
Abstract
This review presents a summary of recent progress in research on the N6-methyladenosine (m6A) modification and regulatory roles in hepatic lipid metabolism. As the most abundant internal modification of eukaryotic RNA, the m6A modification is a dynamic and reversible process of the m6A enzyme system, which includes writers, erasers, and readers. m6A methylation depressed lipid synthesis and facilitated lipolysis in liver. The depletion of m6A methyltransferase Mettl14/Mettl3 raised fatty acid synthase (FAS), stearoyl-CoA desaturase-1 (SCD1), acetyl-CoA carboxylase (ACC), and elongase of very long chain fatty acids 6 (ELOVL6) in rodent liver, causing increases in liver weight, triglyceride (TG) production, and content in hepatocytes. FTO catalyzed m6A demethylation and the suppression m6A reader YTHDC2 promoted hepatocellular TG generation and hepatic steatosis in C57BL/6 mice through sterol regulatory element-binding protein 1c (SREBP-1c) signaling pathway, which upregulated the lipogenic genes FAS, SCD1, ACC, recombinant acetyl coenzyme a carboxylase alpha, and cell death-inducing DNA fragmentation factor-like effector C (CIDEC). Furthermore, FTO overexpression did not only enhance mitochondrial fusion to impair mitochondrial function and lipid oxidation but also promoted lipid peroxidation, accompanied by excessive TG in hepatocytes and rodent liver. Elevated m6A modification potently suppressed hepatic lipid accumulation, while the shrinkage of m6A modification arose hepatic lipid deposition. These findings have highlighted the beneficial role of m6A RNA methylation in hepatic lipid metabolism, potentially protecting liver from lipid metabolic disorders.
Introduction
The concept of the N6-methyladenosine (m6A) modification onto an RNA nucleoside emerged in pioneering studies in the 1970s (Brocard et al., 2017). More than 170 different modifications have been reported thus far (Ma et al., 2019; Zhuo et al., 2022); m6A is the most abundant internal modification of eukaryotic mRNA and plays significant roles in pleiotropic physiological and pathophysiological processes, including intracorporeal hepatic lipid metabolism (Dai et al., 2023; Kang et al., 2023; Petri et al., 2023; Yang et al., 2022).
Furthermore, the m6A modification is a dynamic, reversible chemical process that is strictly mediated by m6A modification enzymes, including methylases (writers), demethylases (erasers), and methylation recognition enzymes (readers) together (Bi et al., 2019; Flamand et al., 2023; Jiang et al., 2021; Shi et al., 2019). These m6A enzymes alter the m6A modification of a target mRNA to affect its splicing, stability, transport, or translation, widely regulating the expression of multiple genes, including hepatic lipid-related genes (Breger et al., 2023; Sikorski et al., 2023; You et al., 2022; Zhu et al., 2023).
Recent studies have authenticated the involvement of m6A modification in hepatic lipid metabolism. Clinical detection demonstrated that the total level of hepatic m6A methylation was diminished in patients with lipid disorders in liver (Salisbury et al., 2021; Tang et al., 2023; Zhou et al., 2021). The experimental investigations showed that systemic and hepatic levels of m6A modification were distinctly decreased in the animal models with hepatic lipid dysmetabolism (Heng et al., 2020; Luo et al., 2019).
Liver-specific deletion of m6A writers led to the deficiency of m6A methylation and the accumulation of excessive triglyceride (TG) in rodent liver (Li et al., 2021; Salisbury et al., 2021). Eraser overexpression and the impairment of m6A methylation gave rise to hepatocellular fat deposition and the occurrence of fatty liver in mice (Chen et al., 2015; Sun et al., 2021; Tang et al., 2023). Specific knockout of hepatic m6A readers heightened TG content in hepatocytes and rodent liver (Zhou et al., 2021). All these findings indicate a beneficial role of m6A modification in hepatic lipid metabolism, which potentially protects liver from lipid metabolic disorders.
Along with advances in m6A research and the discovery of important roles of m6A in hepatic lipid metabolism, m6A modification has received increasing attention. In this article, we summarize the definition of the m6A modification and its regulatory enzyme system, and then review the members and functions of m6A enzyme system, including writers, erasers, and readers; next, we summarize the latest findings of m6A modification involvement on the evidence of hepatic lipid metabolism and molecular mechanisms; finally, we explore the effect of m6A modification on hepatic lipid deposition and lipid metabolic disorders, in an attempt to provide updated information on the ongoing study of m6A modification and novel ideas based on m6A modification for the prevention of hepatic lipid disorders.
m6A RNA Methylation
The m6A modification is a methyltransferase-catalyzed reaction at the nitrogen atom in the sixth position of adenosine (A) and is mostly found in the consensus sequence RRACH (R corresponds to G or A; H corresponds to A, C, or U) within the RNA strand (Luo et al., 2018). Since it was discovered in the 1970s, m6A RNA methylation has been confirmed as the most prevalent internal modification across species from yeast, plants, and flies to mammals (Yadav et al., 2018). However, m6A marks are not equally distributed in the transcriptome; they are preferentially enriched near stop codons and in 3′-untranslated regions (3′-UTRs), 5′-UTRs, and long internal exons (Hong, 2018). m6A RNA methylation at different modification sites exerts different effects on RNA metabolism, including RNA stability, translation, splicing, transport, and localization (Zhao and He, 2015).
The processes of m6A RNA methylation are mainly manipulated by the m6A enzyme system. With the development of enzyme technology, recent studies have established that three main enzymes are involved in this modification process: adenosine methyltransferases (writers), demethylating enzymes (erasers), and the m6A-binding proteins (readers) (Fig. 1). Using an active methyl group (e.g., S-adenosylmethionine) as the donor, the adenosine methyltransferases are responsible for the cotranscriptional catalytic addition of m6A to substrate RNAs in a highly specific manner—only a small fraction of the potential m6A consensus sites are methylated. The basis for this transcript specificity and site specificity remains poorly understood.

Schematic summary of the process and biological function of m6A RNA methylation catalyzed by related enzymes. m6A modification is a dynamic and reversible process manipulated by the m6A enzyme system, including “writers,” “erasers,” and “readers.” m6A modification is catalyzed by writers containing catalytic and regulatory subunits. Removal of the methyl group from m6A-modified RNA is mediated by erasers such as ALKBH5 and FTO demethylases. m6A-modified RNA is recognized and decoded by readers that have distinct effects on the metabolism and biological function of substrate RNA strands. METTL3, methyltransferase-like 3; METTL14, methyltransferase-like 14; METTL16, methyltransferase-like 16; WTAP, Wilms' tumor 1-associating protein; RBM15, RNA-binding motif protein 15; VIRMA, Vir-like M6A methyltransferase-associated protein; HAKAI, the E3 ubiquitin-protein ligase Hakai; ZC3H13, zinc finger CCCH domain-containing protein 13; FTO, fat mass and obesity-associated protein; ALKBH5, α-ketoglutarate-dependent dioxygenase AlkB homolog 5; YTHDC1, YTH domain-containing 1; YTHDC2, YTH domain-containing 2; HNRNPC, heterogeneous nuclear ribonucleoprotein C; YTHDF1, YTH N6-methyladenosine RNA binding protein 1; YTHDF2, YTH N6-methyladenosine RNA binding protein 2; YTHDF3, YTH N6-methyladenosine RNA binding protein 3; IGF2BPs, insulin-like growth factor 2 mRNA-binding proteins; eIF3, eukaryotic initiation factor 3.
The m6A demethylases, which are erasers that remove the m6A modification on the RNA molecule by demethylating the sixth nitrogen atom of adenylate, a key step in the reversibility of the m6A modification to shape the m6A epitranscriptome. m6A erasers now appear to have a limited role under normal physiological conditions, but show potential efficacies upon pathological circumstances. m6A requires binding proteins (readers) to selectively recognize and bind m6A-methylated RNA, which decode the signatures of m6A methylation and perform their biological effects on the fates of target RNA (Berlivet et al., 2019; Chen et al., 2019; Fu et al., 2014; Xu et al., 2022).
Briefly, m6A methylation is the most common posttranscriptional RNA modification throughout the transcriptome and universally affects both coding RNA and noncoding RNA. The m6A modification is not an equally distributed mark across the whole transcriptome, but is enriched at multiple specific sites within the RNA strand. The selective m6A methylation occurred when the consensus sequence RRACH are bound with some RBPs, transcription factors, RNA polymerase, and other yet to be discovered factors, or underwent the Histone 3 Lys 27 trimethylation histone modification, recruiting METTL3/METTL14 complexes to the corresponding mRNA sites and catalyzing the selective m6A modification (He and He, 2021).
m6A RNA methylation is a dynamic and reversible enzymatic reaction, and methylation homeostasis is cooperatively maintained by the m6A enzyme system, which includes writers, erasers, and readers. Aberrant expression or interrupted interplay of these main enzymes under some environmental stimuli or intracorporeal pathogenic conditions leads to loss of m6A modification homeostasis in liver and potentially causes the development of hepatic diseases related to lipid dysmetabolism.
m6A Enzyme System
m6A writers are assembled into methyltransferase complexes, containing catalytic and regulatory subunits. The catalytic subunits currently known include methyltransferase-like 3 (METTL3), methyltransferase-like protein 16, and methyltransferase-like 14 (METTL14), which comprise the core catalytic center of methyltransferase complexes. The known regulatory subunits contain Wilms' tumor 1-associating protein, RNA binding motif protein 15, Vir like m6A methyltransferase-associated protein, E3 ubiquitin-protein ligase Hakai, and zinc finger CCCH domain-containing protein 13, which assist the anchoring of m6A methyltransferase complexes and the positioning of substrate mRNA for m6A modification (Pu et al., 2023; Zhu et al., 2023). Specifically, these regulatory subunits cooperatively found the RRACH consensus sequence within substrate RNA strand and recruited catalytic component to these sites for m6A methylation (Patil et al., 2016; Tong et al., 2018; You et al., 2022).
So far, fat mass and obesity-associated protein (FTO) and ALKBH5 are the main m6A erasers (Pinello et al., 2018; Sun et al., 2019). These two m6A erasers harbor the well-conserved AlkB-like catalytic domain within their respective polypeptide chains. Moreover, FTO and ALKBH5 recognize RNA targets and catalyze m6A demethylation in different specific ways (Pinello et al., 2018). FTO was discovered to efficiently demethylate m6A to adenosine by oxidizing m6A to intermediates—N6-hydroxymethyladenosine and N6-formyladenosine—in a stepwise manner (Rajecka et al., 2019; Yang et al., 2019).
FTO was reported to catalyze the demethylation of m6A residues both in vitro and in vivo, as well as m6A. Upon further investigation, it was determined that FTO not only mediates the demethylation of m6A within the nucleus but also catalyzes the demethylation of cap m6A in the cytoplasm. In contrast to FTO, ALKBH5 is the only specific m6A demethylase in vivo, and prefers single-stranded nucleic acid substrates. ALKBH5 directly reverses the m6A modification to reestablish adenosine with no detected intermediate (Aik et al., 2014; Niu et al., 2013; Rajecka et al., 2019).
At least 10 readers have been identified thus far, and they are all conserved YTH domain-containing proteins or non-YTH RNA-binding proteins. The YTH domain-containing proteins discovered to date include three cytoplasmic proteins: The YTH domain family, YTH domain-containing 1, and YTH domain-containing 2 (YTHDC2) (Chen et al., 2023; Chen et al., 2019; Hong, 2018). The three cytoplasmic YTHDF proteins work in a synergistic manner to influence the foundational biological processes related to m6A modification, such as translation initiation and RNA decay (Meyer, 2019; Sikorski et al., 2023). The two nuclear/nucleocytoplasmic YTH domain-containing proteins regulate the splicing process and subsequent fate of m6A-modified mRNA strand. The non-YTH RNA-binding proteins currently include HNRNPA2B1, insulin-like growth factor 2 mRNA-binding proteins, and eukaryotic initiation factor 3, which are also recruited by m6A-modified RNAs to influence the translation and expression subsequently (Coker et al., 2019; Sun et al., 2024).
m6A Methylation Modulates Hepatic Lipid Metabolism
m6A modification ubiquitously participates in hepatic lipid metabolism (Fig. 2). On the one hand, m6A modification depresses lipogenic genes to reduce hepatic lipid synthesis by impacting mRNA stability and decay, and subsequent translation. Liver-specific Mettl14 depletion augmented mRNA lifetimes and protein expression of diacylglycerol acyltransferase 2, fatty acid synthase (FAS), stearoyl-CoA desaturase-1 (SCD1), acetyl-CoA carboxylase (ACC), and elongase of very long chain fatty acids 6 in C57BL/6 mice administered with AAV8.tbg.cre, resulting in an increase in the production of hepatocellular TG (Salisbury et al., 2021).
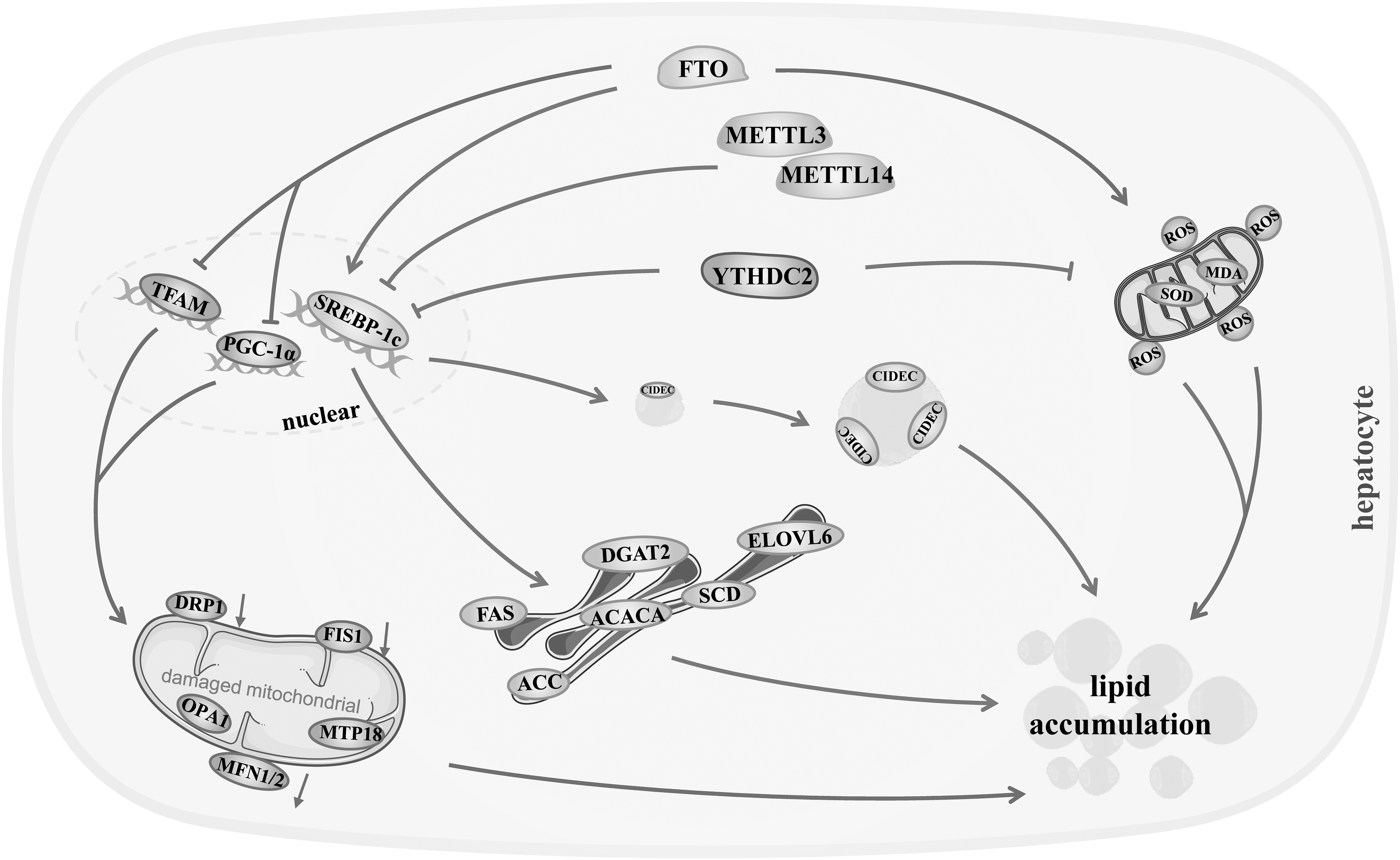
Schematic summary of the molecular mechanisms of m6A modification regulation in hepatic lipid metabolism. m6A modification depresses lipogenic genes to reduce hepatocellular lipid synthesis, and promotes lipolysis through mitochondrial oxidation, thereby suppressing hepatocellular lipid accumulation and the development of hepatic lipid disorders. FAS, fatty acid synthase; SCD, stearoyl-CoA desaturase; ACC, acetyl-CoA carboxylase; ELOVL6, very long chain fatty acids 6; ACACα, recombinant acetyl coenzyme a carboxylase alpha; SREBP-1c, sterol regulatory element-binding protein 1c; PGC-1α, peroxisome proliferator-activated receptor-γ coactivator-1α; MFN1/2, mitofusins; FIS1, fission 1; OPA1, optic atrophy 1; DRP1, dynamin-related protein 1; TFAM, mitochondrial transcription factor A; SOD, superoxide dismutase; MDA, malondialdehyde; YTHDC2, YTH domain-containing 2; CIDEC, cell death-inducing DNA fragmentation factor-like effector C; DGAT2, diacylglycerol acyltransferase 2.
FTO-expressing adenovirus upregulated the mRNA and protein level of ATP-citrate lyase, ACC1, FASN, and SCD1, enhancing hepatic lipogenesis and TG synthesis in chow diet-fed C57BL/6 mice (Tang et al., 2023). YTHDC2 siRNA ascended the mRNA lifetime of SREBP-1c-heightened FASN, SCD1, ACC1, and TG yield in Hep1-6 cell (Zhou et al., 2021). Specific knockout of hepatic YTHDC2 elevated adipogenic genes, including SREBP-1c, FASN, SCD1, ACC1, and at mRNA and protein level, leading to TG production in the liver of lean C57BL/6 mice (Zhou et al., 2021).
FTO knockdown restrained sterol regulatory element-binding protein 1 (SREBP-1), a key transcription factor regulating the de novo synthesis of fatty acids in liver, which raised the lipogenic genes FANS, recombinant acetyl coenzyme A carboxylase alpha, and SCD at both mRNA and protein levels, as well as m6A modification on the mRNA strand of these genes in Hepa1-6 cells (Hu et al., 2020). Besides, the overexpression of FTO accelerated the maturation and nuclear translocation of SREBP-1c in a demethylation-dependent manner, promoting the expression of cell death-inducing DNA fragmentation factor-like effector C (CIDEC) and the formation of lipid droplet in HepG2 cells (Chen et al., 2018).
On the other hand, m6A methylation facilitates the lipolysis through lipid oxidation in hepatocytes (Fig. 2). FTO not only upregulated some genes associated with mitochondrial fusion, including mitofusins and optic atrophy 1, but also downregulated mitochondrial fission genes, including fission 1, dynamin-related protein 1, mitochondrial protein 18, and mitochondrial biogenesis genes such as peroxisome proliferator-activated receptor-γ coactivator-1α and mitochondrial transcription factor A, which ultimately led to a decrease in mitochondrial number and ATPase activity, and an impairment in β-oxidation and lipolysis in mitochondrion, causing an increase in hepatic TG content in HepG2 cells (Kang et al., 2018). FTO overexpression brought a significant decrease in superoxide dismutase activity, which induced hepatocellular lipid peroxidation and elevated the level of lipid peroxidation marker malondialdehyde, and accompanied by excessive TG and total cholesterol in oleic acid-induced L02 cells (Sun et al., 2021).
m6A Modification Inhibits Hepatic Lipid Accumulation
Clinical observations show that m6A methylation or its enzymes are altered in patients with hepatic disorders of lipid metabolism (Table 1). The methyltransferase METTL3 was significantly reduced by 71.2% at the protein level in nuclei of hepatocytes from nonalcoholic steatohepatitis (NASH) patients (Li et al., 2021). Hepatic METTL14 expression was markedly descended, which diminished m6A modification in individuals with fatty liver disease (Salisbury et al., 2021). The demethylase FTO was significantly elevated, and concomitantly accumulated large amounts of fat in the liver of NASH patients (Luo et al., 2019). Significant increases in FTO mRNA and protein were observed in the livers of patients with nonalcoholic fatty liver disease (NAFLD) (Li et al., 2020; Luo et al., 2019). YTHDC2 expression, an m6A reader to recognize and decode the m6A-modified mRNA, was lowered in the livers of population with NAFLD or NASH (Zhou et al., 2021).
The Effect of m6A Modification on Hepatic Lipid Accumulation
NASH, nonalcoholic steatohepatitis; METTL3, methyltransferase-like 3; METTL14, methyltransferase-like 14; NAFLD, nonalcoholic fatty liver disease; FTO, fat mass and obesity-associated protein; YTHDC2, YTH domain-containing 2; ↑, increased; ↓, decreased; —, nonobserved result.
Experimental investigations demonstrate that elevated m6A modification potently suppresses hepatic lipid deposition (Table 1). Betaine-raised m6A methylation obviously decreased the intracellular TG content and relieved hepatic lipid accumulation in HepG2 cells (Kang et al., 2018). Another study also showed that berberine-enhanced m6A modification activated extracellular regulated protein kinases, causing a mitigation in lipid accumulation of zebrafish hepatocytes (Zhang et al., 2022). Dietary supplement of curcumin heightened the abundance of m6A methylation and depressed the expression of lipogenic genes, which alleviated lipopolysaccharide-induced disruption of lipid metabolism in piglet livers (Lu et al., 2018).
Entacapone feeding augmented hepatic m6A modification, and plainly reduced the expression of lipogenic genes in the livers, ultimately suppressing the hepatic TG content and ameliorating high-fat diet-induced steatosis in C57BL/6 mice (Tang et al., 2023). The arsenite (3) methyltransferase depletion facilitated m6A modification and m6A-mediated miR-142-5p maturation in the livers of arsenic-exposed As3MT+/− mice, which reduced the expression of lipogenic genes and opposed NAFLD induced by chronic arsenic exposure (Li et al., 2023). FTO shRNA brought about the reductions in hepatic TG content, lipid deposition, and liver weight in dexamethasone-induced fatty liver of C57BL/6 mice (Hu et al., 2020). YTHDC2 overexpression diminished hepatic TG accumulation, improving steatosis in obese C57BL/6 mice (Zhou et al., 2021).
Conversely, the shrinkage of m6A modification raises hepatic lipid deposition. Chronic arsenic exposure lowered hepatic level of m6A methylation, resulting in lipid accumulation and steatosis in the liver of C57BL/6 mice (Li et al., 2023). Systemic and hepatic levels of m6A modification were distinctly reduced upon a high branched-chain amino acid diet in healthy crossbred weaned piglets, which promoted lipid accumulation in liver (Heng et al., 2020). The global levels of m6A RNA methylation were appreciably decreased in the triclosan- or bisphenol A-administrated groups, causing the blister-like fatty degeneration and numerous lipid droplets in the liver tissue of wild-type AB line zebrafish larvae (Sun et al., 2020).
Subcutaneous injection of corticosterone pronouncedly diminished hepatic level of m6A methylation, leading to an increase in liver weight along with a lift in plasma and hepatic TG concentrations in Rugao yellow-feathered chicken (Hu et al., 2020). Hepatocyte-specific Mettl3 knockout mice possessed the increased liver weight, TG content, and lipid droplets. Liver-specific Mettl14 deletion was shown to evidently elevate hepatic TG content in C57BL/6 mice (Salisbury et al., 2021). FTO expression was upregulated at mRNA and protein level in NAFLD rat model (Sun et al., 2021). FTO overexpression was demonstrated to accelerate the development of hepatic steatosis in the liver of chow diet-fed C57BL/6 mice (Tang et al., 2023).
Both FTO overexpression and hepatic deficiency of m6A methylation were able to expand liver weight dramatically in C57BL/6 mice fed with a high-fat diet (Chen et al., 2015). In addition, FTO overexpression heightened the cellular contents of TG and total cholesterol in oleic acid-treated L02 cells (Sun et al., 2021). High expression of FTO raised intracellular TG contents, and contributed to fat deposition in HepG2 cells incubated with free fatty acids (Chen et al., 2018; Kang et al., 2018).
Concluding Remarks
m6A methylation is the most common chemical modification of eukaryotic Mrna (Huang et al., 2020), in which methyl groups are catalyzed and transferred onto the nitrogen atom in the 6th position of adenosine in the RNA strand by writer methyltransferases (Wang et al., 2020). In contrast, this m6A modification is reversed by eraser demethylases (Geng et al., 2020). Readers, RBPs with conserved YTH domains and non-YTH RBPs, decode the m6A signature and mediate the splicing, translation, and stability of the target RNA strand (Wang et al., 2020).
Dynamic and reversible RNA methylation affect the structures and expression of target RNA strands of lipid-related genes, regulating multiple aspects of hepatic lipid metabolism in vivo. m6A methylation has been preliminarily confirmed to play important roles in numerous lipid-related genes in liver, which suppresses hepatic lipid synthesis and facilitates the lipolysis, ultimately resisting excessive lipid accumulation and hepatic lipid dysmetabolism. Furthermore, m6A methylation possesses the potential capability to confront with hepatic lipid disorder diseases such as NAFLD.
However, research on m6A modification and its role in hepatic lipid metabolism is merely the tip of the iceberg, and many problems still need to be resolved. To date, there is no efficient detection method to timely and directly determine the presence of m6A marks or their action sites in the target RNA strand of lipid-related genes. As RNA methylation is a newly discovered modification, currently unknown novel writers, erasers, and readers await to be identified in the future. Which writers, erasers, and readers mainly exist or function in the liver, and these m6A enzymes probably work together to fine-tune m6A modification on the target mRNA strand of hepatic lipid-related genes.
How do these enzymatic proteins work cooperatively, or what upstream mechanism coordinates their actions. Which m6A enzymes are specially involved in other processes of hepatic lipid metabolism such as lipid decomposition, storage, and even in bile acid metabolism closely associated with hepatic lipid metabolism? Upon induced hepatic disorder of lipid metabolism, how does m6A modification change lipid-related genes. Moreover, the causal relationship between m6A modification and hepatic lipid disorder has not yet been clarified (Tang et al., 2023). The facilitation of m6A modification seemingly resists hepatic lipid accumulation and the progression of NAFLD, but its side effect and systemic safety required comprehensive evaluation in vivo. It is also urgently needed to develop a liver-specific approach to manipulate m6A modification in liver.
The regulatory role of m6A modification is underscored in hepatic lipid metabolism, as indicated by gradually advancing research. Recent studies have confirmed that m6A modification manipulates lipid synthesis and lipolysis in liver, and protects hepatocyte from lipid deposition and lipid dysmetabolism, indicating the therapeutic potential of m6A modification in hepatic lipid disorder diseases (Tang et al., 2023). Large-scale investigations are required to clarify which lipid-related genes are primarily affected by these m6A enzymes under hepatic lipid dysmetabolism.
The urgent need is the creation of a consummate m6A epigenetic map of hepatic lipid disorder diseases through big data analysis and biological methodologies. The identification of the main lipid-related genes affected by m6A methylation will provide novel molecular diagnoses and therapeutic targets for medical workers to fight against hepatic diseases caused by lipid disorders in the future. Furthermore, some natural or artificial bioactive components are eagerly developed to intervene m6A modification or m6A enzyme system in liver, to combat with hepatic diseases caused by lipid dysmetabolism in coming years.
Authors' Contribution
X.M.: Writing original draft; S.C. and H.L.: Collecting literature and providing writing strategy; Y.W. and L.Z.: revising the figures and table; Y.L.: Conceptualizing, reviewing, and editing. All authors read the final article and agreed to its publication.
Footnotes
Disclosure Statement
The authors declare no conflict of actual and potential interest.
Funding Information
The author acknowledges the support of the National Natural Sciences Foundation of China (82160098), the Natural Science Foundation of Guangxi Zhuang Autonomous Region (2019JJA140728), and the autonomous project of Guangxi Key Laboratory of Diabetic Systems Medicine (2023ZZ-03).