
Abstract
Long noncoding RNAs (lncRNAs) have been implicated in coronary artery disease (CAD) processes. However, the relationship between the gene polymorphisms of lncRNA RP1-276N6.2 as a novel molecule and susceptibility to CAD remains unclear. In our case–control study, 949 CAD patients and 892 healthy controls were genotyped using the TaqMan genotyping assay. The quantitative real-time polymerase chain reaction and enzyme-linked immunosorbent assay were performed to examine the expression levels of RP1-276N6.2 and SLC22A3(OCT3). We observed that CAD patients had significantly lower RP1-276N6.2 levels than those healthy participants (p < 0.05). Compared to the wild-type genotype, the rs611950 T allele and the rs10499313 AG genotype and G allele significantly increased the premature CAD risk (p = 0.02, p = 0.002, and p = 0.01, respectively), while the rs505000 G allele reduced this risk (p = 0.01); moreover, the rs505000 CG genotype significantly enhanced the delayed CAD risk (p = 0.03), and the rs505000 G allele reduced the expression levels of RP1-276N6.2 and SLC22A3 (p < 0.05 and p < 0.05, respectively). In addition, RP1-276N6.2 positively regulated the mRNA and secreted protein levels of SLC22A3 (p < 0.05). In conclusion, the RP1-276N6.2 gene polymorphisms were closely associated with CAD risk. LncRNA RP1-276N6.2 may be a potential genetic target for CAD early diagnosis and treatment.
Introduction
Coronary artery disease (CAD) is the most common cardiovascular disease with lipid accumulation and chronic inflammation, which has become the leading cause of global death (Roth et al., 2020). The Annual Report on Cardiovascular Health and Diseases in China 2020 indicated that CAD caused 126.91 deaths per 100,000 inhabitants and 135.88 deaths per 100,000 villagers in China. And the reason for the high mortality rate of CAD was reckoned as the inaccurate and untimely diagnostic strategies (The Writing Committee of the Annual Report on Cardiovascular Health and Diseases in China, 2022). More recently, with the development of gene sequencing, early gene screening and gene-targeted therapy for diseases have become a research hotspot (Depuydt et al., 2020; Tcheandjieu et al., 2022). Therefore, researchers have focused on finding genetic targets for CAD early screening and treatment, which is essential to decrease CAD mortality.
Long noncoding RNAs (lncRNAs), with transcript lengths over 200 nucleotides, play crucial roles in various biological processes, including chromatin modification, transcription regulation, and post-transcription regulation (Bridges et al., 2021; Khan et al., 2023). In recent years, researchers have been committed to finding CAD-associated lncRNAs to reveal the etiological genetics of CAD. For example, lncRNA ANRIL (Fathy et al., 2021), H19 (Hu et al., 2020), MALAT1 (Hu et al., 2019), BANCR (Wang et al., 2019), and lncRNA-MIAT (Ma et al., 2020) were abnormally expressed under disease conditions and closely related to the pathological process of CAD.
Single nucleotide polymorphisms (SNPs) located in lncRNA were critical for exploring the etiological genetics of CAD (Hu et al., 2020; Ma et al., 2020; Manjula et al., 2020). ANRIL rs1004638 AT genotype was linked to higher ANRIL methylation levels and further indicated to be associated with CAD-related clinicopathological parameters (Xu et al., 2021). LncRNA H19 rs4929984 A allele and rs217727 G allele reduced susceptibility to CAD in women (Huang et al., 2021). In short, lncRNA and its genetic variants were associated with CAD risk, revealing the genetic etiology of CAD.
LncRNA RP1-276N6.2 is a newly identified gene in the SLC22A3-LPAL2-LPA gene cluster on chromosome 6q26, whose function is fully unknown. Genome-wide association studies have indicated that this gene cluster was strongly related to lipoprotein a (Lp(a)), a high-risk factor for CAD (Lv et al., 2012; Trégouët et al., 2009). Lp(a), encoded by LPA, is cleaved into fragments by proteases that attach to lesions and promote thrombosis (Reyes-Soffer et al., 2022). Moreover, Lp(a) was linked to pro-inflammatory pathways (Kronenberg et al., 2022). SLC22A3, organic cation transporter protein 3 (OCT3), could affect inflammatory mediator release and low-density lipoprotein cholesterol deposition in vascular endothelial cells by regulating histamine synthesis (Li et al., 2015; Wang et al., 2016).
LncRNA and its risk variants may influence disease risk by regulating the expression of proximate genes in a cis-regulatory manner (Gil and Ulitsky, 2019; Zhao et al., 2015). Winkler et al. (2022) indicated that lincRNA-p21 played a cis-regulatory role in the expression of Cdkn1a/p21. LncRNA CDKN2B-AS1 was linked to atherosclerotic plaque formation by regulating the activity of the neighboring gene CDKN2B promoter (Ou et al., 2020). These investigations revealed that RP1-276N6.2 and its genetic variants may influence the development of CAD by regulating proximate gene expression.
In conclusion, a case–control study was established for investigating the relationship between the RP1-276N6.2 polymorphisms and CAD risk. Cellular experiments explored the function of rs505000 on expression levels of RP1-276N6.2 and SLC22A3 and to further examine the regulatory role of RP1-276N6.2 on SLC22A3. Finally, we proposed that RP1-276N6.2 variants were significantly associated with susceptibility to CAD and that RP1-276N6. 2 might be an early diagnostic and therapeutic target for CAD.
Materials and Methods
Study population
A total of 949 cases and 892 controls matched for age and sex were included between 2012 and 2016 in Chongqing (Chongqing, China). More information on exclusion and inclusion criteria was available elsewhere (Zhou et al., 2008). Questionnaires were completed to collect demographic data. Clinical and biochemical characteristics were obtained from the hospital information collection system. Peripheral venous blood samples were collected from participants and stored at −20℃. Each subject signed the informed consent. All methods were carried out under relevant regulations from the appropriate competent authorities. All experimental protocols were approved by the Ethics Committee at Chongqing Medical University (No. 2021043).
SNP selection and genotyping
Three SNPs (rs611950, rs10499313, and rs505000) in the RP1-276N6.2 gene were selected from the Chinese HapMap project. Linkage disequilibrium analysis was performed by Haploview 4.1 software. We included SNPs with r 2 > 0.80, minor allele frequency >0.1, and the Hardy–Weinberg equilibrium test at p > 0.05. Each subject drew 5 mL of fasting blood with EDTA anticoagulant tubes and centrifuged immediately to obtain plasma. Genomic DNA was separated using the Tiangen DP319-02 Kit (Tiangen Company, Beijing, China). Genotyping was conducted by the TaqMan Genotyping Assay Systems and ABI PRISM 7900HT (Applied Biosystems, Foster City, CA, USA).
Cell culture and transfection
The human umbilical vein endothelial cell (HUVEC) was maintained in DMEM (Gibco, Carlsbad, CA, USA), supplemented with 10% fetal bovine serum and 1% penicillin–streptomycin. Cells were passaged less than seven times and cultured at 37°C in 5% CO2. Specific small interfering RNA for human RP1-276N6.2 (siRNA) and negative control siRNA (siRNA-NC) were purchased from Shanghai Sangon Company (Shanghai, China). Plasmid pcDNA3.1-NC, pcDNA3.1-RP1-276N6.2, pcDNA3.1-rs505000-G, and pcDNA3.1-rs505000-C were synthesized by Wuhan GeneCreate Biological Engineering Company (Wuhan, China). The sequences of siRNA and siRNA-NC are provided in Supplementary Table S1. Transfection was conducted with Lipofectamine 2000 Transfection Reagent (Invitrogen, Carlsbad, CA, USA) according to manufacturer's protocol.
Quantitative real-time polymerase chain reaction
The total RNA was isolated from cells using TRIzol Reagent (TaKaRa, Kyoto, Japan) and converted into cDNA by reverse transcription using 2 × Fast PCR Master Mix (Med Chem Express, Shanghai, China). The real-time polymerase chain reaction (qRT-PCR) was performed with the Bio-Rad® 96 instrument Real-Time PCR System. The sequences of primers are listed in Supplementary Table S1. GAPDH was provided as an internal control for normalization. Data analysis was calculated as the 2−△△CT method.
Enzyme-linked immunosorbent assay
The secreted protein SLC22A3 (OCT3) levels in cell culture supernatants were detected by enzyme-linked immunosorbent assay (Shanghai Yanhui Biotech Company, Shanghai, China). After 48 h of transfection, we harvested the cell culture supernatants. The samples were centrifuged at 3000 r for 15 min, and the corresponding reagents were added according to the instructions. The optical density at 450 nm was measured by a microplate reader. Five independent replicates were conducted.
Statistical analysis
The statistical analysis was performed with SPSS 23.0 (SPSS, Inc., Chicago, IL, USA). Quantitative data were represented as mean ± standard deviation (mean ± SD) and compared using Student's t-test. Count data were presented as percentages and compared using the chi-square test. Univariate and multivariate binary logistic regression were used to demonstrate the association between SNPs and CAD risk. The receiver operating characteristic (ROC) curve analysis and experimental data were analyzed by GraphPad Prism 8.1. All statistical analyses were two sided, with the significance level set at p < 0.05.
Results
Characteristics of the subjects and SNPs
Our case–control study enrolled 949 CAD cases (66.19 ± 10.74 years) and 892 controls (65.62 ± 12.72 years). The basic epidemiological information and clinical characteristics are summarized in Supplementary Table S2. There was no significant difference in sex and age distribution (p > 0.05), indicating that the case and control groups were comparable. Genotype distributions for three SNPs conformed to Hardy–Weinberg equilibrium in the control group (p > 0.05; Supplementary Table S3), indicating that the study subjects were well-representative. All SNPs exhibited a strong LD (r 2 > 0.80; Supplementary Fig. S1). Genotype distribution and allele frequency for the case–control study are displayed in Supplementary Table S4. No significant association was discovered in the case–control groups (p > 0.05).
Association of RP1-276N6.2 polymorphisms with CAD susceptibility stratified by the age of first diagnosis
According to the age of first diagnosis, men ≤55 years and women ≤65 years were defined as the premature CAD group; the others were defined as the delayed CAD group (Wilmot et al., 2015). After adjusting for risk factors, the RP1-276N6.2 rs611950 T allele was significantly linked to an increased risk of premature CAD (odds ratio [OR] = 1.32, 95% confidence interval [CI]: 1.05–1.66, p = 0.02; Table 1). Compared to the wild-type genotype, the RP1-276N6.2 rs10499313 with AG genotype and G allele enhanced premature CAD risk (OR = 2.11, 95% CI: 1.31–3.39, p = 0.002; OR = 1.52, 95% CI: 1.12–2.07, p = 0.01; Table 1). As demonstrated in Table 1, the allele G was significantly linked with a lower risk of developing premature CAD (OR = 0.74, 95% CI: 0.59–0.93, p = 0.01). Interestingly, the rs505000 CG genotype exhibited a higher risk of delayed CAD than that in the rs505000 CC genotype (OR = 1.39, 95% CI: 1.03–1.89, p = 0.03; Table 1), but there were no differences in the frequencies of rs611950 and rs10499313 polymorphism in the delayed CAD subgroup (p > 0.05; Table 1).
Stratification Analysis for Associations Between Three RP1-276N6.2 Single Nucleotide Polymorphisms and the Risk of Premature and Delayed Coronary Artery Disease in the Chinese Han Population
Bold values represent a statistically significant value.
Adjusted for age, gender, smoking, drinking, hypertension, and diabetes mellitus.
CAD, coronary artery disease; CI, confidence interval; OR, odds ratio; SNP, single nucleotide polymorphism.
Further analyses stratified by sex demonstrated that none of the three SNPs had a statistically significant correlation with CAD risk in either the male or female subgroups (p > 0.05; Supplementary Table S5).
Effects of the rs505000 on RP1-276N6.2 and SLC22A3 gene expression levels
Our results found that rs505000 was the only variant associated with premature and delayed CAD. To confirm whether rs505000 regulates the expression levels of RP1-276N6.2 and SLC22A3, the rs505000-C and rs505000-G plasmids were transfected into cells. Our results revealed that RP1-276N6.2 and SLC22A3 expressions in the rs505000-G group were significantly reduced compared with the rs505000-C group (p < 0.05 and p < 0.05, respectively; Fig. 1A, B). We performed a bioinformatic analysis of rs505000 through the GTEx data online website (

The rs505000 affects RP1-276N6.2 and SLC22A3 gene expression levels.
Analysis of the lncRNA RP1-276N6.2 expression levels
In this study, RP1-276N6.2 expression was measured by qRT-PCR. The results revealed that RP1-276N6.2 exhibited lower expression in the CAD case group compared to the control group (p < 0.01; Fig. 2A), indicating that RP1-276N6.2 was closely associated with CAD. According to ROC curve analysis, the diagnostic sensitivity of RP1-276N6.2 was 67.44%, the diagnostic specificity was 77.78%, and the area under the ROC curve was 0.75 (95% CI: 0.65–0.86, p < 0.001; Fig. 2B), which suggested that lncRNA RP1-276N6.2 might serve as a potential diagnostic target for CAD.
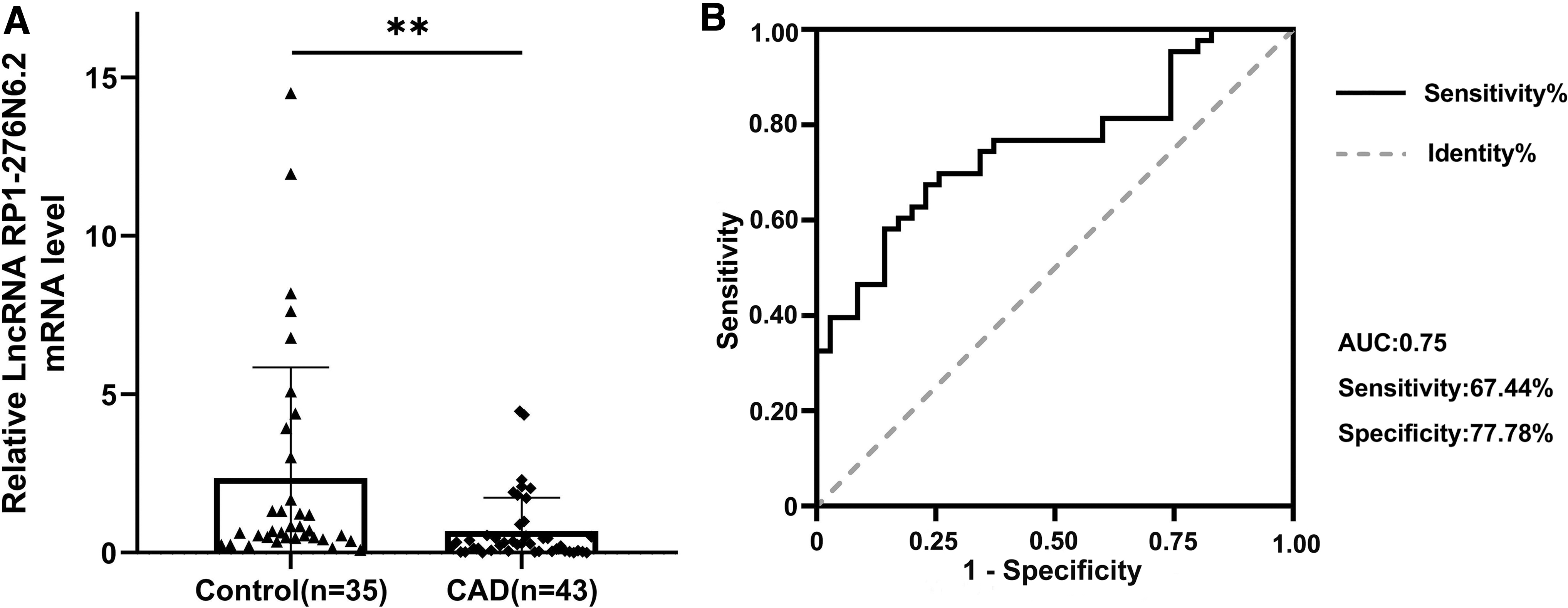
LncRNA RP1-276N6.2 relative expression between CAD cases and controls in the Chinese Han population.
Effects of the lncRNA RP1-276N6.2 on proximate gene regulation
To assess the regulatory role of RP1-276N6.2 on SLC22A3, we transfected siRNA and pcDNA3.1-RP1-276N6.2 into HUVEC. The effciencies of RP1-276N6.2 knockdown and overexpression were verified by qRT-PCR (p < 0.001 and p < 0.001, respectively; Fig. 3A, B). As displayed in Figure 3C, the knockdown of RP1-276N6.2 could significantly downregulate SLC22A3 mRNA expression (p < 0.05; Fig. 3C). After transfection with pcDNA3.1-RP1-276N6.2 plasmid, the gene expression of SLC22A3 was significantly increased (p < 0.01; Fig. 3D). Then, we further explored the effect of RP1-276N6.2 on the protein expression of SLC22A3 (OCT3). As shown in Figure 3, SLC22A3 (OCT3) protein expression dramatically downregulated in the silenced cell model (p < 0.01; Fig. 3E) and significantly increased in the overexpressed cell model (p < 0.05; Fig. 3F). The above data fully demonstrated that RP1-276N6.2 could positively regulate not only the mRNA level but also the protein level of SLC22A3 (OCT3).
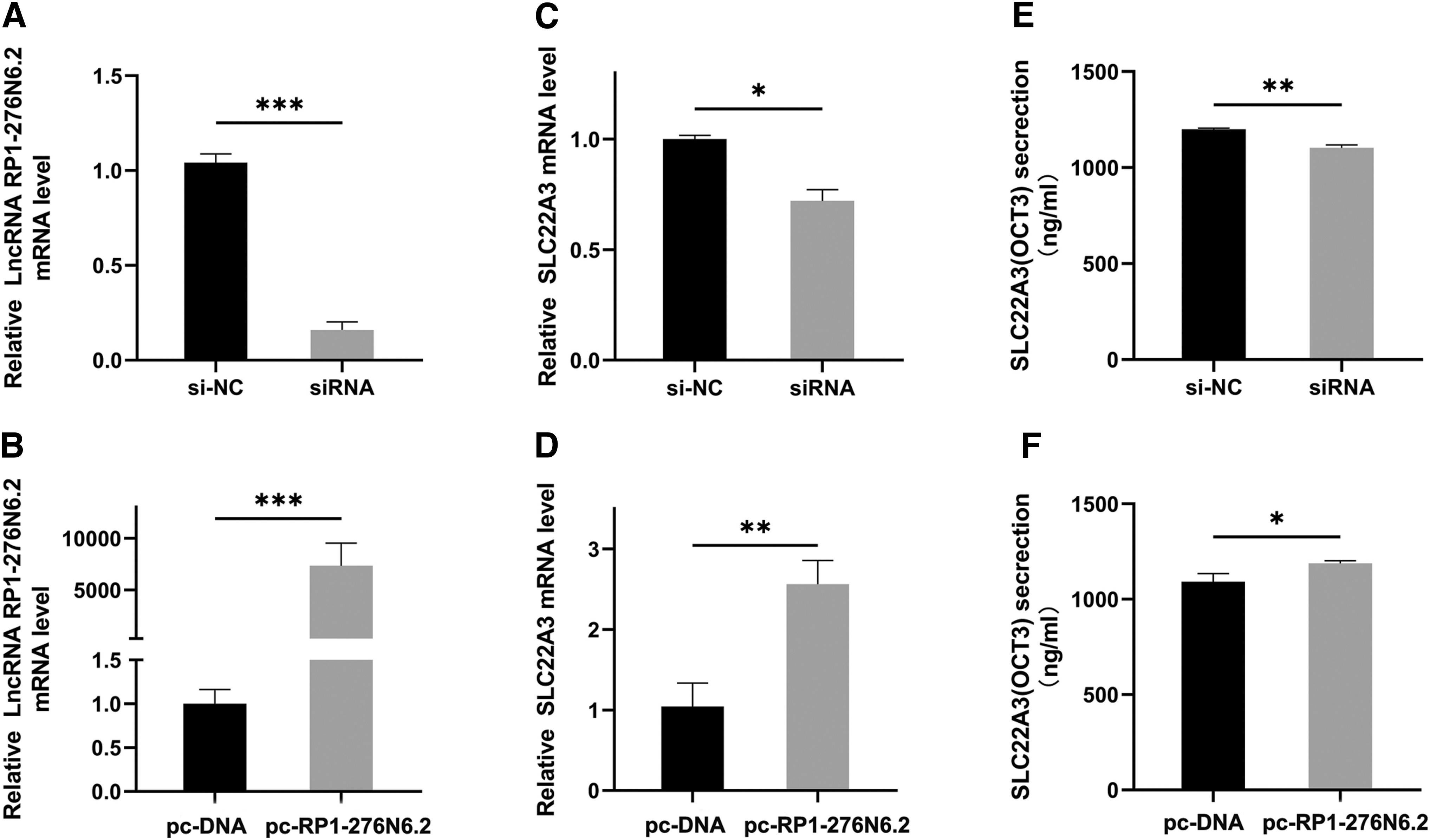
LncRNA RP1-276N6.2 affects SLC22A3 expression.
Discussion
The case–control study investigated the association between RP1-276N6.2 variants and CAD risk in the Chinese Han population. Our findings indicated that the rs611950 C > T and rs10499313 A>G increased premature CAD risk, while the rs505000 C > G decreased this risk. In recent years, the onset age of CAD has tended to be younger (Collet et al., 2019); therefore, SNPs in RP1-276N6.2 were necessary as a sensitive target for premature CAD diagnosis. More interestingly, the rs505000 CG increased delayed CAD risk, which further illustrated the critical role of RP1-276N6.2 variants in CAD risk.
Over the past decades, several studies have documented that functional SNPs affect corresponding gene expression and thus on disease-related biological function (Landeros et al., 2022; Salviano-Silva et al., 2021; Sun et al., 2022; Wang et al., 2020). In our study, the rs611950, rs10499313, and rs505000 are noncoding transcript variants located in RP1-276N6.2 that may regulate biological functions by influencing the expression of lncRNA and the ability to bind to other biomolecules. We concentrated on gene variant rs505000 because only it had a significant association with premature and delayed CAD in our population. The experimental results exhibited that compared with the rs505000-C group, rs505000-G decreased the expression levels of RP1-276N6.2 and SLC22A3, as well as a tendency to regulate LPA expression (Supplementary Figs. S3 and S4).
More interestingly, the GTEx data online website indicated that gene variant rs505000 could affect the relative expression levels of SLC22A3 and SLC22A2 in skeletal muscle, which is consistent with our experimental results. These results imply that rs505000 of RP1-276N6.2 may be a potential functional variation in the etiopathogenesis of CAD.
Our study found that RP1-276N6.2 positively regulated the expressions of SLC22A3 and LPA (Supplementary Fig. S3). LPA and SLC22A3 were well-known in CAD-related lipid metabolism and inflammation (Cheema et al., 2020; Kronenberg et al., 2022; Li et al., 2015; Mo et al., 2018). Thus, RP1-276N6.2 may be involved in inflammation and lipids, which was critical for the pathogenesis of CAD. Similarly, in our findings, rs505000 influenced the molecular expression levels of RP1-276N6.2 and SLC22A3, suggesting that rs505000 may have a potential regulator role in CAD-related inflammation and lipid metabolism. However, its role and mechanisms in lipid metabolism and inflammation may require further experimental confirmation.
LncRNA RP1-276N6.2 is a novel molecule that has not been proven to be associated with any disease. We first identified the connection between the RP1-276N6.2 variants and susceptibility to CAD in the Chinese Han population. However, several limitations remain to be solved. First, the case–control sample size was relatively small, which may limit the representativeness of the general population. Second, due to the Corona Virus Disease 2019 outbreak, we were unable to collect new and eligible case–control samples for this study. And due to the longevity of the previous sample, we were unable to replicate the study in independent sample sets. Third, because of time and resource restrictions, functional experiments on the regulation of CAD-related inflammation and lipids by the RP1-276N6.2 variants were unable to be elucidated.
In summary, our evidence indicated that RP1-276N6.2 rs611950 and rs10499313 were potentially risk variants for premature CAD. RP1-276N6.2 rs505000 may be a potential functional variation, most probably by regulating RP1-276N6.2 and SLC22A3 expression levels. Thus, we proposed that RP1-276N6.2 variants were closely related to the risk of CAD, and RP1-276N6.2 may be a promising early diagnostic and therapeutic target. In the future, larger sample sizes and non-Asian populations will validate the relationship between RP1-276N6.2 variants and CAD risk, and the underlying mechanism of RP1-276N6.2 in the progression of CAD requires further investigation.
Conclusions
Our study found that the RP1-276N6.2 rs611950 C > T and rs10499313 A>G closely increased the premature CAD risk, and rs505000 C > G regulated the expression levels of RP1-276N6.2 and SLC22A3, which was associated with premature and delayed CAD. LncRNA RP1-276N6.2 may be a potential genetic target for CAD early diagnosis and treatment.
Footnotes
Acknowledgments
The authors are grateful to all the study participants. The authors also thank the editors for editing this article.
Disclosure Statement
No competing financial interests exist.
Funding Information
This work was supported by the National Natural Science Foundation of China (Grant No. 81773519 to L.Z.).
Supplementary Material
Supplementary Figure S1
Supplementary Figure S2
Supplementary Figure S3
Supplementary Figure S4
Supplementary Table S1
Supplementary Table S2
Supplementary Table S3
Supplementary Table S4
Supplementary Table S5
References
Supplementary Material
Please find the following supplemental material available below.
For Open Access articles published under a Creative Commons License, all supplemental material carries the same license as the article it is associated with.
For non-Open Access articles published, all supplemental material carries a non-exclusive license, and permission requests for re-use of supplemental material or any part of supplemental material shall be sent directly to the copyright owner as specified in the copyright notice associated with the article.