
Abstract
Patients with chronic obstructive pulmonary disease (COPD) have obstructed airflow through their lungs. Single nucleotide polymorphisms in matrix metalloproteinases (MMPs) genes and the risk of COPD have been the subject of numerous studies, with conflicting results. This investigation was conducted to determine whether the MMP7 (T>C) gene variant (rs10502001) was associated with an increased risk of COPD. A case–control study was conducted with 360 subjects (180 healthy controls and 180 COPD cases). The polymerase chain reaction (PCR)-restriction fragment length polymorphism method was used to genotype the SNP rs1050200. mRNA expression of MMP7 was performed using RT-PCR. The genotypic/allelic frequencies and carriage rates of rs10502001 (T>C) polymorphism were evaluated in 180 each of healthy controls and COPD cases. Cases have higher TC/CC genotype frequencies than controls. The “CC” genotype was found to be significantly associated with increased COPD risk (p = 0.016). The “C” allele frequency was higher in cases than in controls and showed significant association with COPD (p = 0.005). The carriage rate frequencies of T(−) and C(+) were significantly higher in cases than in controls (p = 0.031 and 0.047, respectively). MMP7 expression was significantly upregulated (p = 0.001) in COPD cases as compared with the controls. In addition, comparisons of MMP7 expression between the COPD cases with different genotypes showed that the “CC” genotype cases had significantly higher expression than those with “TT” genotype. The present findings showed statistically significant correlation of MMP7 (T>C) polymorphism and expression with COPD. Therefore, MMP7 responsible for degradation of elastin has been strongly linked to the progression of COPD.
Introduction
Chronic obstructive pulmonary disease (COPD) is an inflammatory lung disorder that causes pulmonary airway obstruction in affected individuals (Jarhyan et al., 2022). The airflow limitation is the key feature of COPD that is not completely reversible. The airflow restriction is caused by an abnormally narrow airway in the lungs that is exposed to toxic particles or gases (Serifoglu and Ulubay, 2019; Zafari et al., 2022). COPD develops due to cigarette smoking (active or passive smoking), dust exposure, chemical agents, and biomass fuel (Choi and Rhee, 2020).
These irritants have the potential to cause structural alterations and chronic inflammation in the lungs' smallest airways (Moldoveanu et al., 2009). Patients with COPD have pathological changes in their proximal airways, lung parenchyma, peripheral airways, and pulmonary vasculature (Karnati et al., 2021). Although smoking is the major risk factor that causes abnormal small airways, however, not all smokers develop COPD. Reports have indicated that the disease often affects middle-aged people and also suggested that hereditary factors cause the disease (Beghe et al., 2021). COPD is associated with numerous other medical problems, including heart disease, osteoporosis, lung cancer, musculoskeletal depression, and anxiety. COPD and other lung disorders cause epithelial damage and tissue healing or remodeling that require enzymes, proteins, and peptides (Karnati et al., 2021).
The macromolecules in the extra cellular matrix (ECM) are essential for development and matrix metalloproteinase (MMP), labeled as matrixins, degrade ECM (Cui et al., 2017). Vertebrates have 28 MMPs, however, only 23 are produced as preproenzymes in human tissues. Their architecture includes a variable-length N-terminal of signal peptide, latency-preserving prodomain, Zn2+ catalytic domain, a linker-“hinge” region, and C-terminal domain. Hemopexin-like domains at the C-terminus of all MMPs except MMP-7 and MMP-26 are necessary for certain of their activities (e.g., cleavage of collagen).
It is produced by healthy and noninflammed adult mucosal epithelial cells (Dufour et al., 2008). The majority of healthy and noninflammed mucosal epithelia produce MMP7. It affects defense, repair, and inflammation (Gharib et al., 2013). MMP-1, a collagenase that breaks down collagen, is strongly linked to COPD and may be a biomarker for smokers had significantly higher serum MMP-1 levels than COPD patients (Christopoulou et al., 2023). MMP-1 was significantly increased in smokers with and without progressive COPD.
MMP-1's functional variation rs1799750 G/GG also increased COPD risk (Sarra Bchir et al., 2016). MMP-8 expression levels were connected with acute dyspnea attacks in atopic COPD patients, suggesting it may be a clinical advisory tool (Christopoulou et al., 2023). Stable COPD patients have higher serum MMP-2 (gelatinase A) levels than asthmatics and controls, suggesting it may be a biomarker for COPD (Mahor et al., 2020). Single nucleotide polymorphisms (SNPs) in the MMP-12 promoter, like (-82) A/G allele of rs2276109/Asn357Ser (A/G) of rs652438, lower COPD risk and improve prognosis (Yang et al., 2020).
Human mucosal epithelium and macrophages express MMP7, or surface chemicals such as prodefensin, pro-TNF, Fas-ligand, and E-cadherin can be broken down by MMP7 (Cui et al., 2017). An SNP in the MMP-7 gene promoter (rs1156818) increases COPD risk (Christopoulou et al., 2023). Emphysema patients had higher MMP-7 serum levels and Global Initiative for Lung Disease (GOLD) stages (Tsay et al., 2020). MMPs degrade ECM and cell-surface proteins to control angiogenesis, wound healing, and immunity (Cui et al., 2017). Researchers have studied the link between SNPs in MMPs and the risk of COPD in different populations over the past 10 years.
MMP7 − 181A>G (rs11568818) is a functional A > G transition in the MMP7 gene promoter (Kesh et al., 2015). One study found that COPD patients exposed to biomass and cigarette smoke had higher serum MMP7 levels than healthy nonsmoking controls, which was negatively correlated with spirometry (Ortiz-Quintero et al., 2022). However, to the best of our knowledge, there is limited study showing the role of functional variations of MMP7 in the progression of COPD (Gharib et al., 2013).
In this regard, we planned to investigate the potential role of the MMP7 polymorphism and assessed the impact of SNP on the expression level of MMP7 as a risk determining factor to the onset of COPD in a north Indian population. This was done to determine whether or not the MMP7 polymorphism plays a role in the development of COPD.
Materials and Methods
Selection of study participants
The institutional ethics committee of King George's Medical University (KGMU), Lucknow, India, duly approved this study (94/Ethics/2021 dated February 02, 2021). All the subjects were between 35 and 70 years of age and both men and women were involved in the study with prior written consent. Controls (n = 180, age and gender matched) were selected from both universities, whereas COPD cases (n = 180) were from the outpatient department of respiratory medicine at KGMU, Lucknow. Spirometry was used to measure the lung capacities of COPD cases and controls, and all data were recorded before and after the administration of two asthilin puffs.
Collection of demographic and clinical data
Demographic variables of all subjects including age (years) and body mass index (BMI) were reported. The GOLD recommendations were used to determine the inclusion criteria for COPD cases (Adeloye et al., 2022). The criteria for COPD cases' inclusion in the study were those with postbronchodilator forced expiratory volume FEV1/forced vital capacity ≤70.0%, and postbronchodilator FEV1 ˂200 mL and ≤12% change in FEV1. COPD cases with comorbid conditions were excluded from the study. Controls were healthy individuals having postbronchodilator FEV1/FVC ≥80% and no family history of COPD, asthma, or any other specific disease (Supplementary Data).
Sample collection
EDTA vials were used to collect 2 mL of blood from each participant in the study for the purpose of DNA extraction. To extract RNA, 1 mL of blood was added to a vial that contains Trizol reagent (Life Technologies). Before any further usage, each sample was frozen at a temperature of −20°C.
DNA isolation and genotyping
The salting-out method was modified to isolate genomic DNA from whole blood (Miller et al., 1988; Sahu et al., 2021). The gene variant in MMP7 (rs10502001) was genotyped through the polymerase chain reaction (PCR) followed by the restriction fragment length polymorphism method. Primer 3.0 (ver online) was used for primer design, whereas NEB cutter (ver online) was used to determine the restriction enzyme. The materials used for PCR were 100 ng of template DNA, 1X Taq buffer, 10 pmol of forward and reverse primers (MMP7F: 5′-CCTGTAGGTGACCACTTTGGA-3′; MMP7R: 5′-AGCCAAACTCAAGGAGATGC-3′), 200 μM of dNTPs, 0.5 U of Taq polymerase (GeNei, Bangalore), and MilliQ water to make up the final volume.
The first denaturation of the DNA template occurred for 3 min. at 94°C. This was followed by 35 cycles of denaturation for 30 s at 94°C, annealing for 30 s at 53.6°C and extension for 30 s at 72°C. A last extension at 72°C for 5 min. brought the PCR process to an end.
The enzyme FauI (240 County Road, Ipswich, MA) was used to digest the PCR products after they were amplified as per instructions. The digested fragments revealing the genotypes were electrophoresed on 3% agarose gel. The Gel documentation system recorded the outcomes (Vilber Lourmat, France).
Expression of MMP7
Total RNA was isolated from the blood samples collected in Trizol reagent, and quantified in a biophotometer (Eppendorf, Germany). The RevertAid First Strand cDNA synthesis kit (Thermo Scientific) was used for cDNA synthesis. Real-time PCR (qPCR) was used to measure the expression of MMP7 with GAPDH as control (Roche Diagnostics, Germany). The reaction mixture (10 μL) contained 1X Maxima SYBR Green master mix (Thermo Scientific), 25 ng of cDNA, 2.5 pmol specific primers (MMP7F: 5′-GTTAAACTCCCGCGTCATAGAA-3′; MMP7R: 5′-GATCCTGTAGGTGACCACTTTG-3′), and MilliQ water.
The reaction conditions were set to 1 cycle at 95°C for 5 min, followed by 55 cycles at 95°C for 10 s, 57°C for 20 s, 72°C for 10 s, and a final hold at 10°C for 1 min. Expression analysis was done using the relative quantification method.
Statistical analysis
A t-test was used to compare demographic and clinical data of controls and patients. The data are shown as mean ± standard error. The strength of link between cases and controls using odds ratios (ORs) with 95% confidence intervals was calculated in GraphPad (ver.8.0). Each and every p-value was two-tailed, and a value of p < 0.05 was considered statistically significant.
Results
Clinical characteristics of case and control groups
In case of anthropometric parameters, the age of controls and cases did not differ. However, there were significant differences in BMI. Between the controls and cases, all spirometry parameters were found to be significantly different. As measured by spirometry, the lung capacity of the controls was greater than that of the cases (Table 1).
Details of Anthropometric Characteristics in Controls and Chronic Obstructive Pulmonary Disease Cases
Data shown as mean ± SD or number of subjects and analyzed by t-test. All differences p < 0.05 except age (p = 0.161). post-FVC: forced vital capacity after bronchodilator; post-FEV1: forced expiratory volume in 1 s after bronchodilator (statistical significance, * p < 0.05, ** p < 0.01).
BMI, body mass index; COPD, chronic obstructive pulmonary disease; FVC, forced vital capacity; SD, standard deviation.
Genotyping of MMP7 T > C (rs10502001)
The PCR products generated with specific primers for MMP7 T > C (rs10502001) polymorphism were 169 base pairs (bp). Fau1 digestion resulted in 169, 99, and 70 bp fragments, The “TT” genotype was represented by 169 bp, “TC” genotype by 169, 99, and 70 bp, and “CC” genotype by 99 and 70 bp (Fig. 1).

Agarose gel (3%) showing different genotypes of MMP7 T/C SNP (rs10502001). TT: 169 bp; TC: 169, 99, and 70 bp; CC: 99 and 70 bp; M: 50 bp Ladder. MMP, matrix metalloproteinase; SNP, single nucleotide polymorphism.
Association of MMP7 T > C (rs10502001) with COPD
The genotypic and allelic frequency distributions, as well as the carriage rates of the MMP7 polymorphism, were calculated in 180 healthy controls and 180 COPD cases. When compared with controls, the frequencies of “TC” and “CC” genotypes were higher in the cases (Table 2). The “CC” genotype was associated significantly with increased risk of COPD (p = 0.016) and a high OR of 2.25. The frequency of “C” allele was higher in cases than in controls and showed risk of COPD with significant association (p = 0.005). The carriage rate of C(+) and T(−) showed significant association with COPD when compared with controls (p = 0.047 and 0.031, respectively) (Table 2).
Genotypic, Allelic, and Carriage Rate Frequencies of MMP7 T/C Polymorphism (rs10502001) in Controls (n = 180) and Chronic Obstructive Pulmonary Disease Cases (n = 180)
Bold numerals show association with disease.
CI, confidence interval; MMP, matrix metalloproteinase; OR, odds ratio.
Expression of MMP7 and correlation with genotypes
The level of mRNA expression in whole blood cells of both controls and patients was measured by real-time PCR or quantitative PCR (qPCR). The relative mRNA expression of MMP7 gene was found to be significantly upregulated in cases when compared with that of controls (p = 0.001). This was determined by the fold change in expression (Fig. 2).
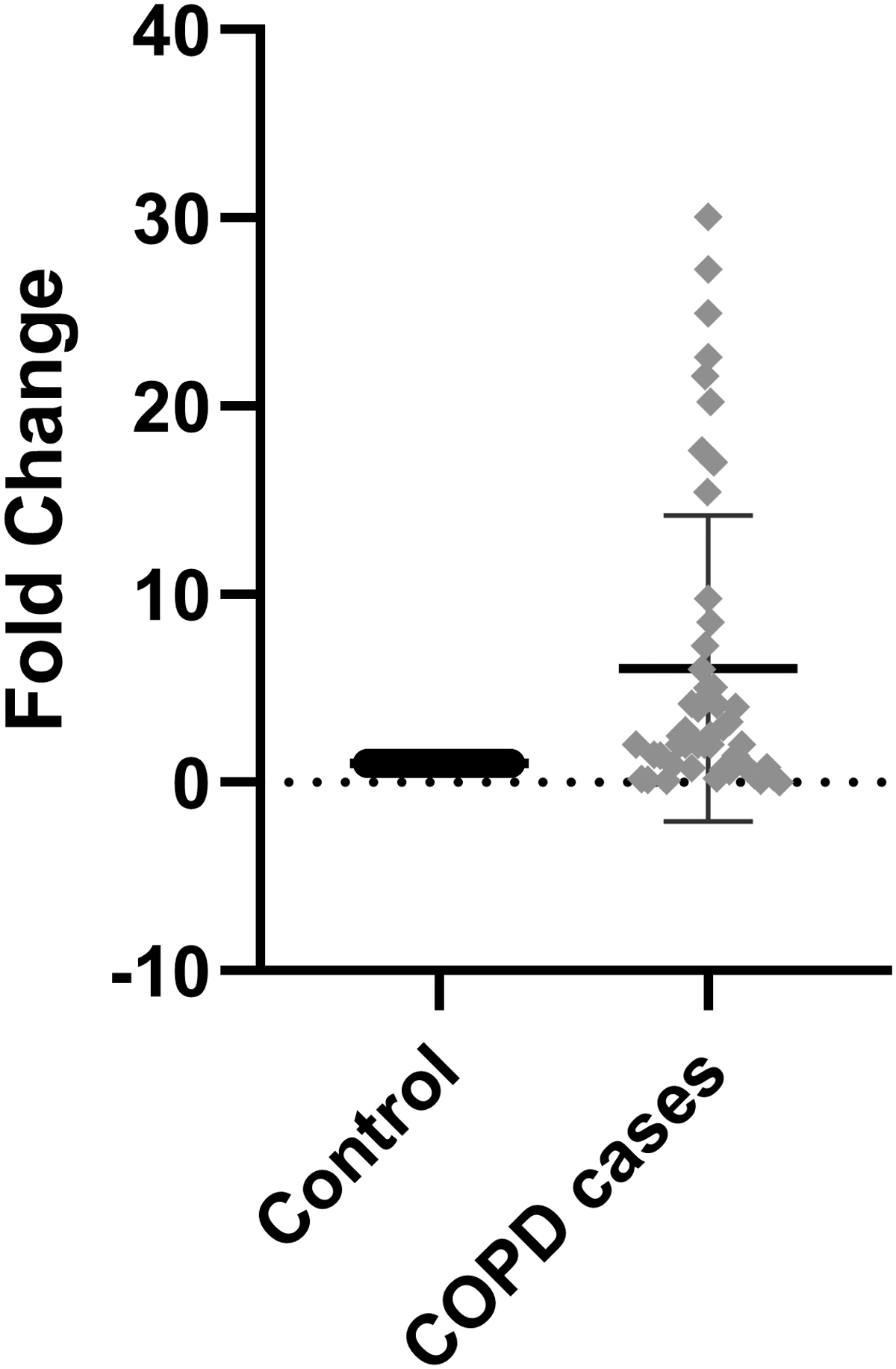
Expression of MMP7 gene in controls and COPD cases. COPD, chronic obstructive pulmonary disease.
Gene expression analysis in COPD cases carrying different genotypes of MMP7 T > C (rs10502001) polymorphism showed that individuals having “CC” genotype showed a significant increase in the mRNA level as compared with those with “TT” genotype (p = 0.0001), whereas “TC” genotype did not show any significant change (Fig. 3).
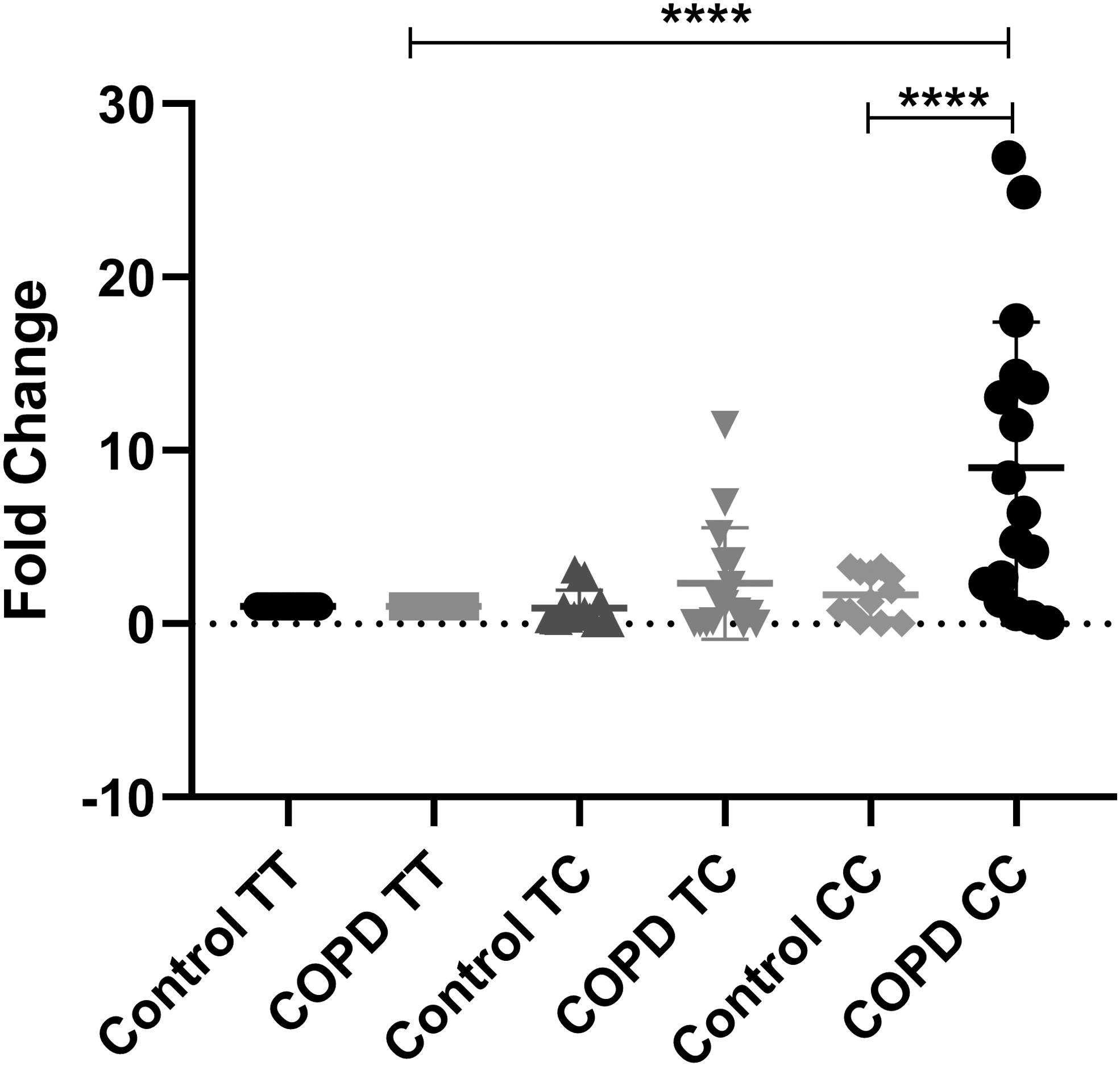
Gene expression in controls and COPD cases with different genotypes of MMP7 T/C variant (statistical significance, ****p < 0.0001).
Discussion
COPD is distinguished by significant tissue remodeling during the course of the disease (Wang et al., 2018), as well as the presence of chronic inflammation in the airways. MMPs are a group of zinc-containing enzymes that have proteolytic activity against a variety of extracellular proteins (Cabral-Pacheco et al., 2020). In normal physiological settings, MMPs transcription, activation of precursor zymogens, and inhibition by endogenous inhibitors are all closely regulated. As a result of their involvement, MMPs participate in a wide range of physiological and pathological processes in the body, such as development, involution, inflammation, tumor formation, and repair (Quintero-Fabian et al., 2019).
Several lines of evidence have revealed that MMPs are closely associated with the onset of COPD. For example, it has been reported that the serum levels of MMP2, MMP3, and MMP9 are upregulated in COPD (Gharib et al., 2013; Kraen et al., 2019). They play an important role in the turnover of nearly all ECM components.
It has been found that an A/G change in the MMP7 promoter may impact promoter activity since the G allele was found to dictate higher basal transcriptional activity in vitro in the human monocyte/macrophage cell line U937 (Sizemore et al., 2014). Those with the G allele (AG and GG genotypes) were found to be at a significantly higher risk of developing COPD at an earlier age than those with the AA genotype. Furthermore, possessing the G allele raises the chances of acquiring COPD before the age of 60 years by a ratio of 3. So yet, just one study has been published that analyzes the role of the MMP7-181A/G (rs11568818) gene variation in COPD (Hsieh et al., 2019). According to Mogulkoc et al. (2012), the MMP7 AA genotype is associated with an increased risk of developing COPD.
The identical variation has not been proven to be a risk factor for idiopathic pulmonary fibrosis, but it does modify the plasma level of MMP7 in patients (Tzouvelekis et al., 2017). Individuals with the AA genotype had a higher percentage than those with other genotypes. The MMP7-181A/G mutation has no association with the risk of developing lung cancer. However, it has been demonstrated that individuals who have received lung transplantation and have the AA genotype are more likely to develop bronchiolitis obliterans syndrome (Kesh et al., 2015).
Serum levels of MMP7, as well as other MMPs such as MMP1 and MMP9, are significantly greater in COPD patients who have been exposed to biomass and cigarette smoke than in healthy controls who have not been exposed to either of these factors. Furthermore, researchers (Lee et al., 2021) revealed that the amounts of those enzymes showed a negative relationship with lung function markers (FEV1% in 1 s).
MMP7 is involved in the breakdown of basement membranes caused by inflammatory cells (Cabral-Pacheco et al., 2020). MMP7 has been shown to act as a mediator during the progression of bleomycin-induced pulmonary fibrosis when E-cadherin is lost from the alveolar epithelium (Chulia-Peris et al., 2022). There was a statistically significant increase in the frequency of the “C“ allele in cases compared with controls. When compared with controls, the carriage rate frequencies of T(−) and C(+) were also considerably increased.
When COPD cases were compared with healthy controls, the level of MMP7 mRNA expression was found to be considerably higher in the COPD cases. Both “TC” and “CC” genotypes in COPD cases exhibited considerably increased mRNA levels as compared with corresponding genotypes in controls (“TT” genotype taken as reference). However, the association of MMP-7 expression with COPD cases was significant in “CC” genotype but not in “TC” individuals. When genotypes of controls were compared, there was no substantial difference in the relative mRNA expression of MMP7 gene. This effect could be ascribed to an increased synthesis of the MMP7 enzyme that degrades elastin.
It also leads to an accelerated rate of airway wall protein degradation and damage of ECM. Therefore, increased MMP7 has been strongly implicated in the development of COPD. According to the findings of our study, there is a strong correlation between the MMP7 T > C gene variant (rs10502001) and its higher expression in COPD cases and may be explored as a potential biomarker.
In conclusion, our finding shows that the MMP7 polymorphism (rs10502001) can affect the risk of COPD. Specifically, people who have both copies of the C allele (CC genotypes) might be more likely to get COPD early. In this regard, it is important to investigate whether or not there is a link between patients' genotypes and their serum levels of MMP7. This will help to determine whether or not the latter idea has merit. Overall, the data suggest that C allele carriers may be responsible for the disruption of ECM and cell–cell interactions, as well as the degradation of the tissue repair or fibrosis process, and hence the early onset of COPD.
Footnotes
Acknowledgments
The authors extend their gratitude to Indian Council of Medical Research (ICMR), New Delhi, Centre of Excellence and Research and Development Schemes, Higher Education, Government of Uttar Pradesh, Lucknow, India, for financial support. S.K. and A.S. thank ICMR and Council of Science and Industrial Research (CSIR), New Delhi, India, for senior research fellowships.
Ethics Approval and Participate Consent
The institutional ethics committee of King George's Medical University (KGMU), Lucknow, India, duly approved the current study (94/Ethics/2021 dated February 02, 2021). All the participants gave written consent of their participation in the study.
Availability of Data and Materials
Generated data or data analyzed during this study are involved in this published article.
Disclosure Statement
No competing financial interests exist.
Funding Information
This study was supported by ICMR (F.No.2017-3561/CMB-BMS), New Delhi, Centre of Excellence and Research and Development Schemes (No. 66/2019/1869/70-4-2019-4(24)/2019), Higher Education, Government of Uttar Pradesh, Lucknow, India.
Supplementary Material
Supplementary Data
References
Supplementary Material
Please find the following supplemental material available below.
For Open Access articles published under a Creative Commons License, all supplemental material carries the same license as the article it is associated with.
For non-Open Access articles published, all supplemental material carries a non-exclusive license, and permission requests for re-use of supplemental material or any part of supplemental material shall be sent directly to the copyright owner as specified in the copyright notice associated with the article.