
Abstract
This study aimed to determine the function of angiopoietin-related protein 4 (ANGPTL4) and bone morphogenetic protein 7 (BMP7) on hepatocellular carcinoma (HCC). Overexpressing plasmids were cotransfected into HepG2 cells to determine the interaction between ANGPTL4 and BMP7. The effect of ANGPTL4 on the stability of BMP7 is examined by detecting the expression and ubiquitination levels. In vitro and in vivo experiments of knocking down ANGPTL4 while overexpressing BMP7 were performed to investigate whether the effects of ANGPTL4 on HCC proliferation, migration, and downstream signaling pathways were dependent on BMP7. ANGPTL4 is able to interact with BMP7, and knockdown of ANGPTL4 increased BMP7 expression and ubiquitination. Overexpression of BMP7 reversed the inhibition of HCC proliferation and migration as well as the decrease in the expression levels of Smad1/5/8 and MAPK14 caused by knockdown of ANGPTL4. ANGPTL4 promotes the proliferation and migration of HCC by inhibiting the ubiquitination degradation of BMP7 and the Smad/MAPK pathway, providing a novel mechanism and a potential therapeutic target for the treatment of HCC.
Introduction
Hepatocellular carcinoma (HCC) is one of the common cancer types, ranking fifth in the overall cancer incidence and second in malignancy mortality (Li et al., 2017; Schweitzer et al., 2015). High aggressiveness and recurrence rates are the main reasons for the low overall survival of HCC (Fang et al., 2022; Zheng et al., 2021). Therefore, further studies are still needed to clarify the pathogenesis of HCC.
Angiopoietin-related protein 4 (ANGPTL4) was previously recognized as a multifunctional protein involved in energy metabolism and vascular homeostasis. However, recent evidence has shown that its coding gene ANGPTL4 plays a crucial role in human tumorigenesis (Fang et al., 2022; Xiao et al., 2022; Zheng et al., 2021). Our previous study revealed its upregulated expression in human HCC tissues and functions of promoting tumor cell proliferation, invasion, and migration. Alternative splicing (AS) is an important form of posttranscriptional modification, which allows the mRNA precursor of a gene to be spliced in different ways to produce various mature mRNA splice isoforms, further resulting in different protein isoforms (Braunschweig et al., 2013; Irimia and Blencowe, 2012; Nilsen and Graveley, 2010). Transcripts produced by AS can exhibit different or even opposite functions (Bechara et al., 2013; Coelho and Smith, 2014; Ladomery, 2013; Lu et al., 2015; Salton et al., 2015; Wang et al., 2014; Zhang and Manley, 2013; Zhou et al., 2014). ANGPTL4 is located on chromosome 19p13.3 and has 7 exons and 6 introns. Through AS, two protein-coding transcripts are currently widely recognized, namely NM_139314.3 (ANGPTL4-transcript-1) and NM_001039667.3 (ANGPTL4-transcript-3). Our previous research also demonstrated that transcript-3 promoted HCC proliferation and inhibited apoptosis more than transcript-1. Moreover, ANGPTL4 upregulates the expression of bone morphogenetic protein 7 (BMP7) to promote metastasis and inhibit apoptosis in colorectal cancer cells. Bone morphogenetic proteins (BMPs) inhibit and stimulate tumor growth, and BMP-7 is a signaling molecule belonging to the transforming growth factor-β superfamily. BMP-7 is expressed in a variety of human cancers and plays an important role in cancer progression. BMP-7 promotes migration and invasion of breast cancer cells (Naber et al., 2012), migration of prostate cancer cells (Dorai et al., 2014), and metastasis associated with colorectal cancer (Li et al., 2015). Related to metastasis, BMP-7 controls tumor progression in a variety of ways. BMP7 regulates breast epithelial cell proliferation and apoptosis by acting as an LIM domain-only 4 (LMO4) response gene. In gastric cancer, the BMP7 promoter has been shown to be methylated, suggesting a role in carcinogenesis (Aoki et al., 2011). In prostate cancer, BMP7 exposure can regulate the biological behavior in a cell-type-specific manner. BMP7 has an important role in the control of lung cancer cell motility and invasiveness (Liu et al., 2012). However, the roles of ANGPTL4 and BMP7 in HCC are unclear. The MAPK pathway plays an important role in tumors. m6A methyltransferase-associated protein KIAA1429 mediates the mechanism of gefitinib resistance in lung adenocarcinoma cells through the activation of the c-Jun N-terminal kinase (JNK)/MAPK signaling pathway (Lin et al., 2023). The piR-YBX1/Y box binding protein (YBX1)/MAPK axis inhibits triple-negative breast cancer proliferation and metastasis (Wu et al., 2024). In this study, we elucidated the interaction between ANGPTL4 and BMP7 and BMP7-mediated ANGPTL4 to promote the proliferation and migration of HCC. ANGPTL4 can effectively regulate the downstreaming of BMP7, which provides a more in-depth mechanism for the role of ANGPTL4 in HCC.
Materials and Methods
Cell culture
The human HCC cell line HepG2 (RRID: CVCL_0027) and the nontumor cell line HEK293 (RRID: CVCL_0045) were obtained from Beina Bio (China). The cells were maintained in Dulbecco’s modified Eagle’s medium (DMEM, Gibco, China) containing 10% (vol/vol) fetal bovine serum (FBS) and 1% (vol/vol) penicillin/streptomycin at 5% (vol/vol) carbon dioxide (CO2) and 37°C incubator. The mycoplasma contamination was excluded by using the MycoFluor™ Mycoplasma Detection Kit (ThermoFisher, USA).
Establishment of knockdown and overexpression of cell lines
To generate ANGPTL4 and BMP7 knockout HepG2 cells, cells were transfected with Cas9/gRNA vectors carrying ANGPTL4 or BMP7 gRNAs using Lipofectamine 3000 (Invitrogen, USA) and subjected to puromycin selection. For ANGPTL4 knockout and BMP7 overexpressing HepG2 cell establishment, the selected ANGPTL4 knockdown cells were transfected with pcDNA3.1-BMP7 by Lipofectamine 3000 (Invitrogen).
Co-IP assay
Plasmids expressing individual gene were transfected into HepG2 cells. At 48-h post-transfection, the transfected cells were lysed using the Co-IP lysis buffer (Proteintech, China). After centrifugation, the supernatants were incubated with the corresponding antibodies at 4°C overnight. Next day, the mixture was incubated with protein A/G agarose beads (Proteintech) for 2 h at room temperature. Precipitated proteins were separated by heating with SDS loading buffer. Subsequently, samples were subject to Western blotting (WB).
WB
Protein in cells was extracted by the RIPA lysis buffer and subject to SDS-polyacrylamide gel electrophoresis (SDS-PAGE). The proteins were transferred onto the polyvinylidene difluoride membrane. After blocking with 5% nonfat milk in Tris Buffered Saline + Tween 20 (TBST), the membrane was incubated with following primary antibodies: ANGPTL4 (Boster A01147, 1:1000), FLAG (Affinity T0003, 1:5000), HA (Affinity T0008, 1:3000), and GAPDH (Affinity T0004, 1:1000), and the secondary antibodies (Affinity S0001, 1:5000 and Affinity S0002, 1:5000). After washing with TBST, membrane was exposed to the Horseradish peroxidase (HRP)-conjugated secondary antibody (Proteintech, SA00001-1, 1:8000) for 1h. The bands were developed by using the Enhanced Chemiluminescence Sensitive Enhanced Chemiluminescence kit (Proteintech).
CCK-8 assay
Cell viability was measured using the Cell Counting Kit 8 (CCK-8) assay kit (DOJINDO, Japan). In brief, cells were seeded in 96-well plates at a density of 1.0 × 104 cells/well and cultured in DMEM containing 10% FBS and 1% penicillin/streptomycin in a 5% CO2 and 37°C incubator. After a 72 h incubation, 10μL of CCK-8 reagent were added to each well. After incubation for 2 h, the absorbance at 450 nm was measured using a microplate reader (BioRad, USA).
Cell migration assay
HepG2 cells were plated into a 12-well plate at 5 × 105 cells per well. A straight line was scratched by a pipette tip. Followed by rinsing with PBS and changing the culture medium to serum-free for an additional 48 h, three randomly selected fields were photographed at different time points. The scratch width was measured by ImageJ software.
Mice xenograft
The protocol of this study was approved by the Ethics Review Committee of the Second Hospital of Harbin Medical University (Committee of the Second Hospital of Harbin Medical University KY-2022-208). Transfected HepG2 cells were made into the cell suspension at 1 × 107 cells/mL density by cell counting. Twenty-four 6-week-old male BALB/c nude mice (Liaoning Changsheng Biotechnology Co., Ltd.) were injected subcutaneously (under the armpit) with 0.1 mL of the cell suspension. Mice were divided into four groups, with six mice in each group. The longest diameter (a) and shortest diameter (b) of the tumor body were measured and recorded twice or three times weekly. The tumor volume (V) was determined using a Vernier caliper by measuring length (L) and width (W) and calculated as (LxW2)/2. Mice under isoflurane anesthesia were sacrificed by cervical dislocation and the tumor body was dissected.
Statistical analysis
In this study, all results are represented as the mean ± standard deviation. One-way analysis of variance (ANOVA) and two-way ANOVA were used to analyze the significance between groups. A p-value <0.05 was set as the significance level.
Results
ANGPTL4 interacts with BMP7
To verify whether ANGPTL4 and BMP7 could interact with each other, the flag-tagged ANGPTL4 plasmid and MYC-tagged BMP7 plasmid were cotransfected into HepG2 cells. Immunoprecipitation experiments were performed 48 h after transfection, and as shown in Figure 1, the BMP7 protein was able to pull down ANGPTTL4, whereas no bands were detected in the control group, suggesting that ANGPTL4 was able to bind to BMP7.
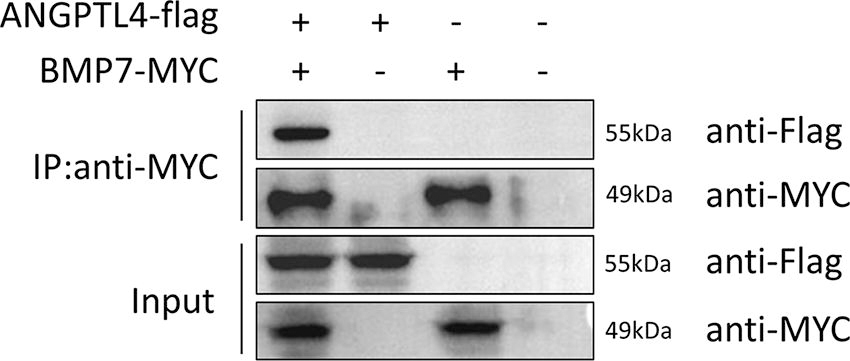
Identification of interaction between ANGPTL4 and BMP7 by Co-IP assay. ANGPTL4, angiopoietin-related protein 4; BMP7, bone morphogenetic protein 7.
ANGPTL4 inhibits BMP7 ubiquitination degradation
To investigate the effect of ANGPTL4 on BMP7, the ANGPTL4 was knocked out in HepG2 cells. BMP7 expression was decreased dramatically compared with wild-type cells. The BMP7 expression was enhanced to normal level when cells were treated with MG132 (Fig. 2A), indicating that BMP7 underwent ubiquitination modification. To further explore the effect of ANGPTL4 on BMP7 ubiquitination, cells were cotransfected with ANGPTL4 and HA-Ub, and then treated with MG132. As shown in Figure 2B, ANGPTL4 protected BMP7 degradation caused by ubiquitination. In addition, knockout of ANGPTL4 significantly impaired BMP7 stability (Supplementary Fig. S1).
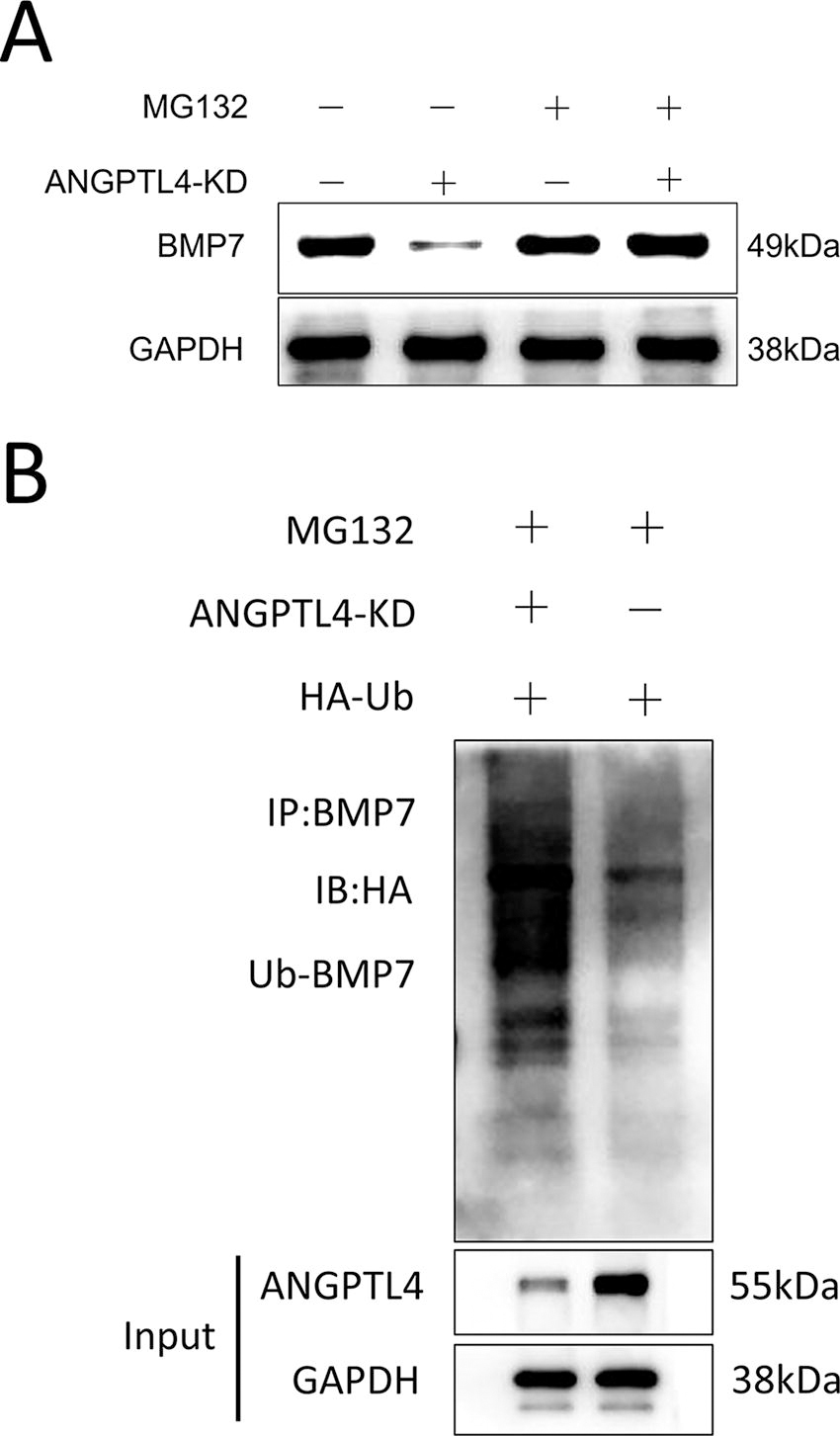
ANGPTL4 inhibits BMP7 ubiquitination degradation.
Effect of ANGPTL4 and BMP7 on the proliferation and migration of HCC
To explore the function of ANGPTL4 and BMP7 on the proliferation of HCC, the cell growth of knockout cells was determined. The cell viability of ANGPTL4-KD and BMP7-KD cells was dramatically decreased compared with the control group (Fig. 3A). However, overexpressing BMP7 in ANGPTL4 knockout cells increased cell viability. To further validate this phenomenon, the wound healing assay was performed to determine cell migration ability. In agreement with CCK8 results, knockdown of ANGPTL4 or BMP7 dramatically decreased the cell migration of HCC, whereas overexpression of BMP7 reversed the diminished cell proliferation caused by ANGPTL4 knockdown (Fig. 3B).
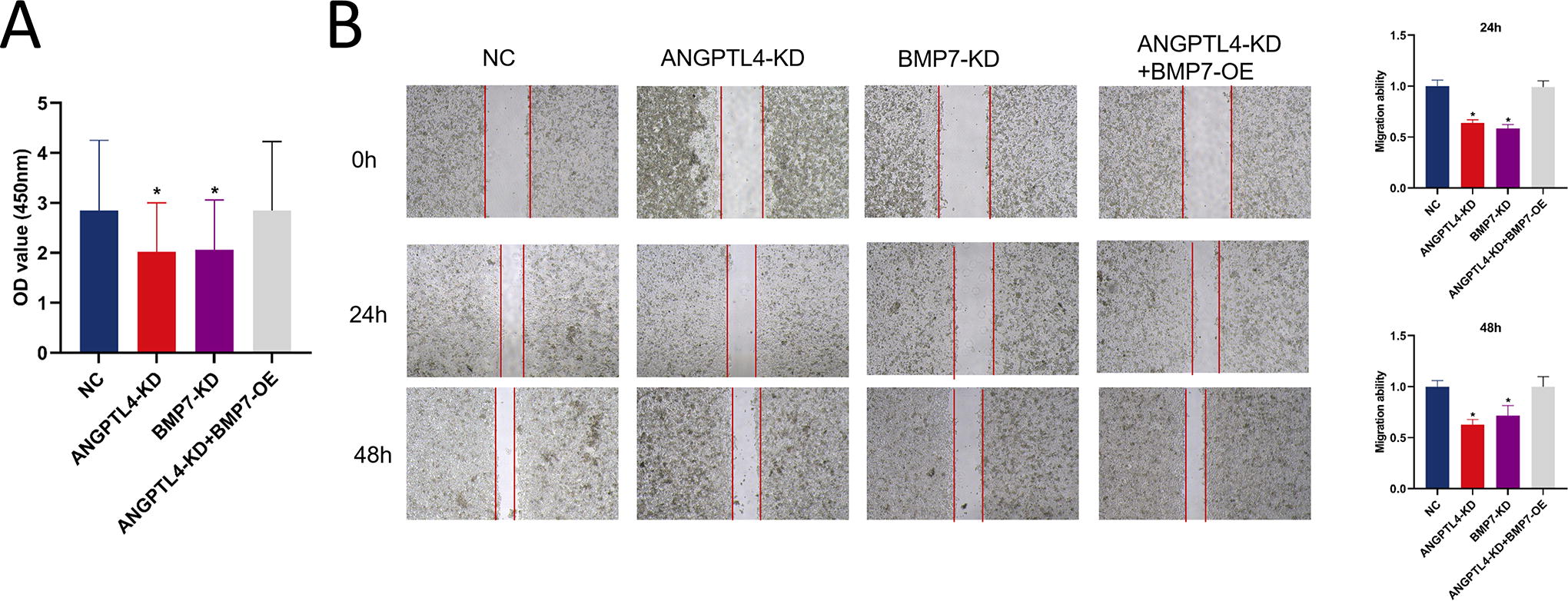
Effect of ANGPTL4 and BMP7 on the proliferation and migration of HCC.
The effect of ANGPTL4 and BMP7 on Smad/MAPK signaling pathways
To explore the mechanism of BMP7 regulation of HCC, we explored the regulation of Smad and MAPK pathways by ANGPTL4 and BMP7. As shown in Figure 4, knockout of ANGPTL4 and BMP7 did not alter the expressions of extracellular signal-regulated kinase (ERK), P-ERK, JNK, and P-JNK. Although no differences of Smad1/5/8 and MAPK14 between the different groups were observed, the P-Smad1/5/8 and P-MAPK14 levels were significantly decreased in the ANGPTL4-KD and BMP7-KD groups. Overexpression of BMP7 on ANGPTL4 knockout cell lines reversed the downregulation of Smad1/5/8 and MAPK14 phosphorylation levels. We further determined the effect of BMP7 on proliferation of HCC by regulation of phosphorylation levels of Smad1/5/8 and MAPK14. The BMP7-overexpressing cells were treated with LY 3200882 (Smad inhibitor) and adjudin (MAPK inhibitor), WB results indicated that inhibitors of Smad and MAPK significantly decreased phosphorylation levels of Smad1/5/8 and MAPK14, and the CCK-8 results showed a corresponding decrease in cell activity (Supplementary Fig. S2), indicating that BMP7 affects the development of HCC by regulating phosphorylation of Smad and MAPK.
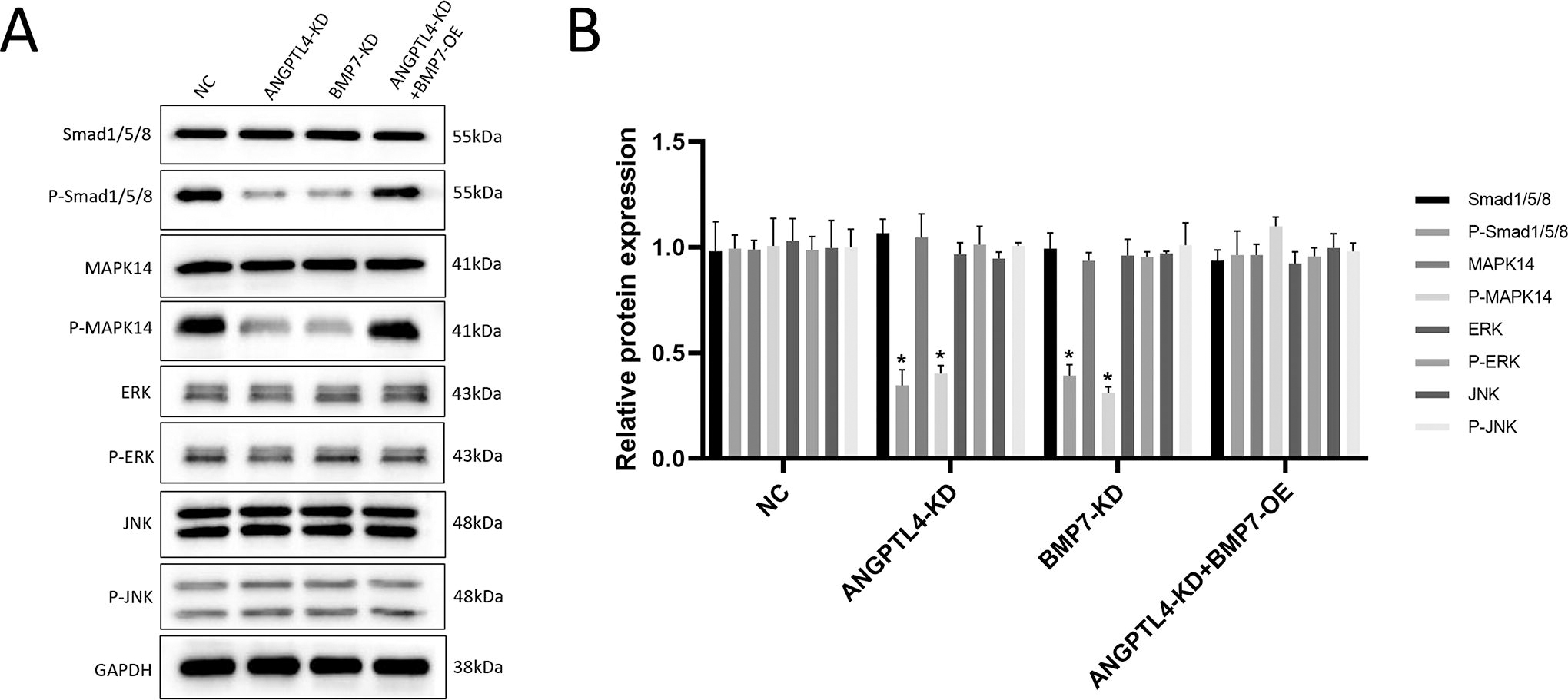
Effects of ANGPTL4 and BMP7 on Smad/MAPK signaling pathways.
ANGPTL4 and BMP7 are essential for tumor growth of HCC
To investigate the effect of ANGPTL4 and BMP7 in tumor growth of HCC in vivo, HepG2 cells with various gene expressions were implanted subcutaneously into mice. The ANGPTL4 and BMP7 knockout significantly decreased the tumor size (Fig. 5A) and tumor weight (Fig. 5B). BMP7 overexpression dramatically enhanced tumor size and weight (Figure 5A and 5B), indicating that ANGPTL4 and BMP7 are critical for tumor growth of HCC.
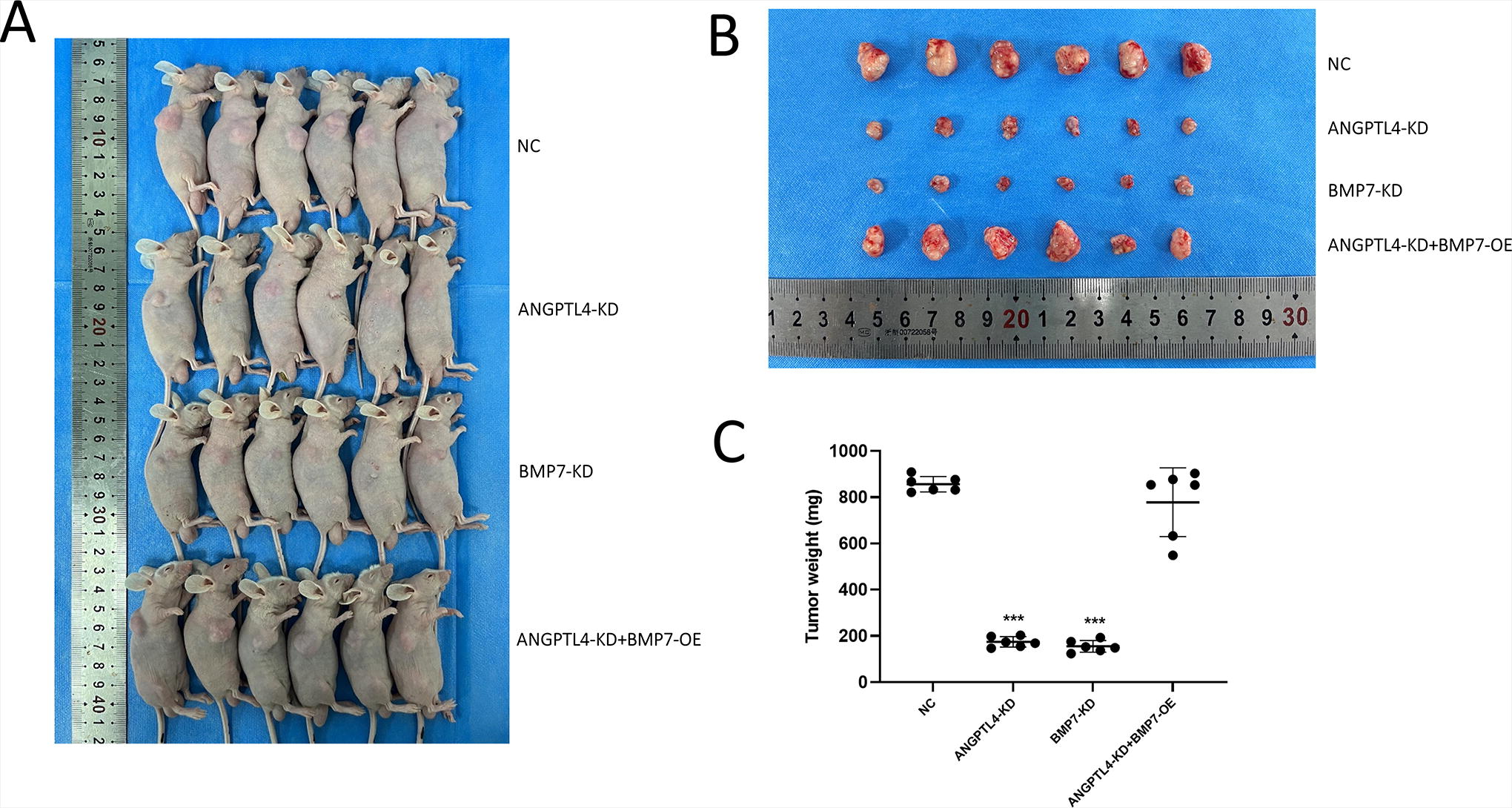
Effects of ANGPTL4 and BMP7 on tumor growth in vivo. (
Discussion
Upregulation of ANGPTL4 promotes tumorigenesis, invasion, drug resistance, and metastasis in human colon, breast, and kidney cancers. Our previous study showed for the first time the role of ANGPTL4 in the development of human HCC and demonstrated that knockdown of ANGPTL4 induced a decrease in tumor cell viability, migration, and invasion and an enhancement of apoptosis (Bai et al., 2024). As an angiopoietin-related protein, ANGPTL4 regulation of cancer cells is associated with increased vascular permeability (Nakayama et al., 2011). Hypoxia is one of the most characteristic features of cancer. Recent studies have elucidated that hypoxia can lead to abnormally elevated levels of ANGPTL4 expression (Kim et al., 2011). In addition, overexpression of ANGPTL4 is positively correlated with the STAT1 signaling pathway in promoting cell proliferation and tumor growth in colorectal cancer (Kim et al., 2011). However, the mechanism of HCC is not well understood. BMP is a member of the transforming growth factor-β (TGF-β) superfamily. About 20 BMP family isozymes have been identified, and these isozymes play a role in different diseases. Among them, BMP7 is aberrantly expressed in many human tumors and is involved in the regulation of cancer cell proliferation, invasion, and migration. However, the role of BMP7 in different types of tumors is not the same. In ovarian cancer studies, BMP7 is highly expressed in advanced ovarian cancer and drug-resistant ovarian cancer cells (Zheng et al., 2021). In gastrointestinal tumors, BMP7 expression is increased significantly more than in normal tissues, and this high expression is associated with poor prognosis and promotion of tumor invasion and metastasis (Fang et al., 2022; Xiao et al., 2022). In lung cancer, BMP7 attenuates tumor cell activity and invasiveness, inhibits bone metastasis, and induces apoptosis and cell cycle arrest. BMP7 inhibits the epithelial–mesenchymal transition-associated genes and cell invasion (Braunschweig et al., 2013; Irimia and Blencowe, 2012), inhibits telomerase, shortens telomeres, and induces senescence and apoptosis in breast cancer cells (Lu et al., 2015). It was also found that BMP7 increased cell proliferation and migration in bone metastatic breast cancer (Wang et al., 2014) and prostate cancer (Coelho and Smith, 2014).
In this study, we demonstrated for the first time that ANGPTL4 is able to interact with BMP7. By constructing ANGPTL4 knockout HepG2 cells, we found that the downregulation of ANGPTL4 expression resulted in a decrease in BMP7 levels. The reversion of BMP7 levels after MG132 treatment of the cells indicated that BMP7 undergoes ubiquitination modification. The ubiquitination level of BMP7 was found to be significantly upregulated on ANGPTL4 knockdown cells by Co-IP assay, suggesting that ANGPTL4 is able to stabilize BMP7 by preventing the degradation that occurs with BMP7 ubiquitination. When ANGPTL4 and BMP7 were knocked down in HepG2 cells, respectively, cell viability and migration ability of the cells were significantly downregulated. However, this downward trend was reversed when BMP7 was supplemented. Current studies on downstream proteins of BMP7-regulated tumors include Smad1/5/8, MAPK14, ERK, and JNK. BMP7 promotes resistance to immunotherapy. BMP7 dose-dependently stimulates proliferation and cadherin-11 expression in mouse renal mesangial cell lines via ERK and p38. BMP7 promotes proliferation of glomerular progenitor cells via a JNK-dependent mechanism. Our results indicate that knockdown of ANGPTL4 and BMP7 had no effect on the expression of ERK, P-ERK, JNK, and P-JNK. However, it was able to downregulate the levels of P-Smad1/5/8 and P-MAPK14. Overexpression of BMP7 in ANGPTL4 knockdown cell lines reversed the downregulation of Smad1/5/8 and MAPK14 phosphorylation levels. Finally, mouse tumor transplantation experiments demonstrated that knockdown of ANGPTL4 and BMP7 significantly inhibited tumor growth in vivo.
In summary, we found for the first time that ANGPTL4 interacts with BMP7, that this binding promotes the stabilization of BMP7 action, and further demonstrated that the regulation of BMP7 by ANGPTL4 alters the expression of a series of proteins downstream of it and that the effect of ANGPTL4 on HCC is achieved through the regulation of BMP7.
Footnotes
Authors’ Contributions
Conceptualization: Y.B. and Y.Y.; Methodology: Y.B., G.C., X.S., and Y.Y.; Validation: Y.B., G.C., and X.S.; Formal analysis: Y.B., M.W., Y.L., J.G., and Y.Y.; Writing—original draft preparation: Y.B. and Y.Y.; Writing—reviewing and editing: G.C. and J.G.
Disclosure Statement
The authors declare no competing interests.
Funding Information
No funding was received for this article.
Supplementary Material
Supplementary Figure S1
Supplementary Figure S2
References
Supplementary Material
Please find the following supplemental material available below.
For Open Access articles published under a Creative Commons License, all supplemental material carries the same license as the article it is associated with.
For non-Open Access articles published, all supplemental material carries a non-exclusive license, and permission requests for re-use of supplemental material or any part of supplemental material shall be sent directly to the copyright owner as specified in the copyright notice associated with the article.