
Abstract
The effector proteins of several pathogenic bacteria contain the Glu-Pro-Ile-
Introduction
Following the discovery of the Glu-Pro-Ile-Tyr-Ala (EPIYA) motif in Helicobacter pylori's Cytotoxin-associated gene A (CagA) effector protein, several studies have shown that this protein has a substantial role in diseases arising after infection with pathogenic bacteria. Also, tyrosine phosphorylation of host cell proteins facilitates the bacterial invasion of nonphagocytic epithelial cells (Rosenshine et al., 1992). In Haemophilus ducreyi, the EPIYA effector protein lipoprotein signal peptidase A has been found to be essential in the promotion of ulcers in a rabbit infection model (Ward et al., 2003). In addition, it has been shown that neutralizing the effector protein ankyrin A can abolish Anaplasma phagocytophilum's ability to infect host cells (Lin et al., 2007).
These proteins are delivered into host cells by type III and IV secretion systems, which allow penetration of the plasma membrane and the direct transfer of bacterial molecules into the cytoplasm of targeted cells. Based on proteomic analyses, it has been shown that these motifs atypically interact with a wide range of Src homology 2 (SH2) domain-containing mammalian proteins through tyrosine phosphorylation (Selbach et al., 2009).
Moreover, in other investigations, it has been demonstrated that these bacterial proteins containing the EPIYA motif can function as pathogenic master keys that lead to the perturbation of multiple signaling cascades in the host by irregular interaction and by binding with SH2 domain-containing proteins (Backert et al., 2010). In the other words, bacterial EPIYA motifs (or like motifs) enable to interact with multiple host proteins because of their structural flexibility (Safari, 2014; Hayashi et al., 2012). Interestingly, it has also been reported that EPIYA (or EPIYA-like) motifs are present in mammalian proteomes, although not in high levels. Improper interactions between these motifs and the SH2 domain-containing proteins may be detrimental for mammalian cells. It should note that EPIYA motifs (or like motif) in mammalian proteins are often found in regions with ordered nature and it suggests that they are nonfunctional (Safari, 2014). In fact, the number of mammalian proteins with functional EPIYA motif (or like motif) is low.
The critical roles of functional EPIYA motif (or like motif) in mammalian proteins to perturb cell transduction pathways leading to increased risk of cancer progression were shown by several studies (Safari et al., 2011; Senda et al., 2016). As an illustration of such harmful connections, consider the many CagAPY SH2-domain contacts that are shown in H. pylori-induced actin-cytoskeletal rearrangements, scattering, and elongation of infected host cells in culture (Backert et al., 2010; Selbach et al., 2009). So far, only 13 bacterial EPIYA effector proteins have been recognized (Table 1). One of the key proteins involved in this process is CagA, which is transported into epithelial cells. Within CagA, specific tyrosine residues located within the EPIYA motifs can undergo phosphorylation by the Src family kinases (SFKs) or by c-Abl kinase. The number of EPIYA motifs can vary within the protein's variable region. It has been observed that the strains of H. pylori that possess CagA with a higher number of these repeats are more strongly associated with the development of gastric cancer than are the strains that possess CagA with a lower number of repeats.
Pathogenic Bacterial Effectors Containing EPIYA (or EPIYA-Like) Motifs and Related Host Src Homology 2 (SH2) Domain-Containing Proteins
CSK, C-terminal Src kinase; SFKs, Src family kinases; PI3K, Phosphatidylinositol-3 kinase; SHP2, SH2 domain-containing protein tyrosine phosphatase2; SHP1, SH2 domain-containing phosphatase1; GRB2, Growth factor receptor-bound protein 7; CRK, CT10 regulator of kinase; NCK, Noncatalytic region of tyrosine kinase adaptor protein; CBL, Casitas B-lineage lymphoma; RASGAP1, Rac GTPase-activating protein 1; ND, not detected.
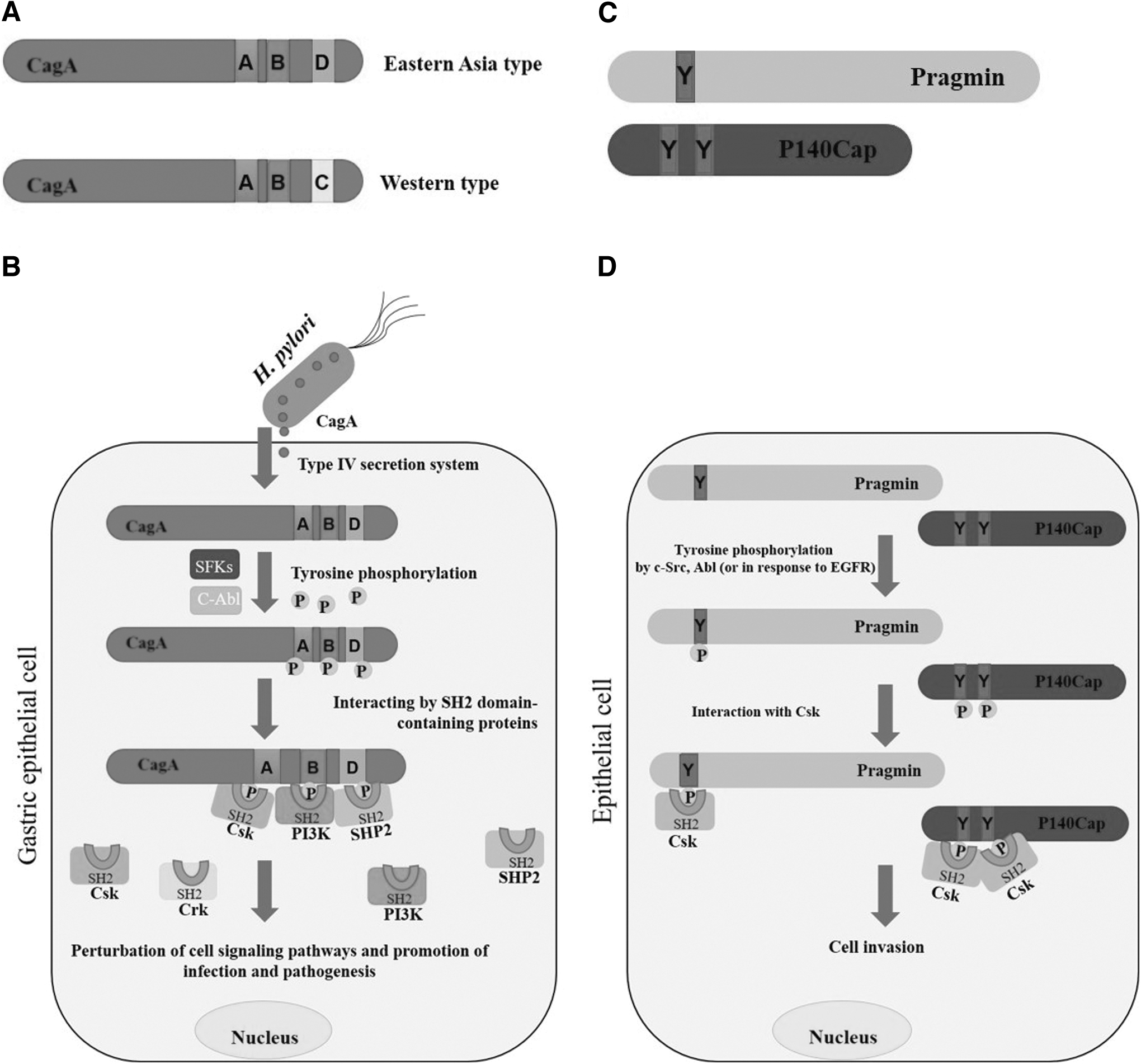
Phosphorylated CagA triggers the elongation of epithelial cells, and the extent of this effect is dependent upon the number of EPIYA motifs within the variable region (Fig. 1A, B) (Argent et al., 2005). Additionally, the absence of SH2 domain-containing proteins in the bacterial proteome suggests that bacteria never engage in the tyrosine phosphorylation-dependent interaction of EPIYA effector proteins. Some in vitro experiments have found that CagA-positive H. pylori strains' infection of gastric epithelial cells induces a specific morphological alteration characterized by cell elongation. This morphological change, which is known as the hummingbird phenotype, is associated with enhanced cell motility and scattering, and this morphology is modulated by SH2-containing protein tyrosine phosphatase 2 (SHP2). However, these bacterial effectors are capable of carrying out a variety of interactions with proteins in mammals that include SH2 domains. This finding suggests the existence of at least one mammalian EPIYA-containing protein, whose activities are used by bacterial effectors (Hayashi et al., 2013).
The implications of the presence of EPIYA-containing proteins in mammalian signaling pathways are multifaceted. These proteins have the potential to interact with key signaling cascades, modulate cellular processes, and contribute to the development and progression of various diseases. Therefore, understanding the structural and functional aspects of EPIYA-containing proteins in mammals is essential for unraveling the complexity of their signaling mechanisms and their role in cellular homeostasis and disease pathophysiology. Interestingly, the presence of 13 motifs similar to EPIYA, including EDLYA, EGLYA, EHIYD, ENIYE, EPLYA, EPIYG, EPIYD, EPVYA, ESIYA, ESIYE, EVVYA, TPLYA, and VNPYA, has been confirmed in proteins within the mammalian proteome through experimental approaches and bioinformatics analyses (Safari, 2014). These findings have unveiled intriguing possibilities regarding the involvement of these motifs in mammalian signaling pathways and their potential impact on cellular processes.
In this review, we focus on the regulation of cellular-signaling pathways by mammalian proteins containing bacterial EPIYA or EPIYA-like motifs predicted to be phosphorylated.
Prediction of Mammalian Proteins with EPIYA and EPIYA-Like Motifs
In a previous work (Safari, 2014), we used the National Center for Biotechnology Information's Basic Local Alignment Search Tool and Phosphosite software to identify mammalian proteins containing EPIYA (or EPIYA-like) motifs that are predicted to be phosphorylated. Our analysis provides valuable insights into the functional significance of these proteins in cellular communication and signaling networks (Table 2).
Mammalian Proteins Containing Bacterial EPIYA (or EPIYA Like) Motifs (Which Was Predicted to be Phosphorylated) and Their Functions in Related Cellular-Signaling Pathways
EPIYA
Pragmin
Pragmin (SgK223) is linked to Rho-related GTPase 2 (Rnd2), which is primarily present in neurons. Pragmin's binding affinity for Rnd2 depends on the presence of GTP and considerably enhances the activity of Ras homolog family member A (RhoA), triggering cellular contraction through the Rho-kinase pathway in HeLa cells. In PC12 cells, Pragmin impedes nerve growth factor-triggered neurite outgrowth in response to Rnd2. Thus, Pragmin plays a substantial role in the regulation of SFK activity and of cellular processes associated with Rnd2 (Tanaka et al., 2006). Additionally, Pragmin is a mammalian protein with active EPIYA sequences in its N-terminal region. Studies have shown that it can undergo tyrosine phosphorylation at these EPIYA sequences through the action of SFKs or C-terminal Src kinase (Csk) or in response to external signals like those from epidermal growth factor (EGF).
Pragmin can interact with the SH2 domain of Csk, a particular negative regulator of SFKs, thanks to this phosphorylation. The Csk–Pragmin connection prevents membrane-associated SFKs from becoming inactivated as a result of Csk's translocalization from the cytoplasm to the cellular membrane.
Therefore, in cells where Pragmin is phosphorylated at the EPIYA motif, SFK activity is maintained (Safari et al., 2011) (Fig. 1C, D). The effects of Pragmin and Src tyrosine kinase activity on cell morphological changes in AGS cells include cell elongation and an invasive phenotype (Safari and Zayer, 2018). Moreover, it has been found that, although Src tyrosine kinase activity is crucial for controlling cell movement, the changes in cell shape induced by Pragmin in AGS cells are independent of Src activity. This suggests the involvement of alternative mechanisms in this process.
In cancer, increased Pragmin expression is observed in different adenocarcinoma cells, and this elevated expression contributes to the promotion of cell migration and invasion. In the case of gastric adenocarcinoma cells, Pragmin expression triggers cell migration through a mechanism that relies on the presence of Csk (Safari and Shokohfard, 2020; Safari and Zaer, 2017; Senda et al., 2016). A 2009 study conducted by Leroy et al. identified human Pragmin as an essential element in oncogenic signaling facilitated by the tyrosine kinase Src. The research revealed that Pragmin plays a crucial role in promoting tumor cell growth and invasion (Leroy et al., 2009).
The α subunit of general transcription factor TFIIE
The α subunit of transcription factor II E (TFIIE) has been reported as an additional protein containing the EPIYA motif (Safari, 2014). TFIIE is a key component of the preinitiation complex involved in transcription initiation and interacts with various proteins, including unphosphorylated RNA polymerase II, transcription factor II B, and both subunits of transcription factor II F.
TFIIE plays a crucial role in recruiting and regulating the activities of transcription factor II H, which includes its kinase and helicase functions. The interaction between TFIIEα and transcription factor II H is specifically mediated by the alpha subunit of TFIIE. The regions responsible for this interaction have been identified in the conserved zinc-finger domain and the acidic carboxyl terminus of TFIIEα (Jawhari et al., 2006; Orphanides et al., 1996). Furthermore, a research has demonstrated that TFIIH kinase module and TFIIEα are released from the promoter concurrently with RNA polymerase II phosphorylation and DNA opening. Remarkably, mutations in TFIIE and TFIIH linked to trichothiodystrophy, a developmental disorder, impact these early transcriptional events (Compe et al., 2019).
Furthermore, TFIIEα and p53 have a similar binding location on the Tfb1/p62 subunit of TFIIH, suggesting that TFIIEα may be involved in signaling pathways and that this may have ramifications for cancer (Di Lello et al., 2008).
EPLYA
P140Cap
P140Cap is a docking protein. In a 2013 investigation, Repetto et al. (2013) demonstrated that the N-terminal segment of p140Cap, also known as SRC kinase signaling inhibitor 1, possesses 2 EPIYA-like motifs: EPLYA and EGLYA. These motifs can undergo tyrosine phosphorylation that is mediated by c-Abl or that occurs in response to integrin-mediated adhesion and EGF stimulation. Subsequently, the phosphorylated EPLYA and EGLYA sequences act as binding sites for the SH2 domain of Csk (Repetto et al., 2013).
The signaling role of p140Cap, a novel Src-binding protein, involves its C-terminal region, which contains a proline-rich sequence facilitating p140Cap's interaction with the Src homology 3 domain. This interaction enables Csk's inhibition of Src kinase activity. Silencing p140Cap increases cell spreading, cell migration rates, and Src kinase activity. Conversely, elevated expression of p140Cap activates Csk, leading to the inhibition of Src and downstream signaling pathways and the modulation of cell motility and invasion (Di Stefano et al., 2007) (Fig. 1C, D). P140Cap also affects the activity of Rac1 GTPase by interacting with Tiam1, leading to decreased Rac activation.
It forms a complex with Tiam1 and E-cadherin in human EGF receptor 2-positive tumor cells, potentially impacting cell adhesion. Additionally, p140Cap upregulates E-cadherin expression, which is typically downregulated during the metastatic process and inhibits the expression of epithelial-to-mesenchymal transition (EMT) transcription factors and N-cadherin. Further research is needed to understand the precise mechanisms and temporal–spatial interactions of p140Cap in these processes and its impact on tumor progression.
In neuroblastoma, p140Cap protein plays a negative regulatory role in the Src kinase activation and tyrosine phosphorylation of p130Cas, as well as in the phosphorylation of signal transducer and activator of transcription 3 (STAT3) and its upstream kinase, Janus kinase 2 (JAK2). The overexpression of Src kinase and the activation of STAT3 has been associated with poor outcomes in patients with neuroblastoma, while their inhibition has been shown to decrease proliferation and enhance apoptosis in neuroblastoma cells. P140Cap-expressing neuroblastoma cells exhibit increased sensitivity to chemotherapy drugs, particularly doxorubicin and etoposide, which induce DNA damage.
These cells also display increased sensitivity to combination chemotherapy with Src inhibitors, suggesting a synergistic effect on decreasing neuroblastoma cell viability. The ability of p140Cap to interact with proteins from multiple pathways contributes to the increased sensitivity of neuroblastoma cells to combined treatments (Salemme et al., 2021). Recently, it was reported that p140Cap inhibits β-Catenin in the breast cancer stem cell (CSC) compartment instructing a protective antitumor immune response (Salemme et al., 2023).
ESIYE motif of retinal dehydrogenase 2
Retinaldehyde dehydrogenase 2 (RALDH2 or also known as ALDH1A2) catalyzes the synthesis of retinoic acid from retinaldehyde and the importance of retinoic acid in signaling was reported. A study has shown that RALDH2 plays a substantial role in modulating the fibroblast growth factor (FGF) and Notch signaling pathways. The research revealed that a mid-gestation deficiency of retinoic acid, specifically of RALDH2, leads to structural and molecular deficiencies in the dorsal regions of the spinal cord in mice. These deficiencies result in notable reductions in FGF and Notch signaling. Furthermore, the findings highlighted the importance of endogenous retinoic acid, regulated by RALDH2, in driving stem cell differentiation.
The impact of RALDH2 on FGF and Notch signaling emphasizes its crucial role in maintaining an appropriate signaling environment during spinal cord development, which ultimately influences proper neural stem cell differentiation. Moreover, the importance of RALDH2 in several diseases was shown (Beecroft et al., 2021; Leon et al., 2023; Paschaki et al., 2012; Ruiter et al., 2021; Sanders et al., 2021; Zhang et al., 2022). The role of the ESIYE motif of RALDH2 has not been determined.
EDLYA
Myotubularin-related protein 6
The myotubularin family, a group of inositol polyphosphate 3-phosphatases, consists of 16 different proteins. Nine of these proteins exhibit catalytic activity, allowing them to dephosphorylate phosphatidylinositol 3-phosphate [PtdIns(3)P] and phosphatidylinositol 3,5-bisphosphate [PtdIns(3,5)P2] at the D-3 position. The remaining seven members lack catalytic activity due to the absence of a critical cysteine residue in the CX5R motif. In a study conducted by Zou et al. (2012), the focus was on a subgroup of related myotubularins, which includes myotubularin-related protein 6 (MTMR6), MTMR7, and MTMR8. These proteins form dimers with the catalytically inactive MTMR9. When active myotubularins interact with MTMR9, their catalytic activity is enhanced, and their substrate preferences are altered. Specifically, the MTMR6/R9 complex favors PtdIns(3,5)P2 as its substrate, while the MTMR8/R9 complex prefers PtdIns(3)P.
MTMR9 significantly increases the enzymatic activity of MTMR6 toward PtdIns(3,5)P2 but has a more modest effect on its activity toward PtdIns(3)P. In contrast, MTMR9 enhances both the activity of MTMR8 toward PtdIns(3,5)P2 and PtdIns(3)P.
Within cells, the MTMR6/R9 complex raises the levels of cellular PtdIns(5)P, a product of PI(3,5)P2 dephosphorylation, and this complex inhibits stress-induced apoptosis. It has been reported that the interaction between MTMR6 and guanosine diphosphate-bound Rab1B through the GRAM domain partly overlaps with the interaction of Rab1B in the pericentrosomal and peri-Golgi regions. The research also has suggested that Rab1B plays a role in regulating the localization of MTMR6. Furthermore, the reduction of either MTMR6 or Rab1B inhibits the formation of tubular omegasomes induced by the overexpression of double FYVE-containing protein 1 in autophagy. These findings provide evidence that MTMR6 is regulated by Rab1B through the conserved GRAM domain in the early secretory and autophagic pathways, proposing a novel mechanism for the regulation of myotubularin phosphatase by Rab1B (Mochizuki et al., 2013). In line with these findings, in 2005, Srivastava et al. demonstrated that KCa3.1 is regulated by phosphatidylinositol 3-phosphate (PI[3]P) and that MTMR6 inhibits KCa3.1 by dephosphorylating the 3′ position of PI(3)P.
This action potentially leads to decreased PI(3)P levels in the lipid microdomains adjacent to KCa3.1. KCa3.1 plays a crucial role in controlling proliferation in T cells, vascular smooth muscle cells, and certain cancer cell lines. Collectively, these data suggest an intricate regulatory network that involves the interplay among MTMR6, PI(3)P, and KCa3.1 and that contributes to the modulation of proliferation in various cellular contexts (Mochizuki et al., 2013; Srivastava et al., 2005). Moreover, an immunohistochemistry analysis has demonstrated a notable increase in the expression of MTMR6 in ovarian cancer tissues compared with adjacent tissues. This finding further supports the significance of MTMR6 in ovarian cancer and strengthens the potential association between MTMR6 dysregulation and tumorigenesis (Wang et al., 2019). The role of the EDLYA motif of MTMR6 has not been determined.
ENIYE
Sodium/hydrogen exchanger 9
Na+/H+ Exchanger 9 (NHE9) is an endosomal membrane protein encoded by the Solute Carrier 9A, member 9 gene (SLC9A9) that works to regulate intracellular pH, sodium levels, and cell volume. In a study by Beydoun et al., ectopic expression of NHE9 (or also known as SLC9A9) in human brain microvascular endothelial cells (hBMVECs) without external cues upregulated transferrin receptors (TfRs) and downregulated ferritin, leading to increased iron uptake. The researchers demonstrated that NHE9 localized to recycling endosomes in the hBMVECs and raised the endosomal pH. This alkalization of the endosomal lumen increased the translocation of TfRs to the hBMVEC membranes.
TfRs on the membrane play a role in recycling-dependent and recycling-independent iron uptake. The findings indicate that NHE9 fine-tunes the endosomal pH in response to paracrine signals, regulating TfR-dependent, recycling-independent iron uptake and the iron mobilization pathway at the blood–brain barrier (Beydoun et al., 2017). In another study, it was found that NHE9 prevented cisplatin resistance-associated gene-induced apoptosis.
The NHE9-dependent induction of CRT resistance involves the activation of RACK1-associated Src/Akt/-catenin signaling, the researchers found. They also identified that receptor for activated C kinase 1 (RACK1) is a new binding partner of NHE9. Additionally, it was discovered that cells with NHE9-induced CRT resistance have increased Bcl-2 protein. These findings highlight the potential of NHE9 as a predictor of CRT response and its relevance in the development of targeted therapies for CRT-resistant esophageal squamous cell carcinoma (Chen et al., 2015).
Furthermore, Zhang-James et al. (2011) demonstrated in their study that the NHE9 C-terminal interacts with signaling molecules such as calcineurin homologous protein and RACK1. These molecules, along with other binding partners of Na+/H+ exchangers, play roles not only in regulating Na+/H+ exchanger activity but also in regulating the calcium-signaling and protein phosphorylation pathways. These pathways are crucial for synaptic transmission and the plasticity of synapses (Zhang-James et al., 2011). Regarding the role of NHE9 in cancer, a study in patients with glioblastoma multiforme found that high expression of NHE9 in tumors was associated with reduced survival and a poor response to neoadjuvant chemoradiotherapy. The researchers also observed that increased NHE9 expression in brain tumor-initiating cells led to enhanced migration, endosome alkalization, and transferrin uptake (Kondapalli et al., 2015). Similarly, Ueda et al. reported that colorectal tumors with elevated NHE9 expression exhibited high rates of liver metastasis and poor prognoses.
They also identified a positive association between NHE9 and EGF receptor (EGFR). An in vitro analysis revealed that overexpression of NHE9 in colorectal tumors promoted anchorage-independent growth and proliferation (Ueda et al., 2017).
In the context of esophageal cancer, a gene expression analysis of biopsied tissue from patients with squamous cell carcinoma found substantial upregulation of NHE9, which predicted the patients' responses to neoadjuvant chemoradiotherapy treatment (Wen et al., 2014). In a follow-up study, it was demonstrated that upregulated NHE9 conferred resistance to chemoradiotherapy in esophageal squamous cell carcinoma by upregulating the Bcl-2 prosurvival pathway. Furthermore, in a separate study, researchers examined the methylation status of NHE9 in oral squamous cell carcinoma and observed a transition from hypomethylation in the earlier stages of cancer to hypermethylation as cancer progressed (Krishnan et al., 2016). Moreover, it was shown that NHE9 regulates phagosome maturation and bactericidal activity in macrophages (Shamroukh et al., 2022).
MICAL-like protein 2
Molecules interacting with CasL (MICAL)-like protein 2 (MICAL-L2) is a member of the MICAL family with roles in the regulation of cytoskeleton dynamics. In gastric cancer cells, increased expression of MICAL-like protein 2 (MICAL-L2) has been found to upregulate EGFR protein levels, leading to enhanced cell migration. Conversely, silencing MICAL-L2 downregulated EGFR and inhibited cell migration. The activation of the heat shock protein 27 (HSP27)/cytoskeleton and HSP27/β-catenin-signaling pathways was positively correlated with MICAL-L2 expression, indicating their involvement in controlling cell migration. MICAL-L2 exerted its upregulating effect on EGFR through a transcription-independent mechanism by inhibiting EGFR protein degradation in lysosomes. Additionally, cell division control protein 42 homolog (Cdc42) activation helped maintain the stability of EGFR in the presence of MICAL-L2. An analysis of clinic specimens further confirmed increased expression of MICAL-L2 in carcinoma tissues, along with a positive correlation between MICAL-L2 and EGFR expression levels.
These findings suggest that MICAL-L2 enhances gastric cell migration by inhibiting EGFR degradation in lysosomes through a Cdc42-dependent mechanism, thereby activating EGFR/HSP27-signaling pathways (Min et al., 2019).
In ovarian cancer cells, silencing MICAL-L2 suppressed cell proliferation, migration, and invasion. It also prevented the nuclear translocation of β-catenin, inhibited canonical Wnt/β-catenin signaling, and induced mesenchymal–epithelial transition. These observations indicate that MICAL-L2 serves as an important regulator of EMT in ovarian cancer cells (Zhu et al., 2015). Furthermore, MICAL-L2 has been found to physically interact with the protein cellular Myc (c-Myc), maintaining its nuclear levels and prolonging its half-life. Knockdown of MICAL-L2 expression resulted in decreased c-Myc protein stability through accelerated polyubiquitylation and degradation. MICAL-L2 has also been shown to deubiquitinate c-Myc, inhibiting its degradation and potentially blocking its phosphorylation at threonine residue 58 (Min et al., 2021). Overall, these studies demonstrate the diverse roles of MICAL-L2 in different cancer types. In gastric cancer, MICAL-L2 promotes cell migration through its interaction with EGFR and modulation of EGFR degradation. In ovarian cancer, MICAL-L2 helps to regulate EMT and canonical Wnt/β-catenin signaling.
Additionally, MICAL-L2 affects the stability and degradation of c-Myc in a phosphorylation-dependent manner. These findings highlight the significance of MICAL-L2 in cancer progression and provide potential avenues for therapeutic intervention.
EPVYA
Palmdelphin
Palmdelphin (PALMD) belongs to the family of paralemmin proteins implicated in cytoskeletal regulation. It plays a crucial role in multiple cellular processes and signaling pathways. It associates with TNFAIP3-interacting protein 1 (TNIP1), a binding protein of tumor necrosis factor alpha-induced protein 3 (TNFAIP3) and inhibitor of nuclear factor kappa B kinase regulatory subunit gamma (IKBKG), in NF-κB signaling. The loss of PALMD disrupts TNFAIP3-dependent deubiquitinating activity, leading to increased ubiquitination of IKBKG and the subsequent activation of NF-κB. Key characteristics of calcific aortic valve disease, such as thicker aortic valve leaflets, increased aortic valve peak velocity, and excessive extracellular matrix deposition, are seen in elderly Palmd-deficient mice. Significantly, PALMD overexpression mediated by an adeno-associated virus reduces aortic valvular remodeling in mice suffering from calcific aortic valve disease, suggesting a protective effect (Han et al., 2023). A splice variant of PALMD, known as PALMD-KKVI, is expressed in endothelial cells.
It interacts with Ran GTPase-activating protein 1 (RANGAP1), a regulator of the GTPase RAN, and disrupts nucleocytoplasmic shuttling through exportin 1 (XPO1). The reduction of PALMD expression alters the subcellular localization of RANGAP1 and XPO1, causing the nuclear arrest of XPO1 cargoes, including p53 and p21. This disrupts gene regulation by affecting the localization of transcriptional regulators. The loss of PALMD in endothelial cells leads to the cells' failure to form a perinuclear actin cap, which normally protects against mechanical stress; this results in the misalignment of the nuclear long axis relative to the cell body. PALMD is essential for nucleocytoplasmic transport, gene regulation, and responsiveness to mechanical stress in endothelial cells (Sáinz-Jaspeado et al., 2021).
A research has demonstrated that PALMD is a proapoptotic gene that is specifically phosphorylated serine-46-induced by p53. In cells transfected with wild-type p53, PALMD was upregulated, but not in cells with a serine-46 mutation. In response to DNA damage, p53 induced the expression of PALMD. It is interesting to note that ATP depletion caused necroptosis-like cell death when PALMD was downregulated.
To cause apoptosis, PALMD primarily accumulated in the nucleus following DNA damage. According to these results, serine-46-phosphorylated p53 targets PALMD, which regulates cell death in response to DNA damage (Dashzeveg et al., 2014).
Regarding PALMD's role in cancer, abnormal genetic changes have been observed in malignant tumors. Additionally, low levels of PALMD have been associated with the metastasis of uveal melanoma, indicating a broader involvement in cancer pathogenesis (Cai et al., 2022). The role of the EPVYA motif of PALMD has not been determined.
Rho GTPase-activating protein 27 isoform
In the intricate landscape of cancer-signaling pathways, the Rho GTPase-activating protein (ARHGAP27) takes center stage. Known for its specific activity toward Ras-related C3 botulinum toxin substrate 1 (Rac1) and Cdc42, ARHGAP27 has emerged as a potential oncogene with notable implications in cancer development (Sakakibara et al., 2004). Adding a layer of complexity, the cellular responses induced by transforming growth factor-beta (TGF-β) involve distinct signaling pathways that diverge at Smad3. Intriguingly, TGF-β-dependent signaling, mediated by Smad3's β4 region, leads to the downregulation of multiple mRNAs encoding GTPase-activating proteins targeting Rac1 (ARHGAPs).
This specific mechanism attenuates Rac1 inactivation and, remarkably, contributes to sustained Rac1 activation, ultimately enhancing cell motility. Thus, the convergence of TGF-β signaling and ARHGAP activity reveals a cooperative relationship downstream of the TGF-β type I receptor, playing a pivotal role in cancer progression (Motizuki et al., 2021). In epithelial ovarian cancer, ARHGAP27 emerges as a key player.
Elevated expression levels of ARHGAP27 in this cancer type suggest its potential as a candidate gene influencing tumor initiation and development (Permuth-Wey et al., 2013). Notably, the intricate regulatory network extends to the ARHGAP27 promoter, where two methylation sites are linked to a lower incidence of epithelial ovarian cancer and a corresponding decrease in ARHGAP27 expression. Further insights into ARHGAP27's multifaceted role come from studies involving rat glioma cells overexpressing CD133, a cell surface marker associated with CSCs. This overexpression correlates with enhanced stem cell characteristics, increased migration, proliferation, and higher expression of ARHGAP27.
Silencing ARHGAP27, on the other hand, results in reduced invasion, suggesting a potential involvement of ARHGAP27 in the migration of cancer cells (Kreider-Letterman et al., 2022). Further investigations are required to establish a comprehensive understanding of the precise role of ARHGAP27 in cancer and its implications for epithelial ovarian cancer in particular. The role of the EPVYA motif of ARHGAP27 has not been investigated.
EHIYD
Occludin isoforms
Occludin is a tight junctional strand protein that interacts with scaffolding proteins to maintain the integrity of tight junctions. According to one study, occludin plays a crucial role in signaling processes. The research revealed that suppressing occludin leads to changes in claudin levels in epithelial cells and influences the cytoskeleton and the function of tight junctions. Additionally, the study discovered that occludin acts as a mediator for external and intramembrane signals through the Rho-signaling pathway, triggering the reorganization of the actin cytoskeleton. In the absence of occludin, impaired signaling occurs, potentially disrupting the dynamic behavior of tight junction strands. This disruption can result in the tight junction's increased permeability to large organic cations (Yu et al., 2005). A noteworthy discovery in human lung cancer patient samples revealed a significant increase in occludin protein expression. To delve deeper into this observation, researchers conducted studies involving the knockdown of occludin.
Remarkably, the results demonstrated that occludin knockdown not only inhibited lung cancer cell proliferation both in vitro and in vivo, but also had a profound impact on apoptosis and invasion abilities. Mechanistically, the study shed light on the compromised activity of the key growth pathway AKT/PI3K following occludin knockdown. Additionally, the expression of apoptosis-related proteins, including BAX, caspase-3, caspase-9, and AIF, exhibited an upregulation, while Bcl-2 remained unaffected (Wang et al., 2018). Extending beyond lung cancer, occludin's involvement in various malignancies becomes evident. In gastric cancer, occludin is linked to increased expression levels of matrix metalloproteinases (MMP-2 and -9), thereby enhancing cancer cell invasion. In nasopharyngeal carcinoma, occludin overexpression is specifically associated with advanced disease stages. Meanwhile, in glioma, occludin downregulation correlates with increased permeability and the suppression of zonula occludens-1 (ZO-1) (Bhat et al., 2019). The role of the EHIYD motif of occludin has not been investigated.
EPIYA (or EPIYA-Like)-Motif Mammalian Proteins as Potent Targets in Cancer Therapy
It has been found that the expression of EGFR, Csk, Src, and Pragmin was inhibited by the secretome of human amniotic mesenchymal stromal cells (hAMSCs) in the MiaPaca2 cell line, leading to suppression of tumor cell growth (Rahmani and Safari, 2021). Another study showed that hAMSCs in Panc1 cancer cells inhibit tyrosine phosphorylation of SgK223 (at the Y411 site) (Safari et al., 2022). These results indicate that the expression of SgK223 (or tyrosine phosphorylation of SgK223 at the Y411 site) plays a critical role in the suppression of tumors.
Clinical Effects of EPIYA (or EPIYA-Like) Motifs on Various Cancers
Given that H. pylori infection is among the most significant risk factors for gastric cancer, several studies have investigated the relationship between H. pylori infection (specifically with strains producing the CagA toxin) and the progress of gastrointestinal diseases. In one of these studies, associations between cagA genotype and clinical outcomes in the Iranian population were examined. The researchers found a direct relationship between the cagA genotypes ABCC and ABCCC and a number of clinical outcomes of infection, such as peptic ulcers and gastric cancer. The authors also concluded that there is a substantial correlation between the number of copies of EPIYA-C and the development of gastric cancer. Based on those results, it appears that the EPIYA-ABCCC motif has a substantial positive relationship with gastric cancer in the Iranian population (Keikha et al., 2021; Karbalaei et al., 2021).
Furthermore, in a study investigating the EPIYA motif in the Colombian population, researchers found a direct relationship between the number of EPIYA-C repeats and patients' ages; a considerable number of EPIYA ABCC and ABCCC repeats were found in patients older than 50 years of age (Gómez et al., 2020). Also, in the Mexican population, a study found that, in patients from southern Mexico who had chronic gastritis, the H. pylori strains with the vacA s1 m1, CagA+, and babA2− genotypes were the most common. In the cagA+ strains, the EPIYA-ABC motif was the most prevalent (Atrisco-Morales et al., 2018). It has previously been reported that, in Southeast Asians, patients infected with vacA m1 or cagA-positive H. pylori strains are more likely to develop peptic ulcer disease (Sahara et al., 2012). Additionally, a recent investigation in the Amazon region of Brazil showed that many EPIYA-C motifs were associated with intestinal metaplasia.
Although this study also found that infection with H. pylori strains with more than 1 EPIYA-C motif in the cagA gene were associated with intestinal metaplasia and gastric adenocarcinoma, neither neutrophil activity nor inflammation was observed in patients with these strains (Vilar e Silva et al., 2014). On the other hand, a study of patients in northeast Thailand demonstrated that there is no direct association between increased numbers of EPIYA-C motifs and worsening clinical outcomes. The most prevalent CagA type observed among northeastern Thai patients with dyspepsia was the Western CagA type, which is associated with peptic ulcer disease and which may be a predictive parameter for adverse clinical outcomes (Chomvarin et al., 2012).
Among mammalian proteins with bacterial EPIYA motif (or like motif), only in Pragmin (or SgK223) and p140Cap, the functional roles of their EPIYA motifs (or like motifs) were detected in various cancer cells. Pragmin (or SgK223) showed similarity sequence (or function) with oncoprotein CagA of H. pylori (segment B). However, EPIYA motif of Pragmin interacts specifically with CSK (Safari et al., 2011). Moreover, EPIYA-like motifs of p140Cap were reported to interact with CSK (Repetto et al., 2013). Of note, some studies showed that overexpression of SgK223 promotes the cell migration and cell morphological changes (invasive form) in different cancer cells (Safari et al., 2011; Senda et al., 2016; Tactacam et al., 2015). However, the related mechanisms were not fully understood. It seems that EPIYA motifs (or like motif) in mammalian proteins are often found in regions with ordered nature and thereby, they are nonfunctional. However, bacterial EPIYA motifs (or like motif) are mostly functional and enable to interact with multiple host proteins because of their structural flexibility (Hayashi et al., 2012; Safari, 2014).
Taken together, it seems that our current studies about functional EPIYA motifs (or like motif) in mammalian proteins is low and thereby, more studies will be required to confirm their harmful effects in mammalian cells.
Conclusions
Mammalian proteins with bacterial EPIYA (or EPIYA-like) motifs have the potential to interact with key signaling cascades, modulate cellular processes, and contribute to the development and progression of various diseases. However, the possible roles of these proteins have not been well investigated. Understanding the structural and functional aspects of EPIYA-containing proteins in mammals is essential to unraveling the complexity of their signaling mechanisms and their role in cellular homeostasis and disease pathophysiology. Notably, it has been shown that most of the EPIYA (or EPIYA-like) motifs in mammalian proteins occur naturally. Furthermore, a limited number of them are tyrosine-phosphorylated, and most of their EPIYA (or EPIYA-like) motifs are not functional. EPIYA (or EPIYA-like) motif mammalian proteins may be potent targets in the treatment of diseases such as cancer. Further studies are required.
Footnotes
Acknowledgments
The authors express their gratitude to the editors at MD Anderson Research Medical, in particular to Laura L. Russell, as well as to Pari Rohani, for their valuable editorial Scientific Editor assistance.
Authors' Contributions
Conceptualization: F.S.; writing/original draft preparation: M.R., F.S., M.A., and R.R.; and writing, review, and editing: M.R., F.S., M.A., N.S., and R.R. All authors have read and agreed to the published version of the article.
Disclosure Statement
The authors declare no competing financial interests.
Funding Information
No funding was received for this article.