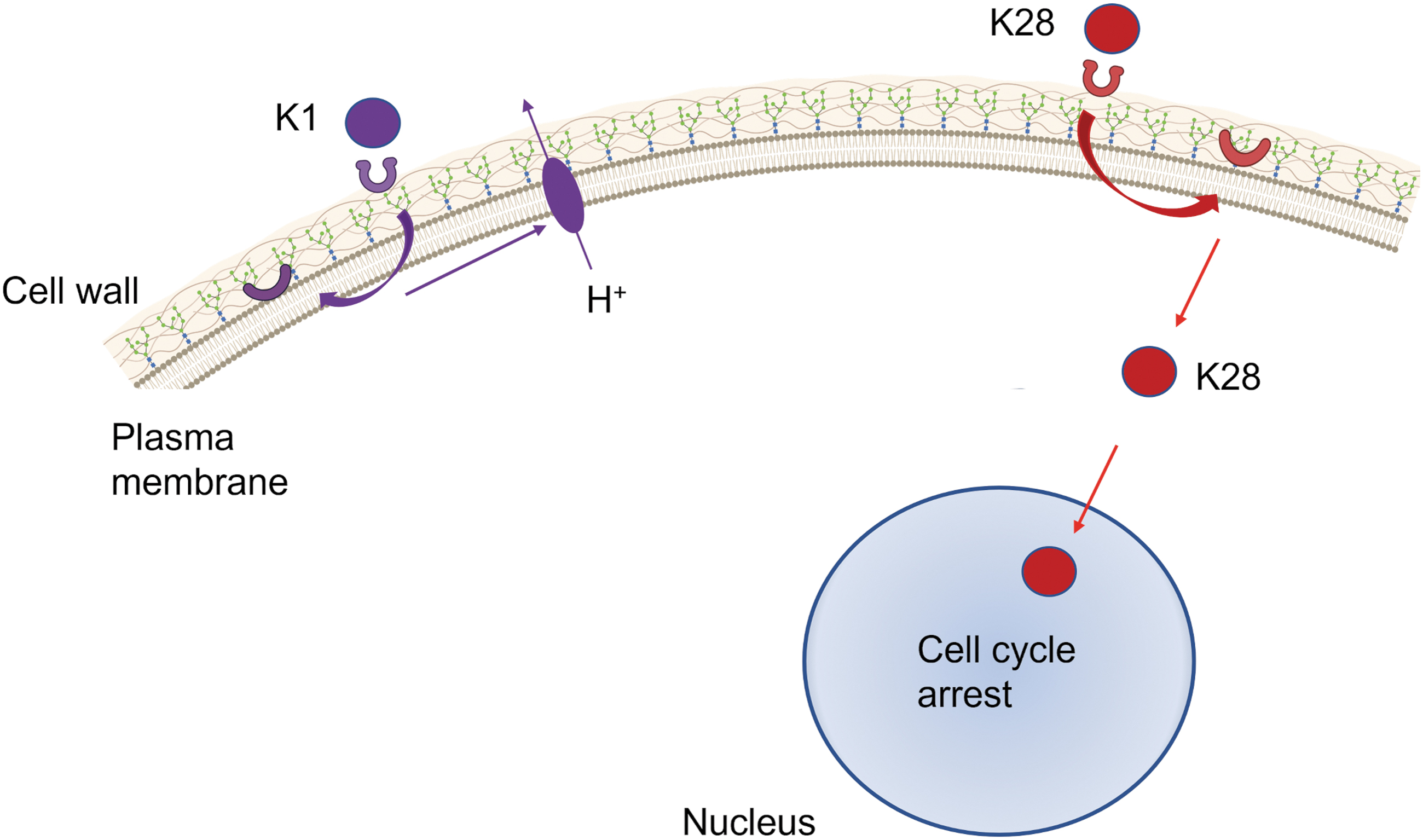
Virus-encoded killer toxins general mode of action. The first step in K1 mode of action (in purple) is the interaction with the first receptor (β-1,6-
A different dynamic is shown by K28, where the viral killer toxin enters a sensitive yeast cell by endocytosis, reaching the nucleus through a reverse secretion pathway (through Golgi and ER), where the toxin mediates cell cycle arrest and DNA synthesis inhibition inducing cell death. An interesting feature of all three toxins is that the toxin-producing strain is immune to its toxin, but not to other strains' toxins (Becker and Schimit, 2017).
The yeast strains infected with viruses that encode these toxins are thought to possess the “killer phenotype” and are already used for biotechnological purposes to kill contaminating wild-type yeasts in the food and fermentation industries (Mannazu et al., 2019).
Two-step killer toxins for Leishmania spp.
The in vitro tests on killer toxins to reduce the viability of Leishmania sp were already performed. A monoclonal antibody (KTmAb) was developed through idiotypic vaccination using a yeast killer toxin produced by the Pichia anomala. This study showed that the in vitro leishmanicidal activity against Leishmania infantum and Leishmania amazonensis promastigotes was dose dependent, reaching 30% of viability reduction (Savoia et al., 2002). Following this study, this group tested a synthetic killer decapeptide that mimetics the yeast killer toxins treating Leishmania promastigotes. Even though the viability decreased, no apoptosis was detected. The promastigotes exhibited extensive morphological alterations, such as parasite ghosts, swelling of the mitochondrion–kinetoplast complex, and changes in the organization of the nuclear and kinetoplast chromatins (Savoia et al., 2006).
Since Leishmania sp. is an intracellular parasite, the host membrane characterizes an additional challenge to any approach using antibodies/peptides, as the two-step yeast killer toxins. Combining oncolytic viruses armed with killer toxins to kill the intracellular parasite could improve its efficiency. In fact, its medical use has been discussed (Giovati et al., 2021), as other yeast derivatives, as yeast membrane vectorization (glucan particles derived from the yeast cell wall) (Scariot et al., 2019), to kill intracellular parasites (amastigote). The number of membranes is critical to define a strategy to treat cells infected with amastigote, as Leishmania lipid and protein membrane composition (Oliveira et al., 2020).
Giant virus-induced lysis of Acanthamoeba castellanii and other protists
Acanthamoeba is a unicellular eukaryote with ubiquitous distribution. Its life cycle is divided into two stages: cyst and trophozoite. In the cyst stage, Acanthamoeba acquires a cell wall and became metabolically inactive, whereas in the trophozoite stage this organism exhibits vegetative growth with metabolic activity (Siddiqui and Khan, 2012). Acanthamoeba can be found in a wide range of water sources (fresh water, seawater, etc.), anywhere these sources are used (air-conditioning units, ventilation ducts, swimming pools, etc.) (Marciano-Cabral et al., 2003), and in tissue samples from humans and other animals (Newsome et al., 1992).
Among several pathological conditions, Acanthamoeba may cause granulomatous amoebic encephalitis (Marciano-Cabral et al., 2003), which, even though rare, may lead to patient death (Ringsted et al., 1976); and keratitis, an inflammation of the cornea that can lead to blindness. Acanthamoeba is the second most isolated microbe among patients who wear contact lenses (Booton et al., 2002).
A case report on a patient with keratitis associated with amoeba infection showed that this amoeba was infected with A. polyphaga mimivirus (Cohen et al., 2011). This virus belongs to the Megavirales group and possesses an icosaedral capsid surrounded by a lipid membrane, and a dsDNA genome (Colson et al., 2017) with 1.2 Mbp in length (Raoult et al., 2004).
The invasion of amoeba by mimivirus occurs through phagocytosis, and once inside the phagosome, a special viral structure, the vertex (also called “the stargate”), allows the fusion of the internal viral membrane with the membrane of the host phagosome, releasing viral genome into host cytoplasm (Zauberman et al., 2008), where the virus replicates and generates a large and elaborate viral factory as a result of the coalescence of smaller structures, and the viral proteins are translated, leading to virion assembly, host lysis, and viral progeny release (Oliveira et al., 2020; Zauberman et al., 2008); however, not all mimiviruses induce host cell lysis. Other confirmed hosts for the A. polyphaga mimivirus include Vermamoeba vermiformis (Geballa-Koukoulas et al., 2022), zooflagellates (Fischer et al., 2010), and algae as Chlorella (Sheng et al., 2022).
Organisms that are the hosts of several of these giant viruses were identified using laboratory cocultures (Schulz et al., 2022), with some of these virus–host interactions not found yet in nature. Of particular interest for this discussion, Chlorella sp can live as free-living algae, so as an endosymbiotic of Paramecium bursaria (Kodama and Fujishima, 2022), this organism can also be infected by other giant viruses such as Chlorella Virus XW01 (Sheng et al., 2022) and the giant virus Paramecium bursaria Chlorella virus-1 (PBCV-1), inducing chlorella cell lysis (Clasen and Elser, 2007). Most studies involving chlorella infection were performed in chlorella cultures in vitro, but it was not reported chlorella infection by a large virus while in endosymbiotic condition, nor the fate of paramecium when populated with chlorella infected by any of the viruses it hosts.
Given the natural tropism of giant viruses toward free-living protists (Schulz et al., 2020), so as endosymbiont protists (Kodama and Fujishima, 2022) and the strong lytic outcome due to these viruses' infection, giant viruses could be an interesting candidate to be studied regarding its potential to infect intracellular protozoa, as Leishmania species.
Infection of Leishmania by lytic viruses—a matter of membranes
The use of viral-induced lysis for the treatment of neglected parasitic diseases has finally been discussed. Barrow et al. (2020) presented a discussion on the protozoal viruses pointing out the family Totiviridae, and the potential to treat Leishmaniasis, Giardia, and Candida. The authors also explore the molecular genetics of this viral family and how some molecules produced by viral genomes of this family could be exploited for antiprotozoal activity using technologies such as viral-like particles as drug delivery systems and protozoan virus modification (Barrow et al., 2020).
As observed in early sections, several interactions between viruses and their hosts were not found in nature; otherwise, they were established through laboratory cocultures. To adapt these approaches, the acknowledgment of the parasite is essential, as the observation of the main common features shared by the parasite and viral candidates. The lipid and protein composition of parasite membranes in their different forms (promastigotes/amastigotes) has been investigated for some Leishmania species (Bouazizi-Ben Messaoud et al., 2017; Brotherton et al., 2012). In addition, parasite intracellular dynamics such as amastigote proliferation and differentiation offer interesting opportunities.
Although extracellular promastigotes face condition as lower temperatures (26°C, sandfly gut temperature) and mild alkaline environment (7.0–7.5), Leishmania amastigotes multiply in macrophages, specifically within compartments that fuse with endosomes and lysosomes, generating the Parasitophorous Vacuole resembling mature phagolysosomes, with higher temperature (due to vertebrate host) among other features, and acidic environment (McConville and Naderer, 2011), with pH ranging from 4.7 to 5.5 during Leishmania differentiation (Antoine et al., 1990; Tsigankov et al., 2014). This acidification event is mediated by the vacuolar proton-ATPase, which is present on various endocytic organelles (Flannagan et al., 2009) (Fig. 3, left).

Left, pH range required for viral uncoating through the endosomal pathway. Right, macrophage phagolysosomes used by Leishmania sp. to achieve amastigote differentiation and proliferation.
Similarly, for some strains, virus uncoating is severely affected by pH (as Bunyaviruses, Filoviruses, and Rhabdoviruses) among others (Villanueva et al., 2005). After membrane-mediated endosomal pathway activation, the virus reaches the early endosomes, with mild acidity (pH 5.5–6.0), maintained by membrane-associated vacuolar-type ATPases. The endosome pathway progresses to the late endosomes, where most viruses with pH-dependent uncoating, achieve their invasion with a pH ranging from 5.0 to 5.5. That is the case of VSV (VSV, a widely studied oncolytic virus (Rigaut et al., 1991) (Fig. 3, right).
But studies have revealed that some giant viruses may need even lower pH levels. After being phagocyted, the vertex structure of a giant virus, responsible for nucleocapsid release and fusion with the phagosomal membrane, opens at the pH range as wide as 2.0–4.0 (Schrad et al., 2020) (Fig. 4).
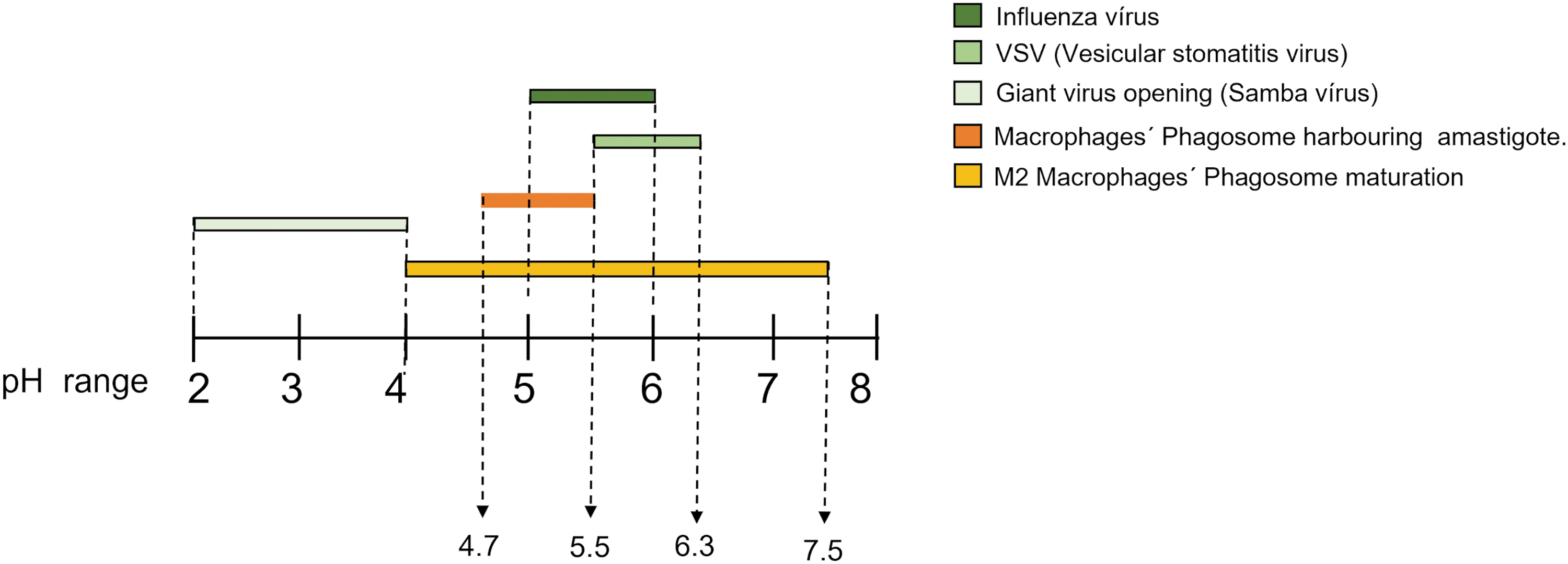
pH range of different viruses uncoating within endosomes and macrophage's vacuoles. Some of the data regarding the pH were observed only in vitro.
The recent discoveries about the viral exosomal pathway in Leishmania strains and its capacity to allow Leishmania viruses to infect other parasites could provide another possibility for lytic virus entry.
Conclusions
The question that imposes itself is if multidisciplinary could extend virotherapy to neglected parasitic diseases. The interactions of different viruses with different unicellular eukaryotes discussed so far provide interesting insights into the possibilities of the use of a suitable approach to deal with diseases such as Leishmaniasis. The transposition of elements from one system to act as a tool to another system was successfully achieved in CRISPR/cas9, discovered in bacteria, and adapted as a biotechnological tool for the manipulation of several life forms (Balderston et al., 2021).
Some challenges in using viral lysis for Leishmaniasis, involve, among other things (if possible), (1) finding a way to move forward an existing natural latent infection to a lytic one; (2) using a virus that could naturally infect the Leishmania sp. through in vitro coculture, or (3) genetically modify lytic viruses to infect the Leishmania sp, selecting those who can use the same endocytic pathway, and tolerate the similar conditions as the intracellular parasites. In particular, the generation of viral modifications in giant viruses with the potential to infect and forward Leishmania into the lytic cycle also depends on the generation of data regarding the function of several (still) orphan proteins of these viruses.
At the systemic level, since there are several types of Leishmaniasis (cutaneous, mucocutaneous, and visceral), the approach for each kind of Leishmaniasis should probably be different: cutaneous and mucocutaneous Leishmaniasis offers the lesions as external sites for intralesional injection, which was extremely successful in viral lysis of melanoma. However, this approach would not be suitable for visceral Leishmaniasis.
The fight against Leishmaniasis continues to have both technological and economic challenges since this disease affects mostly populations from developing countries, and the studies depend on heavy science budgets. Thus, for now, it seems that the neglected status of this disease will not change in the near future, but this should not impair our efforts in enhancing both the budget and the number of groups working on the subject.
Footnotes
Acknowledgment
The author thanks Graziele Silva for the assistance with figures.
Author's Contribution
J.F. designed and wrote the article.
Disclosure Statement
No competing financial interests exist.
Funding Information
This study was supported by FAPERJ—Fundação Carlos Chagas Filho de Amparo à Pesquisa do Rio de Janeiro (grant no. E-26/211.058/2021).