
Abstract
We have shown in the past decade, for the first time in a bivalve mollusc, detection, isolation, and purification of β-1,3 glucan binding protein (β-GBP) in the plasma of the marine mussel Perna viridis and demonstrated its role in a nonself-induced activation of plasma prophenoloxidase system. In this study, we present evidence for its ability to function as an opsonin during phagocytosis of trypsinized yeast cells by the hemocytes of P. viridis. The in vitro pretreatment of target cells (trypsinized yeast cells) with β-GBP enhanced the phagocytic response of hemocytes. Such β-GBP-mediated enhanced phagocytic response appeared to be dose dependent. This opsono-phagocytic response could be inhibited by the presence of laminarin (a polymer of β-1,3 glucans), glucose, as well as polyclonal antibodies raised against β-GBP. These observations clearly indicate that the plasma β-GBP can possibly recognize and bind to β-1,3 glucans on the surface of targets and facilitate hemocyte recognition processes possibly by forming a bridge between the hemocytes and the target, consequently leading to opsono-phagocytosis. These observations together with our earlier annotations indicate the multifunctional potential of plasma β-GBP in the marine mussel P. viridis.
Introduction
The innate immune system in invertebrates plays a vital role in defense against different pathogens due to the lack of adaptive immune response characteristics like that found in vertebrates. In innate immunity, phagocytosis appeared to be the major effector mechanism by which invertebrates defend against the invading micro-organisms, and this phagocytosis involves recognition, binding, and internalization of pathogens. Such a phagocytic uptake of pathogens by hemocytes, together with humoral components, constitutes the first line of defense (Ratcliffe et al., 1985; Söderhäll and Cerenius, 1992). The uptake of particles by these cells has been shown to be significantly promoted by factors called as opsonins (Richards and Renwrantz, 1991). Although, the process of phagocytosis has been extensively characterized in a few selected invertebrates, especially in the case of crustaceans and insects, such information in molluscs are limited (Mullainadhan and Renwrantz, 1986).
Recognition and phagocytosis of foreign particles in molluscs have been shown to involve lectins with specific carbohydrate binding properties and this comprises both cell membrane-bound and humoral lectins (Sminia et al., 1979). This has been demonstrated in the clearance of foreign particles from Helix pomatia (Renwrantz et al., 1981) and phagocytosis of yeast by hemocytes in vitro of Mytilus edulis and Mytilus galloprovincialis, thereby demonstrating the opsonic role for hemolymph lectins in the phagocytic uptake of target cells (Renwrantz and Stahmer, 1983; Santarem et al., 1994).
A number of humoral opsonizing proteins, including lectins, hemolin, lipopolysaccharides (LPS), β-1,3 glucan binding protein (β-GBP), and peptidoglycan recognition protein, have been described as candidate pattern recognition proteins (PRPs) for a variety of bacterial and fungal cell wall components in crustaceans and insects, which are also referred to as pathogen-associated molecular patterns (PAMPs) (Du et al., 2007; Dunphy et al., 1998; Lee et al., 2000). PAMPs are biologically important as they trigger a variety of innate immune responses, as has been shown in insects and mammals. The process of phagocytosis was triggered by the binding of PAMPs with their cell surface receptors on the phagocytes. These receptors not only perform phagocytic functions but also bridge the gap between extracellular and intracellular communication, leading to signal transduction and the production of inflammatory mediators, which are crucial for clearing the invading pathogens and maintaining cell homeostasis (Zhang et al., 2022).
In invertebrates, proteins with affinity for β-1,3 glucan have been demonstrated in silkworm (Ochiai and Ashida, 1988), crayfish (Duvic and Söderhäll, 1990), earthworm (Beschin et al., 1998), and horseshoe crab (Muta et al., 1995). These β-GBPs have been purified (Duvic and Söderhäll, 1990; Seki et al., 1994; Söderhäll et al., 1988) and shown to enhance the activity of both the prophenoloxidase (proPO)-activating enzyme (Wang and Jiang, 2006) and phenoloxidase (PO) (Ma and Kanost, 2000). In addition, crayfish β-GBP also induced spreading and partial degranulation of isolated granular hemocytes (Barracco et al., 1991) and the presence of binding site of the β-GBP on the hemocyte surface was demonstrated with immunofluorescence using anti β-GBP, and this β-GBP membrane receptor was purified from crayfish hemocytes (Duvic and Soderhall, 1992).
The phagocytic and encapsulation activity of purified β-GBP from Portunus pelagicus using yeast cells of Saccharomyces cerevisiae alongside sepharose bead suspension revealed that, β-GBP has the ability to detect the PAMPs found on the surface of fungi and bacteria. The recognition of invading foreign substances and the involvement of multifunctional activities induced the activation of proPO, thereby leading to a conclusion that β-GBP plays a major role in the innate immune system of crustaceans by stimulating the proPO system and its cascade mechanisms (Anjugam et al., 2016).
Nevertheless, most studies on β-GBP in invertebrates have indicated a major role of this protein in proPO system activation and only three studies, including two crustaceans (Cerenius et al., 1994; Thörnqvist et al., 1994) and a mosquito (Wang et al., 2006), have suggested a role of isolated β-GBP in phagocytosis and melanotic encapsulation. However, earlier studies on the presence of β-GBP in the circulating fluid ( = plasma) or hemolymph in molluscs are limited (Söderhäll et al., 1988).
We have recently shown the presence and purification of β-GBP from plasma of Perna viridis and this β-GBP had a native molecular weight (MW) of 510 kDa and involved in activation of plasma proPO (Jayaraj et al., 2008). Furthermore, the purified β-GBP had inherent serine protease activity, but not glucanase, and agglutinated a variety of mammalian red blood corpuscles, bacteria and trypsinized yeast cells (Jayaraj et al., 2008). This suggested that the purified β-GBP is a multipotential protein and also enabled us to speculate whether it has any role in cellular immune response, particularly phagocytosis.
In lieu of the lack of literature on other possible functions of β-GBP with invertebrates in general and molluscs in particular, we undertook this study primarily to identify the opsonic role, if any, of P. viridis β-GBP. We report in this study the “lectin-bridging hypothesis” for P. viridis β-GBP in enhancing hemocyte-mediated phagocytosis of yeast cells, in vitro.
Materials and Methods
Experimental animals and plasma collection
Specimens of P. viridis (shell length 8–12 cm) collected from the rocky shores of Royapuram, Chennai, India, were maintained in laboratory in seawater (30%; 26°C ± 2°C) with continuous aeration until use. The experimental animals were collected from beneath the rocks or any substratum and were acclimatized in the laboratory conditions for 48 h before use. The experimental animals were not fed with anything directly as they are filter feeders; they feed on the planktons available in the seawater [360/01/a/CPCSEA is the ethical clearance number from the institutional review board]. The hemolymph of P. viridis was collected following Asokan et al. (1997) and immediately centrifuged (400 g, 5 min, 4°C) and the resulting supernatant was used as plasma.
Preparation of yeast cell suspension and trypsinization
The yeast cell targets for phagocytosis by hemocytes were prepared following Arumugam et al. (2000). Yeast on its own nature has β-glucans on their surface, which possess binding sites for β-GBP and that makes the phagocytosis study more relevant. Essentially, 500 mg commercial grade baker's yeast (S. cerevisiae) suspended in 10 mL saline was heat inactivated, cooled to room temperature (RT) and washed thrice with saline and once with tris buffer saline (TBS)-I (50 mM tris and 403 mM NaCl, pH 8.4, 840 mOsm) by centrifugation (400 g, 5 min, RT), and finally suspended in the same buffer.
Trypsinization of yeast cells was carried out by suspending 1 mL of yeast cell suspension prepared as above, with 20 volumes of TBS-II (10 mM tris-HCl and 145 mM NaCl, pH 7.5, 300 mOsm) containing trypsin (0.5%) and incubated (1 h at 37°C) with occasional and gentle shaking. At the end of incubation, the trypsinized yeast cells were washed once with TBS-II and finally re-suspended in TBS-III (50 mM tris-HCl, 100 mM NaCl, and 10 mM CaCl2, pH 7.5, 300 mOsm) as 0.5% (v/v) suspension.
Purification of β-GBP
β-GBP was purified from plasma as previously described by Jayaraj et al. (2008). Briefly, the plasma diluted with TBS-III (50 mM tris-HCl, 100 mM NaCl, and 10 mM CaCl2, pH-7.5, 300 mOsm) exhibited an yeast agglutination titer value of 16 against trypsinized yeast cells, was passed through laminarin-Sepharose CL 6B column at a flow rate of 3 mL·h−1. After removing unbound components and reequilibration, bound β-GBP was eluted (4 mL·h−1) with 10 mL of 0.1 M acetate buffer (pH 4.5) containing 2.5 M MgCl2 and 1 M D-glucose, and the fractions that showed absorbance at 280 nm and agglutinated trypsinized yeast cells (yeast agglutination titer: 256) were collected, pooled, and dialyzed extensively (MW exclusion limit <14,000 Da) against TBS-II at 10°C, and protein content measured using bovine serum albumin as standard (Lowry et al., 1951).
Analysis of purified β-GBP in polyacrylamide gel electrophoresis
The protein profile of purified β-GBP (25 μg) was analyzed in discontinuous polyacrylamide gel electrophoresis (PAGE) under nondenaturing conditions following Maure (Ansorge, 1985) r's method, using a 3% stacking gel (pH 6.7), using a 4.5% separating gel (pH 8.9) in tris-glycine buffer (pH 8.3), and at a constant current of 4 mA per sample and at 10°C in a slab gel measuring 170 × 150 × 1.5 mm and stained with silver nitrate following the procedure of Ansorge (1985). The MW of the purified molecule was calculated to be 510 kDa (Fig. 1A).

Native PAGE of β-GBP isolated from the plasma of Perna viridis [Lane I: Protein profile of β-GBP (25 μg) isolated from the plasma of P. viridis stained with silver nitrate; Lane II: Position of MW markers]
Preparation of polyclonal antibody against purified β-GBP and immunoblotting
Polyclonal antibody against purified β-GBP was raised in Swiss albino mice by five injections of 100 μg purified β-GBP each time with Freund's complete adjuvant for the first injection followed by Freund's incomplete adjuvant for the other four injections. The antisera collected (1 mL) at the end of the immunization schedule (5 weeks) was passed through a column of Protein A-CL Agarose matrix (8 mm diameter × 45 mm height) to purify the polyclonal antibody raised against β-GBP.
The column was washed and equilibrated with 0.1 M sodium phosphate buffer (pH 8.4) and the bound antibody was eluted using 10 mL of 0.1 M glycine—HCl (pH 2.5). The eluants (10 mL) collected were estimated for protein concentration following Bradford method (Bradford, 1976) and dialyzed extensively against 1 × phosphate buffer saline (PBS) (150 mM NaCl, 50 mM Na2HPO4, and 50 mM NaH2PO4, pH 7.0) and stored in vials at −20°C for further use. Analysis of antiserum by double immunodiffusion (Fig. 1B) tests, immunoelectrophoresis (immunoelectrophoretic, Fig. 1C), and enzyme-linked immunosorbent assay (data not shown) was carried out to confirm the presence of polyclonal antibody against purified β-GBP following Phillips, 1978.
For immunoblotting, the proteins were subjected to 4.5% native-PAGE under denaturing conditions and electrotransferred to a nitrocellulose membrane (0.45 μm) in transfer buffer (25 mM tris-HCl, 192 mM glycine, and 10% methanol) for 2 h at 280 mA on ice. The membrane was subsequently blocked using a blocking buffer (1 × PBS, 0.05% Tween 20, and 5% non fat dry milk) for 1 h. The membrane was then incubated with a primary antibody, that is, anti β-GBP polyclonal mice antibody at a dilution of 1: 500 (primary antibody), overnight at 4°C, and with a secondary antibody, goat anti-mouse IgG-HRP conjugate at a dilution of 1:3000 (secondary antibodies). The dilutions of 500 × and 3000 × are the standard protocol for both primary as well as secondary antibodies, respectively, for 2 h at 4°C. For detection, 0.01%, 3′-diaminobenzidine tetrahydrochloride, and hydrogen peroxidase (H2O2) were used.
Phagocytosis of trypsinized yeast cells by hemocytes
Phagocytosis assay was carried out using trypsinized yeast cells as targets following Arumugam et al. (2000).
Preparation of hemocyte monolayer
For the preparation of hemocyte monolayers for phagocytosis assays, the hemolymph (200 μL) from each mussel was withdrawn into a 2 mL sterile plastic syringe containing 800 μL ice-cold iso-osmotic TBS (50 mM tris, 403 mM NaCl, and 0.1 mM CaCl2, pH 8.4, 850 mOsm), centrifuged (400 g, 5 min, 4°C), and hemocyte pellet was resuspended in iso-osmotic TBS. Hemocyte monolayers were prepared by placing 200 μL of diluted hemocyte suspension (3.29 ± 0.14 × 106 cells/mL), spread on an alcohol washed, dried, and wiped glass slide, and kept in a moist chamber for 30 min at 23°C to allow the hemocytes to settle and attach to the surface of the glass slide (Asokan et al., 1998).
Hemocyte viability
The viability of hemocytes in monolayers was determined using the trypan blue dye exclusion test following Garvey et al. (Phillips, 1978). Trypan blue bye exclusion is based on the principle that live cells possess intact cell membranes that exclude certain dyes, such as trypan blue, eosin, or propidium, whereas dead cells do not. In this test, a cell suspension is simply mixed with trypan blue dye and then visually examined to determine whether the cells take up or exclude dye.
In vitro phagocytosis assay
A known volume of trypsinized yeast cells (10 μL) were overlaid and incubated with hemocyte monolayer for 10 min. The monolayers were then gently washed with iso-osomotic TBS and fixed with Tris-buffered glutaraldehyde (1.5%). The hemocytes were then examined under Carl Zeiss phase contrast or Axioplan ACE microscope system. The phagocytic rate of hemocytes containing one or more yeast cells was counted after evaluating 200–300 hemocytes randomly, and the percentage phagocytosis was calculated.
Effect of β-GBP on phagocytosis of yeast cells by hemocytes
There were totally seven sets of experiments performed in vitro to study the influence of purified β-GBP on yeast cell phagocytosis by P. viridis hemocytes, which include trypsinized yeast cells suspended in buffer alone (control), trypsinized yeast cells incubated with plasma, laminarin alone, β-GBP alone, β-GBP preincubated with laminarin, glucose, or polyclonal antibody raised against purified β-GBP. All these pretreated trypsinized yeast cells were presented to individual hemocyte monolayers and assessed for phagocytic uptake as described elsewhere.
Laminarin is the polymer of beta glucans and therefore has binding sites for β-GBP in a natural way, and the experiment was conducted for laminarin alone to substantiate the above said statement. The following are the seven sets of experiments performed in vitro to study the influence of purified β-GBP on yeast cell phagocytosis by P. viridis hemocytes. (1) Trypsinized yeast cells suspended in buffer alone (control), (2) trypsinized yeast cells incubated with plasma, (3) laminarin alone, (4) β-GBP alone, (5) β-GBP preincubated with laminarin, (6) glucose, or (7) polyclonal antibody raised against purified β-GBP. The results are discussed in 3.5 in detail along with Figure 4.
Dose dependency
To study the β-GBP-induced dos- dependent phagocytic uptake of trypsinized yeast cells by hemocytes, six tubes containing 10 μL trypsinized yeast cell suspension each were pretreated with different concentrations of purified β-GBP (0, 10, 20, 30, 40, 50, 60, and 75 μg protein·mL−1) and incubated with occasional shaking at 23°C. After 30 min, all the pretreated trypsinized yeast cells were washed with iso-osmotic TBS and resuspended in the same buffer as 1% (v/v) suspension and were used for phagocytic assay, as described elsewhere.
Statistical analyses
Each test series in an experimental condition was performed using hemocytes from five mussels. The difference between control and experimental values or between two experimental values was tested for statistical significance using paired sample Student's t-test and correlation co-efficient (r value) (Bailey, 1995).
Results
Purification of plasma β-GBP
The plasma β-GBP was purified using Laminarin-Sepharose CL6B affinity column chromatography as 1:1 ratio (Plasma: Tris Buffer Saline-I) with a total protein value of 32.45 mg which was having a agglutination titer value of 16 against trypsinized yeast cells was passed through the column for purification purpose. The effluent collected during subsequent washing of this matrix with TBS-I did not contain detectable level of agglutinating activity against trypsinized yeast cells. This indicates that all the agglutinating activity in the plasma was adsorbed to the affinity matrix. When 0.1 M acetate buffer (pH 4.5) containing 2.5 M MgCl2 and 1 M D-glucose was passed through the column, a sharp symmetrical peak with absorbance at 280 nm, as well as a coincident peak of agglutinating activity (titer = 256) against trypsinized yeast emerged from the affinity matrix.
The chromatographic separation of β-GBP using laminarin-sepharose 6B yielded a 90-fold increase in specific yeast agglutinating activity compared to the initial sample. In PAGE analysis, the MW of the isolated protein was calculated to be 510 kDa, as represented in Figure 1A. (Jayaraj et al., 2008). The actual MW of the purified β-GBP in native PAGE is 510 kDa and Lane II of Figure 1A represents the SDS PAGE of the β-GBP, which is split up from one single purified β-GBP molecule from the plasma.
Production of polyclonal antibody against purified β-GBP and immunoblotting
The purified β-GBP was subjected to native PAGE (4.5%) under nonreduced condition and processed for Western blot analysis, using anti β-GBP polyclonal antibody to identify and confirm the presence of anti β-GBP IgG molecules against isolated β-GBP from the plasma of P. viridis (Fig. 1D).
Hemocyte viability and morphotypes
The hemocyte monolayers prepared on a glass slide showed good attachment and spreading of hemocytes. Over 90% of hemocytes were viable up to 3 h, as revealed by trypan blue dye exclusion test (data not shown). During spreading, both nuclei and cytoplasmic contents of hemocytes became more clearly visible, which enabled easy identification of three distinct hemocyte morphotypes based on the presence and abundance of cytoplasmic granules (Fig. 2).
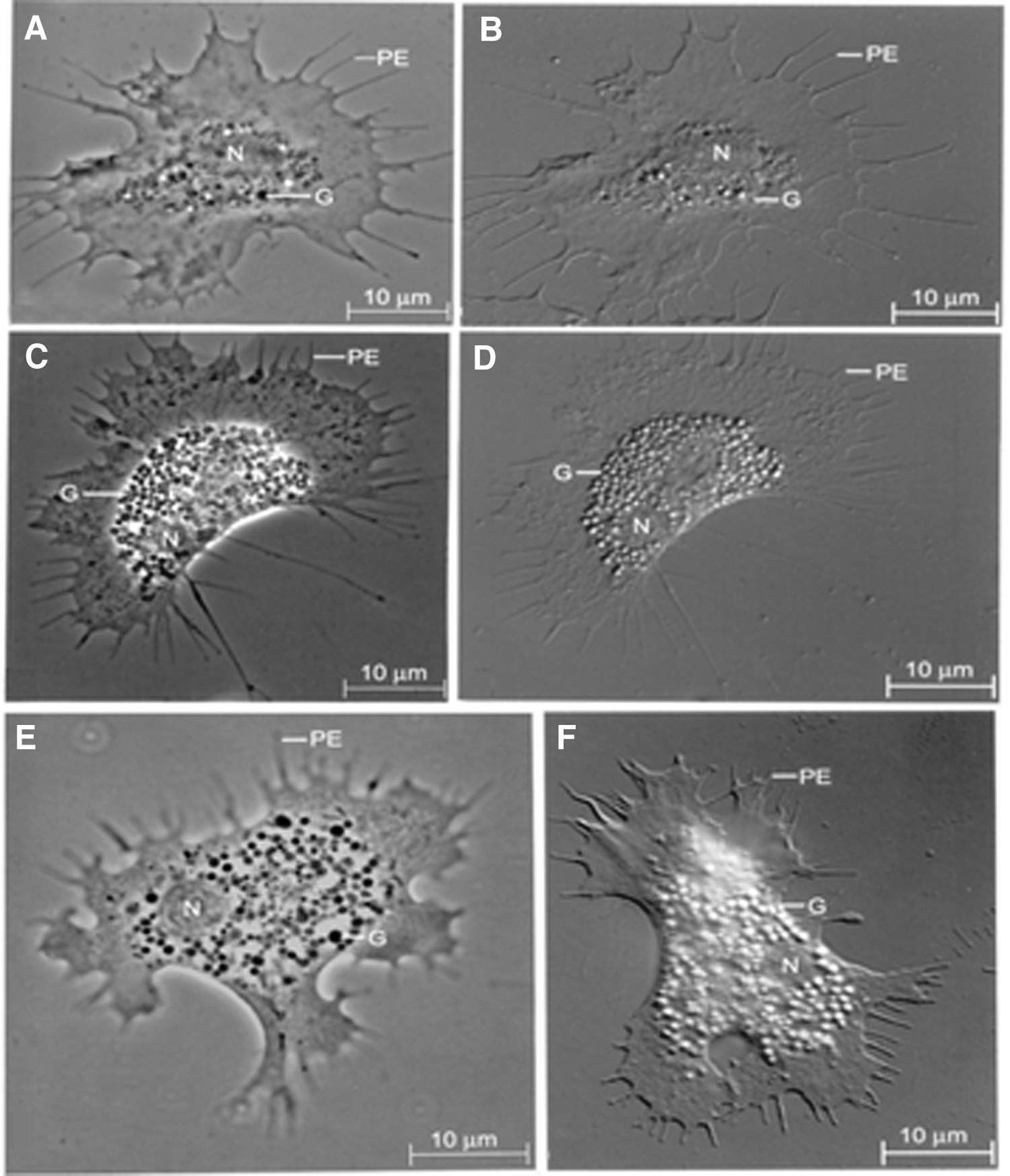
Phase contrast
These morphotypes were hyaline or agranular cells with relatively few or no granules, semigranular cells with moderate number of granules, and granular cells with abundant granules of varying size in the cytoplasm, constituting 14.81% ± 1.4%, 24.60% ± 2.1%, and 60.67% ± 3.03% of the total hemocytes, respectively. The three different hemocyte morphotypes were identified based on the cytoplasmic granules present in each of them. As shown in Figure 2, when the presence of granules is none or few, then they are considered agranular or hyaline cells, when moderately present, they are considered semigranular, and when they are present abundantly, they are considered granular cells.
To calculate the proportion of different hemocyte morphotypes, differential hemocyte counting is done either manually using phase contrast microscope or cell sorter.
In vitro phagocytosis assay
The hemocyte monolayers overlaid and incubated for 10 min with yeast cells suspended in iso-osmotic buffer exhibited attachment of a few yeast cells to hemocyte surface, while many free yeast cells were found in monolayer (Fig. 3A). Under these conditions, free as well as hemocyte-attached yeast cells were refractile, surrounded by bright rings, and clearly visible under phase optics. Upon extension of the incubation time, several hemocytes contained up to five yeast cells intracellularly, thereby demonstrating phagocytic ability of P. viridis hemocytes in vitro. In this test system, the intracellular (ingested) yeast cells appeared darker and markedly lost their surrounding bright rings compared with free or extracellularly attached yeast cells (Fig. 3B).
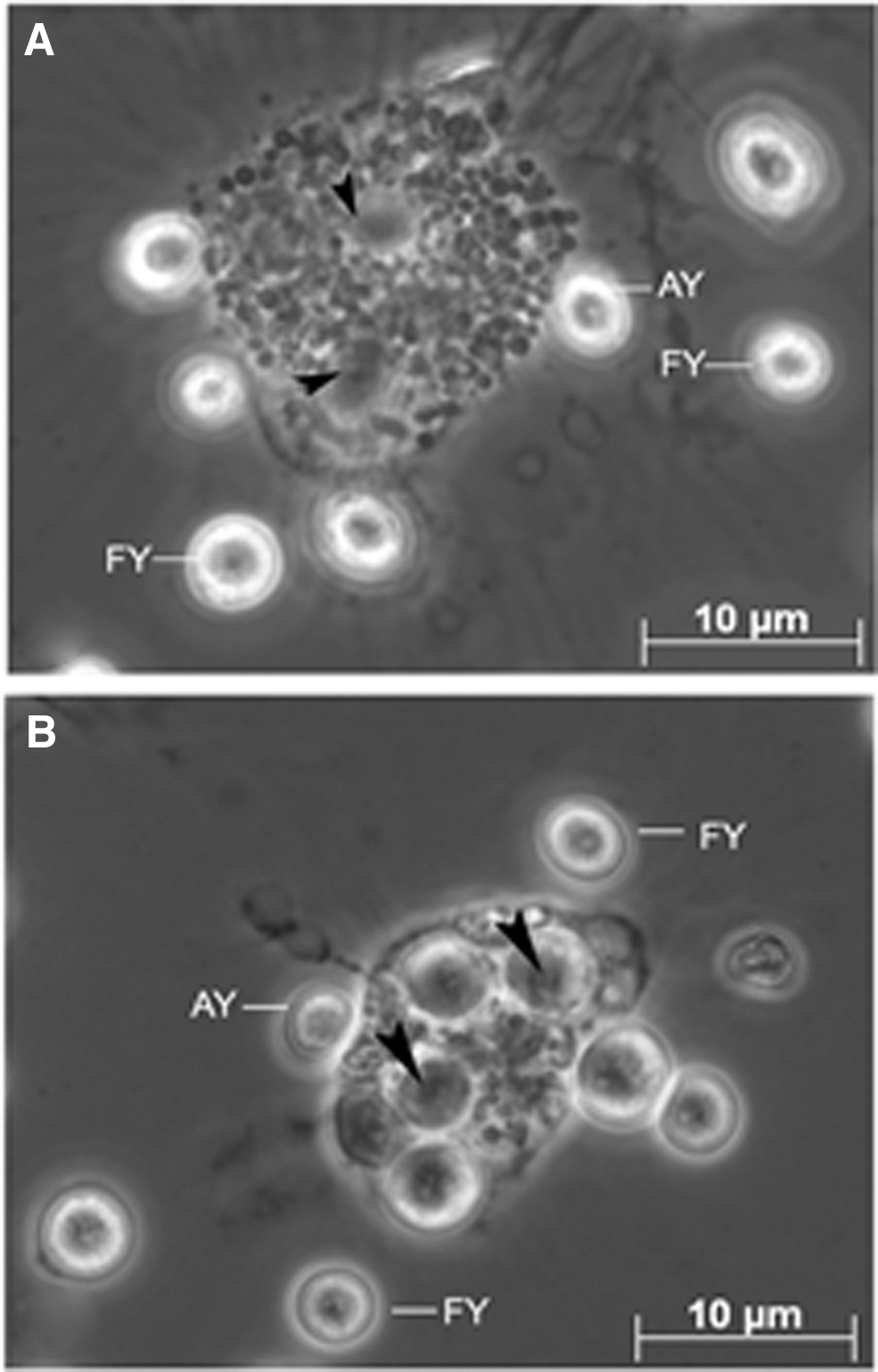
Phase contrast photomicrographs of the phagocytic uptake of yeast cells by live hemocytes of P. viridis.
Effect of β-GBP on phagocytosis of yeast cells by hemocytes
The hemocyte monolayers overlaid with trypsinized yeast cells, and incubated for 10 min at 23°C ± 2°C, showed that 20% of hemocytes appeared with ingested trypsinized yeast cells, (control, Fig. 4), whereas hemocyte monolayers presented with trypsinized yeast cells pretreated with laminarin (1 mg) showed a hemocyte phagocytic rate up to 26%. On the other hand, the trypsinized yeast cells pretreated with purified β-GBP (10 μg) significantly increased the number of phagocytic hemocytes to 66% (p < 0.003; Fig. 4). Interestingly, incubation of hemocytes with trypsinized yeast cells pretreated with (preincubated) laminarin and β-GBP mixture showed a significant decrease (35%) in the number of phagocytic hemocytes (p < 0.05; Fig. 4), compared to pretreatment of yeast cells with β-GBP alone (66%). Similarly, the hemocyte monolayers presented with yeast cells pretreated with β-GBP+glucose or polyclonal antibody raised against purified β-GBP showed a phagocytic rate of 28%, and 27%, respectively.
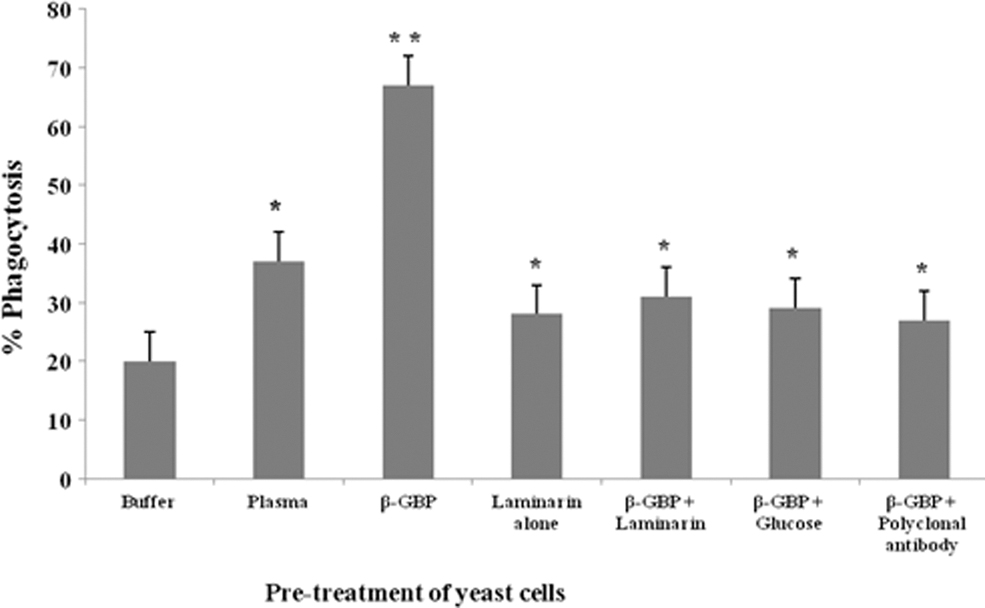
In vitro phagocytosis of trypsinized yeast cells by the hemocytes of P. viridis. Each vertical bar represents mean ± SD using samples from three different preparations. (1) The difference in phagocytic response between hemocytes provided with buffer-treated yeast cells and plasma (1 mg protein) or purified β-GBP (10 μg protein) is statistically significant at *p < 0.05 and **p < 0.003 levels, respectively. (1). The decrease in phagocytic response between hemocytes provided with β-GBP-treated yeast cells and laminarin (1 mg/mL), β-GBP preincubated with laminarin, glucose (1 mg/mL), or polyclonal antibodies raised against purified β-GBP (1 mg protein) are statistically significant at *p < 0.05 level. SD, standard deviation.
Dose-dependent changes of β-GBP on phagocytosis
When hemocyte morphotypes were incubated with trypsinized yeast cells pretreated with increasing concentrations of purified β-GBP (10 to 75 μg), there was a dose-dependent increase in the number of phagocytic hemocytes from 28% to 48% for trypsinized yeast cells (r = 0.995; Fig. 5) compared to control.
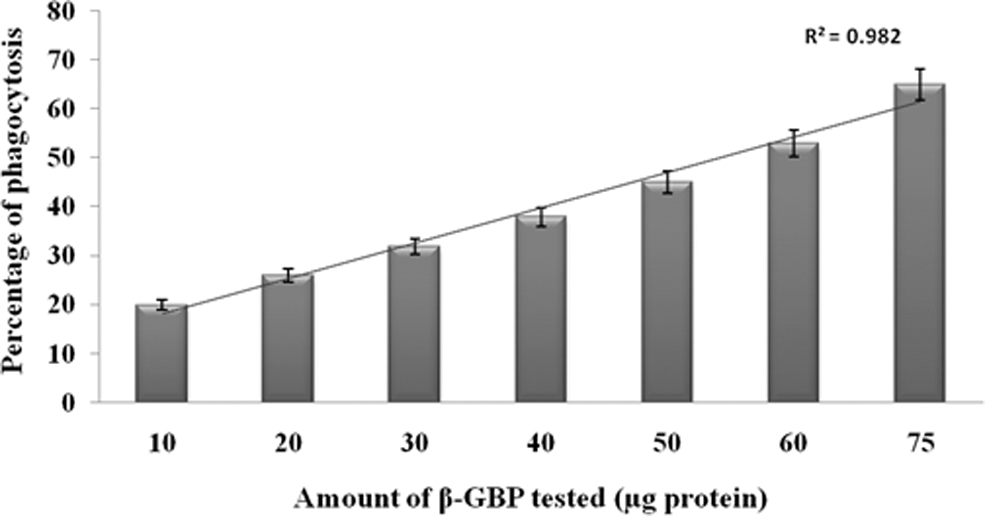
Dose-dependent enhancement of phagocytic response of hemocytes of P. viridis against trypsinized yeast cells pretreated with different concentrations of purified β-GBP. Each vertical bar represents mean ± SD using samples from six different preparations.
Discussion and Conclusion
In arthropods and molluscs, a variety of nonself materials such as bacteria, erythrocytes, and fungal spores are recognized and ingested by hemocytes (Ratcliffe et al., 1985). The uptake of these particles by the molluscan hemocytes has been significantly promoted by humoral components either present in the circulating fluid or released by the granular cells (Mullainadhan and Renwrantz, 1986). In addition, there are also evidences pointing to the factor(s) associated with the activation of humoral or cellular proPO by specific components of bacteria or fungal cell wall substances. Supportingly, Foukas et al. (1998) have clearly shown that both the attachment as well as the internalization of radiolabeled LPS-catechol adduct (LPS-Y) and Escherichia coli by the hemocytes of an insect Ceratitis capitata were inhibited by anti-PO antibodies and phenylthiourea, a known PO inhibitor.
Interestingly, phagocytosis does not appear to be solely dependent on these factors, since a few studies with crustacean hemocyte morphotypes such as agranular or hyaline cells have shown that they are capable of responding to bacteria or fungal spores in the absence of detectable levels of PO or its precursors (Cerenius et al., 1994; Thörnqvist et al., 1994). Our earlier study has indicated that the plasma of the bivalve mollusc P. viridis possesses bacterial or yeast agglutinating property and a specific laminarin binding protein (β-GBP) with a native molecular mass of 510 kDa (Jayaraj et al., 2008). Hence, in this study, we attempted to study the opsonic role of this purified β-GBP in phagocytosis of yeast cells.
In this study, we report the role of β-GBP isolated from P. viridis plasma in mediating opsono-phagocytosis of yeast cells by hemocytes (Fig. 4); in this connection, it may be noted that β-GBP isolated from the plasma of crustaceans (Carcinus maenas and Pacifastacus leniusculus) has been shown to enhance the phagocytic activity of hyaline cells when presented with fluorescein isothiocyanate (FITC)-labeled yeast cells (Cerenius et al., 1994; Thörnqvist et al., 1994). In addition, β-GBP isolated from crustaceans appears to be a multifunctional protein involved in the activation of proPO system (Duvic and Söderhäll, 1990) and triggering of partial degranulation and spreading of granular cells (Barracco et al., 1991).
The hemocytes of P. viridis are capable of phagocytic uptake of yeast cells, as observed in control experiments. A twofold to threefold increase in the phagocytic uptake of β-GBP-treated yeast cells by hemocytes clearly indicated that β-GBP may serve as an opsonin by enhancing the phagocytic activity of hemocytes and this effect was dose dependent. An abrogation of phagocytic uptake of β-GBP-treated yeast by hemocytes in the presence of laminarin (a polymer of β-1,3 glucans) lends additional support to our contention that β-GBP indeed possibly serves as bridging molecule between trypsinized yeast cells and hemocytes, thus indicating that β-GBP also functions as a recognition molecule of foreign materials with β-1,3 glucan-rich surface such as yeast cells.
In a previous study, Ratcliffe et al. (1984) have suggested that laminarin binding to the surface receptor/binding protein on the hemocytes of the insect Galleria melonella resulted in degranulation and release of activated proPO from hemocytes, as well as generation of intermediary opsonic molecules, which could be responsible for opsono-phagocytosis of yeast cells. The abrogation of such opsono-phagocytosis by the presence of p-nitrophenyl p-guanidinobenzoate, an inhibitor of proPO activation system, further supported these suggestions. β-GBP, a PRP, plays a critical role in triggering the innate immune response by detecting β-glucan found on the surface of microbes and the purified Ph-β-GBP exhibited increased agglutination, phagocytic activity, and encapsulation in a dose-dependent manner, indicating the involvement of Ph-β-GBP in cellular immune response against pathogens in crustaceans.
Moreover, the addition of Ph-β-GBP increased the proPO and serine protease activity, possibly contributing to the clearance of pathogens leading to a conclusion that β-GBP triggers proPO activating system in rice field crab Paratelphusa hydrodromus and plays a vital role in innate defense mechanism against the invading pathogens (Iswarya et al., 2017). The purified serine protease beta-glucan binding protein (Ss-β-GBP) was functionally analyzed by yeast agglutination and phagocytic reaction assays along with PO enhancing ability of Ss-β-GBP. The antibacterial activity of the Ss-β-GBP against Gram-positive (Enterococcus faecalis and Staphylococcus aureus) and Gram-negative (E. coli and Pseudomonas aeruginosa) bacteria was also evaluated by determining its minimum inhibitory concentration for all tested species leading to a fact that β-GBP is a multifactorial or multifunctional protein, which has the ability to enhance most immunological activities in invertebrate animals (Divya et al., 2018).
Lipopolysaccharide and β-1,3 glucan-binding protein (LGBP), which existed in the diversity of invertebrates, including shrimp, plays a crucial role in an innate immunity by mediating the recognition of invading pathogens. Litopenaeus vannamei LGBP (LvLGBP) might act as a pathogenic recognition protein to activate shrimp immune defense against invading pathogens through the agglutination, binding, and enhancing encapsulation and PO activity of hemocytes.
Although, to date, this protein lost its activity during the course of evolution, its glucan binding activity has been maintained as a PRP, thereby leading to the confirmation again by the multifunctional aspects of the purified native LvLGBP, including bacterial agglutination, direct binding activity, proPO activation, encapsulation mediating, and antibacterial activity, to confirm a pivotal role of LGBP in shrimp immunity in vivo. (Phupet et al., 2018). In this connection, previous studies in our laboratory have clearly shown that activation of plasma proPO system in P. viridis (Asokan et al., 1998) by laminarin resulted in the generation of certain intermediary opsonic factors leading to opsono-phagocytosis of yeast cells by hemocytes of P. viridis.
In this study, a slight, but statistically significant increase in the phagocytic uptake of yeast cells pretreated with laminarin alone compared to controls indicates the possibility of laminarin interaction with receptor/binding proteins on the surface of P. viridis hemocytes, leading to the release of opsonic molecules from the hemocytes. This contention also derives support from the observations of Asokan et al. (1997), demonstrating that pretreatment of P. viridis hemocyte monolayer with laminarin led to exocytotic release of hemocyte proPO system into the external milieu under in vitro conditions (data not shown). These observations also indicate the possible presence of β-GBP on the surface of hemocytes, which needs further elucidation.
Thus, this study shows that plasma β-GBP can function as an opsonin for hemocytes of a bivalve mollusc P. viridis by increasing phagocytic rate of trypsinized yeast cells. The opsonization with β-GBP was probably a direct binding of the molecules to β-1,3 glucan on the cell wall of yeast cells (Thörnqvist et al., 1994), as β-GBP previously reacted with laminarin failed to show enhancement in the phagocytic activity. In the latter case, β-GBP that had reacted with soluble β-1,3 glucans was probably blocked and unable to bind the yeast cell wall β-1,3 glucan. Furthermore, although β-1,3 glucan itself showed stimulation of limited phagocytic activity, it did not significantly enhance the phagocytic activity.
Preincubation of hemocytes of P. viridis with β-GBP—laminarin seems to block a stimulation of the phagocytosis with β-GBP. This is in accordance with results on crayfish 76-kDa protein that appears to be the same as β-GBP receptors (Renwrantz and Stahmer, 1983). Similarly, a significant reduction of β-GBP enhanced the phagocytic rate of trypsinized yeast cells pretreated with glucose or polyclonal antibody raised against β-GBP, provided an additional support to the fact that plasma β-GBP indeed serves as an opsonic molecule, and enhanced the phagocytic rate of trypsinized yeast cells by hemocytes since trypsinized yeast cells' surface is rich with polymers of β-1,3 glucans, and thus, β-GBP can possibly bind with β-1,3 glucans and recognize nonself molecules.
Although there are reports indicating that possibly, hyalinocytes or agranulocytes are involved in phagocytosis of yeast cells by crayfish hemocytes (Duvic and Soderhall, 1992), the specific hemocyte morphotype involved in opsono-phagocytosis of trypsinized yeast cells mediated by β-GBP in P. viridis requires further elucidation.
In conclusion, we show that the opsonic β-GBP from P. viridis plasma described in this study corresponds to phagocytosis-stimulating activities detected earlier in hemolymph from various invertebrates (Thörnqvist et al., 1994) and that this protein is essential in molluscan cellular defense. Based on the results of our previous and present studies, molluscan β-GBP appears to be a multipotential protein, wherein it is involved in proPO activation, agglutination reactions, partial degranulation, and release of molecules exocytotically, as well as its vital role in enhancing phagocytic responses of P. viridis hemocytes.
Footnotes
Disclosure Statement
There is no conflict of interest between the authors.
Funding Information
We gratefully acknowledge the financial support through the major research grant form Department of Science and Technology (DST), Government of India (Ref. No. SR/SO/AS-08/2008) and University Grants Commission (UGC) New Delhi for UGC-SAP – DSA-I (Ref. No. F.4 – 4 – 2007 (SAP-II) Dated 12 March 2007).