
Abstract
Sjogren's syndrome (SS) is a complex systemic autoimmune disease. This study aims to elucidate a humanized NOD-PrkdcscidIl2rgem1/Smoc (NSG) murine model to better clarify the pathogenesis of SS. NSG female mice were adoptively transferred with 10 million peripheral blood mononuclear cells (PBMCs) through the tail vein from healthy controls (HCs), primary Sjogren's syndrome (pSS), and systemic lupus erythematosus (SLE) patients on D0. The mice were subcutaneously injected with C57/B6j submandibular gland (SG) protein or phosphate-buffered saline on D3, D17 and D31, respectively. NSG mice were successfully transplanted with human PBMCs. Compared with NSG-HC group, NSG-pSS and NSG-SLE mice exhibited a large number of lymphocytes infiltration in the SG, decreased salivary flow rate, lung involvement, decreased expression of genes related to salivary secretion, and the production of autoantibodies. Type I interferon-related genes were increased in the SG of NSG-pSS and NSG-SLE mice. The ratio of BAX/BCL2, BAX, cleaved caspase3, and TUNEL staining were increased in the SG of NSG-pSS and NSG-SLE mice. The expressions of p-MLKL and p-RIPK3 were increased in the SG of NSG-pSS and NSG-SLE mice. Increased expression of type I interferon-related genes, PANoptosis (apoptosis and necroptosis) were identified in the SG of this typical humanized NSG murine model of SS.
Introduction
Sjogren's syndrome (SS) is a systemic autoimmune disease that primarily affects the exocrine glands, primarily the salivary and lacrimal glands, causing severe dryness of the mucosal surfaces, primarily in the mouth and eyes (Fox, 2005). SS can be divided into two types (Yao et al., 2013): primary Sjogren's syndrome (pSS) and secondary SS. Secondary SS refers to SS occurring on the basis of connective tissue diseases such as systemic lupus erythematosus (SLE) or rheumatoid arthritis (RA). pSS is more common in middle-aged women, and its incidence is second only to RA among rheumatic immune diseases (Brandt et al., 2015; Qin et al., 2015). At present, the etiology of pSS is still unclear. Genetic, environmental, epigenetic, infection, autoimmune abnormalities, sex hormone levels, and other factors are involved in the pathogenesis (Ambrosi and Wahren-Herlenius, 2015; Imgenberg-Kreuz et al., 2018; Nocturne and Mariette, 2013).
Interferon (IFN) plays an important role in the pathogenesis of SS and is a potential therapeutic target (Bodewes and Versnel, 2018). Type I IFN has been considered as modulators of innate immune response against viral invasion (Huijser and Versnel, 2021; Marketos et al., 2019). The expression of type I IFN-induced genes is markedly elevated in the peripheral blood and salivary glands of patients with SS. The mRNA level of type I IFN-inducible genes in SS also showed positive correlation to the titers of SS-specific autoantibodies such as anti-Ro and anti-LA (Brkic and Versnel, 2014; Marketos et al., 2019; Yao et al., 2013). In addition, abnormal cell apoptosis is also crucial in the pathogenesis of pSS (Manganelli and Fietta, 2003; Nakamura et al., 2018; Sisto et al., 2022).
Several SS models have been developed, including induced models and genetic models (spontaneous development of SS-like disease in mice) (Gao et al., 2020; Scuron et al., 2019). NOD/ltj mice and salivary gland protein-induced experimental SS mice are widely used to study the pathogenesis of SS. However, sometimes these animal models do not fully mimic the clinical manifestations of SS, such as extraglandular manifestations, and secondary SS. NOD-PrkdcscidIl2rgem1/Smoc (NSG) (Walsh et al., 2017) is a severe immunodeficient mouse lacking T, B, and NK cells that can efficiently engraft human CD34+ hematopoietic stem cells (HSCs), peripheral blood mononuclear cells (PBMCs), cell line-derived allografts, patient-derived xenografts, or adult stem cells and tissues. The humanized mouse model of SLE has been successfully established in NSG mice to facilitate the exploration of pathophysiology and targets for therapy (Mihaylova et al., 2020; Wen et al., 2021).
In this study, a representative humanized SS animal model was successfully established by injecting NSG mice with PBMCs from patients with pSS and SLE. The mouse model corresponding to pSS patients is clinically pSS, whereas the mouse model corresponding to SLE patients is clinically secondary SS. Both primary and secondary SS murine models have decreased salivary flow rate, a large number of lymphocyte infiltration in the submandibular gland (SG), autoantibodies production, lung lesions, and decreased expression of genes related to salivary secretion. However, in the secondary SS model, there are lupus symptoms such as renal damage and anti-ds-DNA antibody production. In addition, type I interferon pathway-related gene expression, apoptosis, and necroptosis were significantly increased in the SG of this mouse model of SS.
MDA5 was mainly expressed in infiltrating lymphocytes and acinar epithelium of mouse SG, while ZBP1 was mainly expressed in ductal epithelial cells. In general, this humanized model of SS can better simulate the actual clinical situation of primary and secondary SS. By establishing this animal model of SS, we have provided a new way to study its pathogenesis.
Materials and Methods
Human samples
Patients diagnosed with pSS or SLE and fulfilling the respective classification criteria (Aringer, 2019; Vitali et al., 2002) were recruited from the Department of Rheumatology, Xiangya Hospital, Central South University. All experiments with human samples were approved by the Ethics Committee of Xiangya Hospital, Central South University (reference number: 2019030465). The subjects were informed about the collection of their specimen and signed an informed consent form. The clinical information of the pSS and SLE patients is shown in Tables 1 and 2.
Clinical Characteristics of Primary Sjogren's Syndrome Patients
Clinical Characteristics of Systemic Lupus Erythematosus Patients
The European League Against Rheumatism (EULAR) Sjogren's syndrome disease activity index (ESSDAI) is a commonly used clinical criterion for assessing the disease activity of SS, which includes the assessment of systemic damage of 12 systems related to pSS (Seror et al., 2015). A moderately active disease was defined as an ESSDAI ≥5. All the enrolled patients with pSS were in an active state of disease with an ESSDAI score greater than 5.
The SLE Disease Activity Index (SLEDAI) score is commonly used to assess disease activity in SLE (Bombardier et al., 1992). An active disease was defined as SLEDAI ≥5. All included SLE patients had SLEDAI scores greater than 5.
PBMCs collection
PBMCs were isolated from the blood of healthy controls (HCs), pSS, and SLE patients by Ficoll (GE Healthcare) density-gradient centrifugation according to the instruction.
Generation of humanized SS mice
C57/B6j mice 8-weeks old were purchased from the Hunan Slac Jingda Laboratory Animal Co., Ltd. (Changsha, China). Mice were maintained in a specific pathogen-free environment at the Department of Laboratory Animals, Central South University. All animal experiments were approved by the Ethics Committee for Animal Experiments of Central South University (reference number: 2020sydw0950). Female 8–10 weeks NSG mice (kindly provided by Shanghai Model Organisms) were adoptively transferred with 10 × 106 PBMCs from HCs (n = 4), active pSS (n = 5) and SLE (n = 5) patients on Day 0. Then the SS mouse model was induced by immunization with SG proteins as described (Lin et al., 2015). To prepare SG proteins, bilateral SG from wild-type C57/6j mice were collected, homogenized in phosphate-buffered saline (PBS), and centrifuged at 12,000 g for 5 min at 4°C.
The supernatant was collected and the protein concentration was determined by diquinolinic acid assay and emulsified to a concentration of 2 mg/mL by the addition of an equal volume of Freund's complete adjuvant (Sigma-Aldrich). For pSS and SLE groups, each mouse received subcutaneous multi-injections on the back with 0.1 mL of the emulsion on days 3 and 17, respectively. On day 31, the booster injection was carried out with a dose of 1 mg/mL SG proteins emulsified in Freund's incomplete adjuvant (Sigma-Aldrich). For HC group, each mouse received subcutaneous multi-injections on the back with equal PBS. Eight weeks later, all mice were harvested. The experiment was repeated twice.
Saliva and blood collection
After the mice were anesthetized, pilocarpine (Selleck, 5 mg/kg) was injected intraperitoneally to induce salivary secretion (Lin et al., 2011). Stimulated whole saliva was collected by weighing from the oral cavity using a 200 μL pipette tip for 15 min at room temperature. Blood samples were collected for serum preparation.
Histological assessment
According to standard procedures, SG, lung, and kidney tissues were embedded in paraffin, and sections were stained with hematoxylin & eosin. A widely used scoring system was used to assess the severity of SG damage based on the size of the infiltrate and the extent of lymphoid tissue (Scardina et al., 2007).
Flow-cytometric analysis
Single-cell suspensions were prepared from the mouse spleen. First, cells were resuspended and blocked with anti-Fc (BioLegend, 422301) for 10–15 min on ice. Then, Zombie Aqua fixable viability dye (BioLegend, 423102) was added to the single-cell suspensions to exclude dead cells. For surface marker detection, the cells were labeled with fluorochrome-conjugated antibodies for 30 min on ice. Surface markers were identified with the following monoclonal antibodies (BioLegend): anti-CD4 (FITC, clone RPA-T4), anti-CD8 (PE-Cy7, clone SK1), anti-CD27 (APC, clone M-T271), anti-CD38 (PE-Cy7, clone HIT2), anti-CD20 (APC-Cy7, clone 2H7), and anti-CD138 (PerCP/Cy5.5, clone MI15). Finally, the cells were washed twice and detected by flow cytometry (BD Biosciences).
Quantitative real-time PCR
Total RNA was extracted from the SG, lung, spleen, liver, and kidney of mice using TRIzol (Accurate Biology), and reverse transcribed into cDNA with the Evo M-MLV RT Mix Kit (Accurate Biology, AG11728). qPCR analysis was performed using SYBR® Green Premix (Accurate Biology, AG11701) according to the manufacturer's instructions. The PCR primers used are listed in Table 3.
The Primers Used in This Study for RT-qPCR
Terminal deoxynucleotidyl transferase dUTP nick end labeling assay
Paraffin sections of mouse SGs were prepared as required, and experiments were performed according to the instructions of the TUNEL Kit (APExBIO, Cat. No. K1133). Finally, the images were taken under a fluorescence microscope (Pannoramic MIDI & Pannoramic 250; 3DHISTECH Ltd., Budapest, Hungary). The images were analyzed using ImageJ software.
Immunofluorescence
ANAs were also detected by indirect immunofluorescence analysis of HEp-2 cells with serum samples (EUROIMMUN, FA1520-1005C). Anti-dsDNA antibodies in mouse serum were also detected by indirect immunofluorescence (EUROIMMUN; CF201105AG). Anti-AQP5 antibody (Proteintech; 120334-1-AP), anti-AQP4 antibody (Proteintech; 16473-1-AP), anti-NKCC1 antibody (Proteintech; 13884-1-AP), anti-MDA5 antibody (Invitrogen; PA5-89344), anti-ZBP1 antibody (AdipoGen; AG-20B-0010), anti-MLKL (Cell Signaling Technology; 37333), anti-RIPK3 (Cell Signaling Technology, 91702) were used for immunological fluorescent staining, images were taken by a fluorescent microscope (Pannoramic MIDI & Pannoramic 250; 3DHISTECH Ltd., Budapest, Hungary). The images were analyzed using ImageJ software.
Western blotting analysis
Proteins from SG whole-tissue lysates were electrophoresed on 10% SDS-polyacrylamide gels and transferred to PVDF membranes. The membranes were blocked for 1 h at room temperature. The blots were then incubated overnight at 4°C with rabbit antibodies against caspase 3 (Cell Signaling Technology; 9662S), GSDMD (Cell Signaling Technology; 39754), BAX (ZENBIO, R23596), and BCL2 (ZENBIO, R23309). The membranes were washed three times for 10 min and then incubated for 1 h at room temperature with HRP-conjugated goat anti-rabbit IgG (BOSTER, BA1055). Finally, the membranes were visualized using ECL reagents (Bio-Rad; #1705060).
Statistical analysis
Flow-cytometric data analysis was performed using FlowJo software (BD). Statistical analyses were performed with SPSS 23.0 (SPSS, Inc.) and GraphPad Prism 7.0 (GraphPad Software, Inc.). Values are represented as the means ± SEM. p < 0.05 was considered statistically significant.
Results
The establishment of humanized NSG murine model of SS
Female NSG mice at 8–10 weeks were adoptively transferred with 10 million PBMCs from HCs, active pSS, and SLE patients on D0. Then the mice were subcutaneously injected with C57/B6j SG protein or PBS on D3, D7, and D31, respectively. The schematic diagram of the whole modeling is shown in Figure 1A. There were obvious foci of lymphocyte infiltration in the SG of pSS and SLE mice (Fig. 1B). Compared with the NSG-HC group, the damage scores of SG were significantly increased in the NSG-pSS group and the NSG-SLE group (Fig. 1E). Besides, there were a large number of inflammatory cell infiltration in the lung of NSG-pSS group and NSG-SLE group, and the lobular tissue was severely damaged (Fig. 1C).
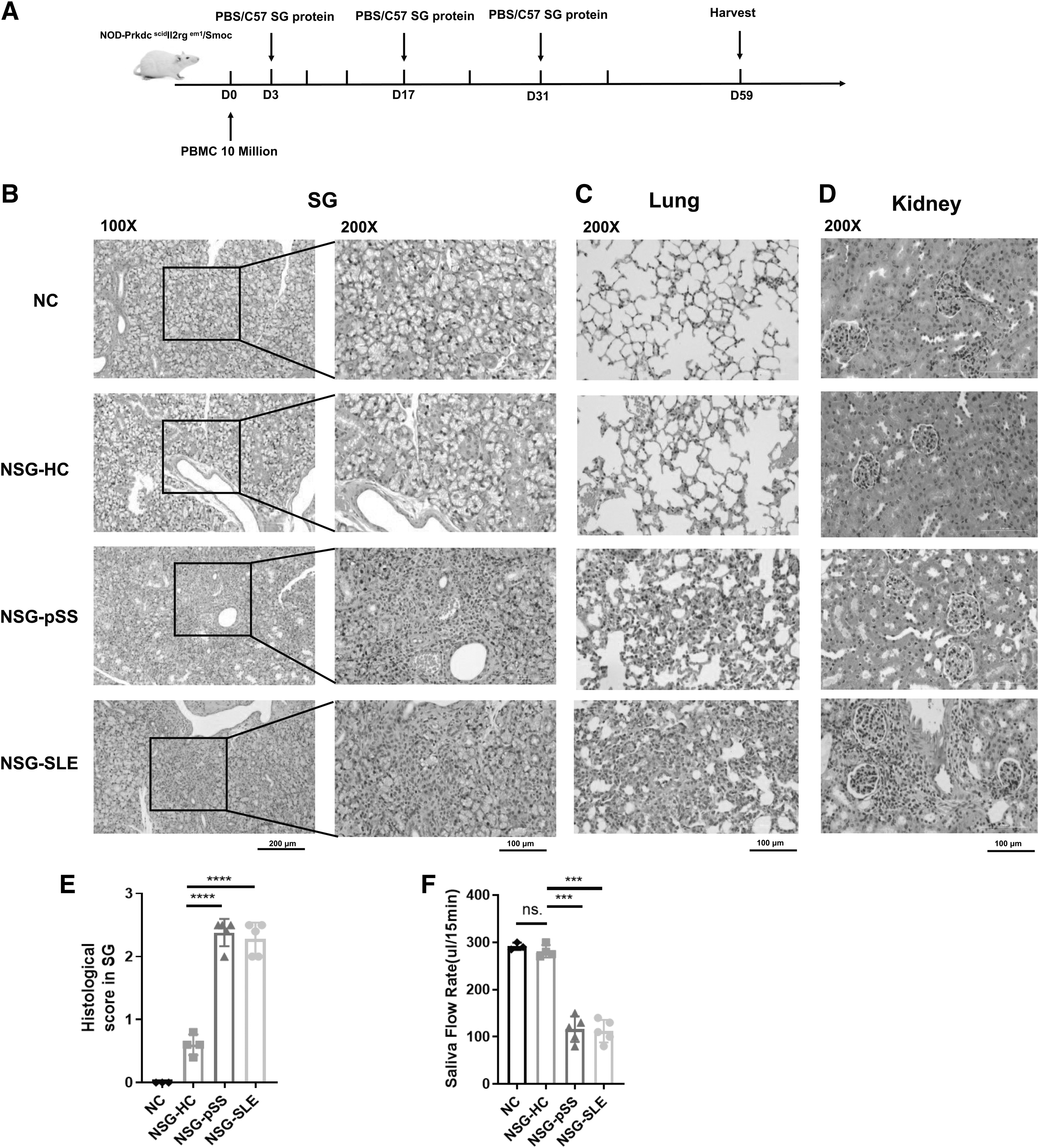
Establishment of a humanized NSG murine model of primary and second Sjogren's syndrome.
Moreover, there was a large number of inflammatory cell infiltration in the renal interstitium of the NSG-SLE group, but this was not observed in the NSG-HC and NSG-pSS groups (Fig. 1D). The salivary flow rate showed that the NSG-pSS group and the NSG-SLE group mice had significantly reduced salivary secretion compared with the NSG-HC group mice (Fig. 1F). While there was no significant change in saliva flow rate in the NSG-HC group and NC group mice. Taken together, there were significant lymphocyte infiltration foci and decreased salivary flow rate in the SGs of pSS and SLE mice. As expected, humanized CD4+T cells, CD8+T cells, CD20+B cells, and plasma cells were detected in the spleen of NSG-HC, NSG-pSS, and NSG-SLE mice by flow cytometry (Supplementary Fig. S1). Overall, we successfully transplanted human PBMCs into NSG mice.
The expression of saliva-related genes was decreased in a humanized NSG murine model of SS
Then, we further investigated the expression of saliva-related genes in the SG of the model mice. Real-time qPCR analysis revealed that the important genes related to salivary secretion, such as water channel (AQP4, AQP5), calcium channel (ITPR3), Na-K-Cl cotransporter (SLC12A2), and muscarinic acetylcholine receptor M3 (CHRM3), were decreased in the SG of NSG-pSS and NSG-SLE mice (Fig. 2A). Immunofluorescence analysis also showed that the expression of AQP4, AQP5, and NKCC1 encoded by SLC12A2 gene in the SG of NSG-pSS and NSG-SLE mice was significantly decreased (Fig. 2B). These results suggest that salivary secretion was impaired in the humanized NSG murine model of SS.
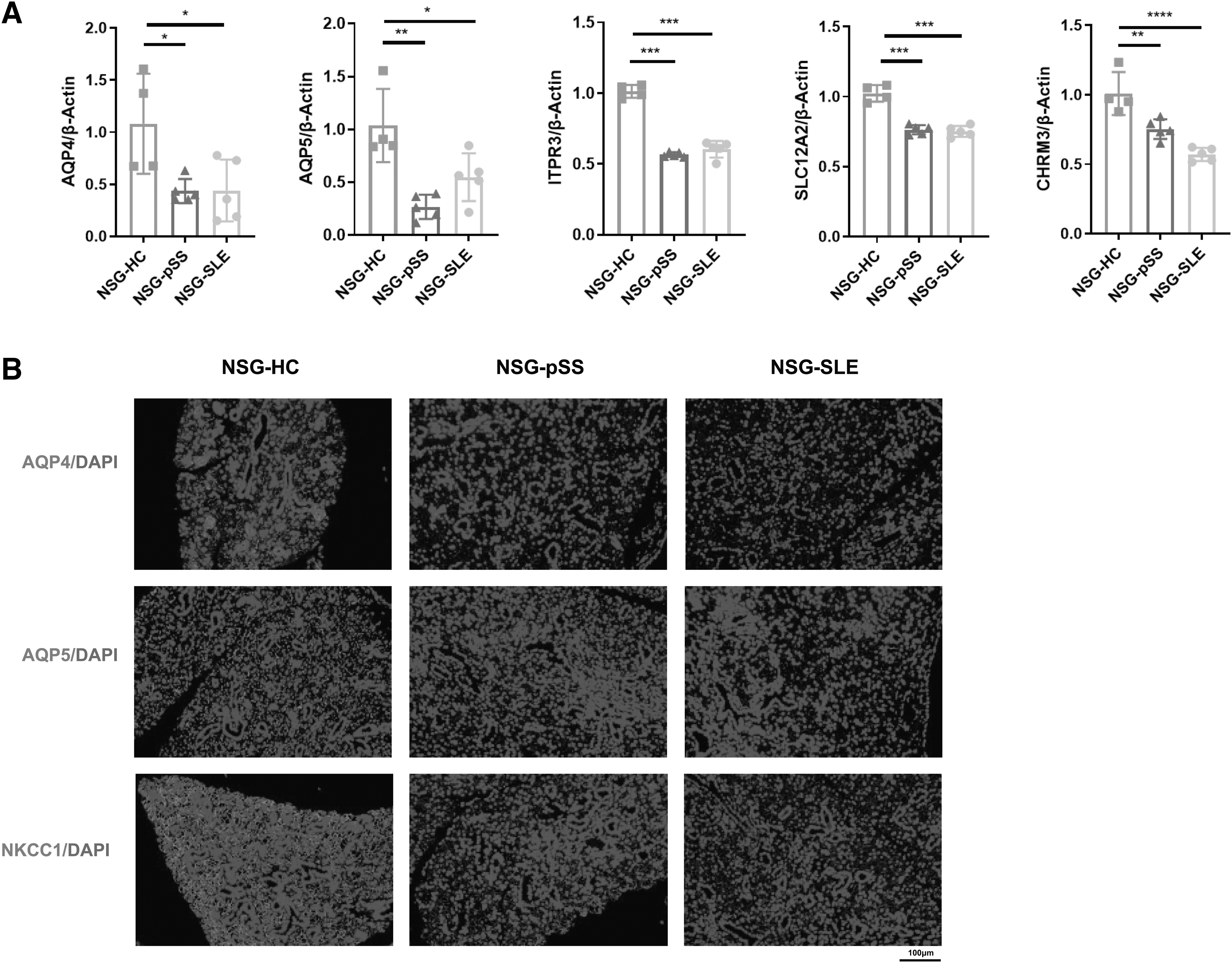
The expression of saliva-related genes was decreased in a humanized NSG murine model of SS.
The production of autoantibody was elevated in a humanized NSG murine model of SS
A large number of autoantibodies are produced in patients with SS. We next explored the production of autoantibodies in this humanized NSG murine model of SS. Indirect immunofluorescence analysis revealed that there were a large number of antinuclear antibodies in the serum of NSG-pSS and NSG-SLE mice (Fig. 3A). Compared with the NSG-HC group, the serum ANA+% in NSG-pSS group and NSG-SLE group was significantly increased (Fig. 3B). Meanwhile, anti-dsDNA antibody was also observed in the serum of NSG-SLE mice, but not in the serums of NSG-HC and NSG-pSS mice (Fig. 3C). This indicated that the humanized NSG murine model of SS is consistent with the clinical patients with SS, and both can detect autoantibodies.

Increased autoantibody production was observed in a humanized NSG murine model of SS.
The expression of IFN genes was increased in the SG of the humanized NSG murine model of SS
IFN signaling pathway is closely related to the pathogenesis of SS (Bodewes and Versnel, 2018). We next examined the expression of IFN-related genes in the humanized NSG murine model of SS. RT-qPCR results indicated that the expression of IFN-related genes (ISG15, IFIT3, RSAD2, IFIH1, ZBP1) was significantly upregulated in the SG of NSG-pSS and NSG-SLE mice compared with NSG-HC mice (Fig. 4A). Meanwhile, immunofluorescence revealed that the expression of MDA5 encoded by IFIH1 was increased mainly in the acinar epithelial cells and infiltrating lymphocytes of SG of NSG-pSS and NSG-SLE mice (Fig. 4B). Interestingly, ZBP1 was mainly upregulated in the duct epithelium of SG of NSG-pSS and NSG-SLE mice (Fig. 4C). Furthermore, IFN-related genes were also increased in liver, lung, spleen, and kidney of the humanized NSG murine model of SS, including ISG15, IFIT3, and RSAD2 (Supplementary Fig. S2).

The expression of IFN genes was upregulated in the SG of the humanized NSG murine model of SS.
Apoptosis was increased in the SG of the humanized NSG murine model of SS
Apoptosis of labial gland epithelial cells also plays an important role in the pathogenesis of SS (Nakamura et al., 2018). We next examined apoptosis in the model mice. TUNEL assay showed that apoptosis was significantly increased in the SG of NSG-pSS and NSG-SLE mice (Fig. 5A). The western blotting results showed that the expression of proapoptotic protein BAX was increased in the SG of NSG-pSS and NSG-SLE mice, and BAX/BCL2 was significantly increased (Fig. 5B, D). The expression of cleaved caspase3 was significantly increased in the SGs of NSG-pSS and NSG-SLE mice, while no significant change was observed about N-GSDMD (Fig. 5C). Taken together, apoptosis is inseparable from SS and plays a crucial role in the humanized NSG murine model of SS.
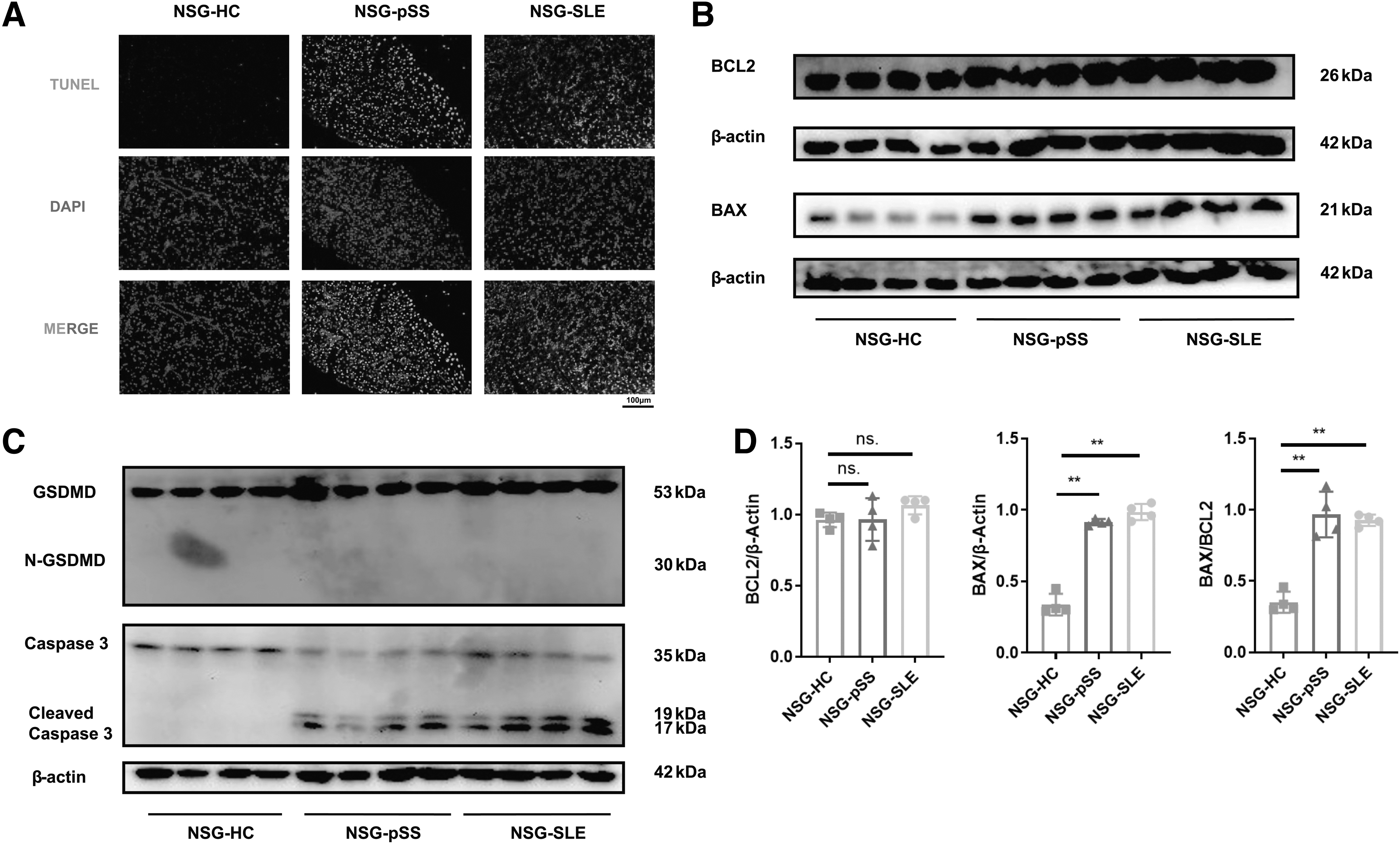
Apoptosis was increased in the SG of the humanized NSG murine model of SS.
Necroptosis was increased in the SG of the humanized NSG murine model of SS
We next examined whether necroptosis existed in the SG of the humanized NSG murine model of SS. Immunofluorescence results revealed that the expression of p-MLKL and p-RIPK3 was increased mainly in the duct epithelium of SG of NSG-pSS and NSG-SLE mice (Fig. 6 A, B), similar to the expression of ZBP1 in SG of the humanized NSG murine model of SS.
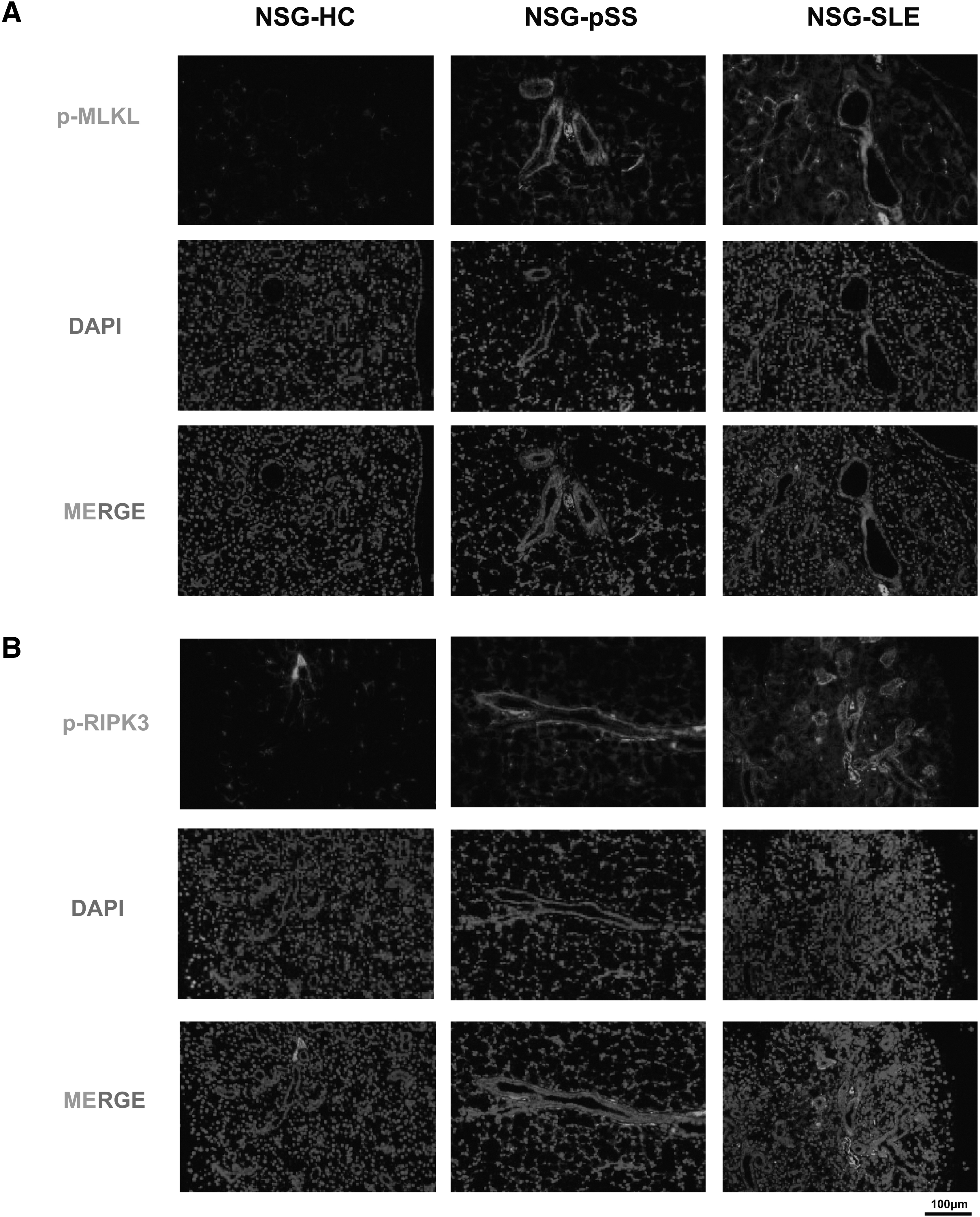
Necroptosis was increased in the SG of the humanized NSG murine model of SS.
Discussion
SS is a complex chronic autoimmune disease (Fox, 2005). Appropriate animal models are the key to understanding and recognizing SS. In this study, we developed a humanized murine model of SS, including primary and secondary SS, with decreased salivary flow rate, massive lymphocyte infiltration in the SG, autoantibody production, lung lesions, and decreased expression of genes related to salivary secretion, which were similar to clinical patients. Moreover, in this mouse model of SS, not only type I IFN-related genes were upregulated but also PANoptosis, including apoptosis and necroptosis, was identified. The successful establishment of this murine model will help us study the pathogenesis of SS better and provide new ideas for the diagnosis and treatment.
A growing number of mouse models of SS have been developed, both spontaneous and induced (Gao et al., 2020; Scuron et al., 2019; Xiao et al., 2019). However, existing mouse models have two major limitations. On the one hand, patients with SS have a wide range of clinical features, including glandular and extraglandular manifestations and the development of B cell malignancies. However, no mouse model has yet been developed that covers all immunological and clinical aspects of human SS. On the other hand, patients with SS can be classified as those with mild disease, such as those with only dry eyes and/or dry mouth, or those with severe systemic disease, such as those with severe interstitial lung disease.
However, most mouse models of SS exhibit only mild disease, and there is a lack of mouse models that exhibit symptoms similar to pSS with systemic disease, with the exception of some mouse models, such as NOD mice, which exhibit multiple autoimmune diseases. There may be several reasons for the differences between the human disease model and the mouse model. Thus, an ideal mouse model to simulate the full profile of human SS is still lacking. Here, the humanized mouse model of SS using NSG mice effectively improved the shortcomings of previous mouse models. This humanized NSG murine model of SS has manifestations of not only glandular involvement in the SG, but also extraglandular involvement, such as lung involvement, which is consistent with clinical patients.
In addition, NSG mice were able to replicate not only the primary SS mouse model but also the secondary SS mouse model, which is not comparable to previous mouse models of SS.
Type I IFNs are an important class of cytokines produced by innate immune cells and also acting on adaptive immune cells (Jiang et al., 2020). Studies have shown that the abnormal expression of type I IFNs and/or type I IFN-inducing gene signatures in serum or labial glands of patients with SS is related to its pathogenesis, clinical manifestations, and disease activity (Brkic and Versnel, 2014; Fernandez-Ruiz and Niewold, 2022; Li et al., 2013; Yao et al., 2013). Therefore, abnormal expression of type I interferon-related genes can be considered as a signature of SS.
PANoptosis (Pandian and Kanneganti, 2022), an inflammatory programmed cell death, is regulated by the PANoptosome (pan-apoptotic body) complex, mainly including apoptosis, necroptosis, and pyroptosis (Bertheloot et al., 2021; Ketelut-Carneiro and Fitzgerald, 2022). The infiltration of a large number of T cells, B cells, and other lymphocytes, the reduced killing effect of NK cells on CD4+T cells, and the impaired phagocytosis of monocytes (reduced phagocytic ability and inability to produce immunosuppressive cytokines) can directly or indirectly induce apoptosis of ductal cells, acinar epithelial cells, and goblet cells of exocrine glands (Nakamura et al., 2018). However, pyroptosis and necroptosis in SS are rarely studied. Type I IFN increased pyroptosis associated with inflammasomes in salivary glands of patients with primary SS (Hong et al., 2020). Also, the necroptotic markers, p-MLKL and RIPK3, were highly expressed in TNF-α-treated salivary gland cells (Hwang et al., 2021).
TUNEL staining, BAX/BCL2 ratio, BAX, and activated caspase3 were significantly increased in the SG of this humanized NSG murine model of SS, indicating that apoptosis was a key factor in the pathogenesis of this model. Increased expression of p-MLKL and p-RIPK3 was also detected in the ductal epithelial cells of the SG of this humanized NSG murine model of SS, which was consistent with the localization of ZBP1. These results suggest that ZBP1 may be involved in the pathogenesis of SS by mediating the phosphorylation of MLKL and RIPK3, leading to necroptosis of ductal epithelial cells. N-GSDMD was not detected in the SG of this humanized NSG murine model of SS, indicating that GSDMD-mediated pyroptosis may not be a major cause of the pathogenesis.
We have successfully established a humanized NSG murine model of SS and demonstrated that type I IFN, apoptosis, and necroptosis are closely related to this model. A limitation of this study is that it does not demonstrate how type I interferon mediates cell death in different cell types in SG of this humanized NSG mouse model. Next, this model will be used to investigate the specific mechanisms of ductal epithelial cell death, hopefully providing new ideas for the diagnosis and treatment of SS.
Conclusion
A typical humanized murine model of SS, including primary and secondary SS, was successfully established. It was characterized by decreased salivary flow rate, downregulated expression of genes related to salivary secretion, SG involvement, extrathyroidal organ involvement, and the production of autoantibodies, which were very similar to those in clinical patients. PANoptosis features, including apoptosis and necroptosis, were present in the SG of this model. Further study will help us better elucidate the pathogenesis of SS.
Ethical Approval Statement
All experiments with human samples were approved by the Ethics Committee of Xiangya Hospital, Central South University (reference number: 2019030465). All animal experiments were approved by the Ethics Committee for Animal Experiments of Central South University (reference number: 2020sydw0950).
Footnotes
Acknowledgment
We thank all patients who participated in the study.
Authors' Contribution
Y.Y. conceived and designed the study, performed the experiments, analyzed the data, and drafted this article as the first author. X.X. performed the experiments and prepared figures and/or tables. M.G. and H.Z. contributed to the analysis of the data and to the revising of the article. All authors have read and approved the final article.
Disclosure Statement
No competing financial interests exist.
Funding Information
This work was supported by the National Natural Science Foundation of China (Grant No. 81901664, 82070018, 82270097), Natural Science Foundation of Hunan Province (2020JJ5894).
Supplementary Material
Supplementary Figure S1
Supplementary Figure S2
References
Supplementary Material
Please find the following supplemental material available below.
For Open Access articles published under a Creative Commons License, all supplemental material carries the same license as the article it is associated with.
For non-Open Access articles published, all supplemental material carries a non-exclusive license, and permission requests for re-use of supplemental material or any part of supplemental material shall be sent directly to the copyright owner as specified in the copyright notice associated with the article.