
Abstract
We aim to explore the potential mechanism of bone marrow mesenchymal stem cells–derived extracellular vesicles (BMSCs-Exo) in improving spinal cord injury (SCI). Thirty male 12-week specific pathogen-free (SPF) Sprague–Dawley (SD) rats were used to construct SCI model in vivo. Ten male 12-week SPF SD rats were used to extract BMSCs. The Basso, Beattie, Bresnahan (BBB) score was used to evaluate the motor function of rats. Real-time fluorescence quantitative PCR (RT-PCR), western blot (WB), and double luciferase assay were used to explore the regulation between rno-miR-208a-3p and Cdkn1a (p21) in BMSCs. Primary spinal cord neurons were treated with lipopolysaccharide (100 ng/mL) for 30 min to mimic SCI in vitro. Compared with the model group (14 scores), BMSCs-Exo increased BBB score (19 scores) in SCI rats. Compared with the sham group, Cdkn1a was upregulated, whereas rno-miR-208a-3p was downregulated in the model group. However, compared with the model group, Cdkn1a was downregulated, whereas rno-miR-208a-3p was upregulated in the BMSCs-Exo group. In addition, rno-miR-208a-3p inhibited the expression of Cdkn1a via direct binding way. BMSCs-Exo-rno-miR-208a-3p promoted the proliferation of primary spinal neurons via inhibiting apoptosis in vitro. Moreover, BMSCs-Exo-rno-miR-208a-3p promoted cyclin D1, CDK6, and Bcl-2 and inhibited Bax expression in a cell model of SCI. In conclusion, BMSCs-Exo-carried rno-miR-208a-3p significantly protects rats from SCI via regulating the Cdkn1a pathway.
Introduction
Spinal cord injury (SCI) is recognized as an important global health priority (Maas et al., 2017). SCI not only causes loss of health and disability for patients but also imposes a burden on health care systems and economies (Injury GBDTB, Spinal Cord Injury C, 2019). Many therapeutic strategies have been proposed to develop neuroprotective and neuroregenerative therapies to promote neuronal recovery. Although there have been varying degrees of success, neuronal recovery involves complex mechanisms and remains elusive (Anjum et al., 2020).
Bone marrow mesenchymal stem cells (BMSCs) are pluripotent stem cells with relatively low immunogenicity, which play an active role in SCI repair (Nakazaki et al., 2021). Extracellular vesicles are extracellular particles of 30–150 nm that are produced by almost all cells (Kalluri and LeBleu, 2020). Recently, BMSCs-derived extracellular vesicles (BMSCs-Exo) have been found to have therapeutic clinical implications; for example, BMSCs-Exo promoted regeneration and modulated immune responses (Guo et al., 2019). In addition, BMSCs-Exo could improve cerebral infarction via delivering miR-23a-3p to activate microglia and ameliorate lipopolysaccharide (LPS)-induced acute uterine injury (Dong et al., 2022; Liu et al., 2021).
Extracellular vesicles have the advantage of delivering materials to target cells and exerting regulatory functions. Among substances carried by extracellular vesicles, microRNAs (miRNAs) play an important role in SCI pathology (Pan et al., 2021). Extracellular vesicles secreted by BMSCs modified with miRNA-29b (Yu et al., 2019), miR-338-5p (Zhang et al., 2021), or miRNA-124 (Song et al., 2017) can improve SCI in rats. However, BMSCs-Exo carry a large number of miRNAs, and miRNAs that may improve SCI need to be further explored.
Neuronal recovery after SCI is determined by the interaction between a variety of cells, such as neuronal cells, BMSCs, and immune cells (O’Shea et al., 2017). Based on the above studies, our study sheds light on investigating the important connections between BMSCs-Exo, miRNA, and neurons with the aim of finding key miRNAs, which could be delivered by BMSCs-Exo for improving SCI. In this study, we found BMSCs-Exo-carried rno-miR-208a-3p significantly protects rats from SCI, and this finding might provide new mechanistic insights for the clinical treatment of patients with SCI.
Materials and Methods
Animal
Forty male, 12-week specific pathogen-free Sprague–Dawley (SD) rats (250–300 g) were purchased from Saiye Biotechnology Co., LTD. All SD rats were reared at 23 ± 0.5°C with a light cycle of 12/12 h without pathogens. Research related to animal use complies with all relevant national regulations and policies on animal care and use. All experiments involving animals are approved by the animal Ethics Committee of Shanghai East Hospital, Tongji University School of Medicine (20200026). The National Institutes of Health Guide for the Care and Use of Laboratory Animals was followed in this study. The rats were sacrificed using carbon dioxide (CO2) at a displacement rate of 40% volume/min at the end of the experiment.
Differentially expressed gene analysis
The spinal cord was isolated from SD rats, and the RNA was collected with TRIpure Total RNA Extraction Reagent (ELK Biotechnology). The DNBSEQ platform (BGI) was used to conduct sequencing, and RNA libraries were established by Shenzhen BGI Co., Ltd. as well. Next, the differential expression genes’ (DEGs) analysis among sham, model, and BMSCs-Exo groups was performed using the PossionDis package. DEGs were identified with |log2 (fold change)| > 2 and an adjusted false discovery rate ≤0.001.
Gene signaling transduction research was finished using gene ontology and Kyoto Encyclopedia of Genes and Genomes enrichment analysis (Kanehisa and Goto, 2000).
BMSCs isolation and characterization
Ten SD rats were used to extract primary BMSCs as previously described (Chen et al., 2017). Briefly, SD rats were sacrificed with CO2 and soaked in 75% ethanol for 10 min. Bone marrow was then isolated from the tibias and femurs and subsequently rinsed with minimum essential medium (MEM) alpha medium (A34992L1, Thermo Fisher Scientific) containing 10% fetal bovine serum (FBS, 10091148, Thermo Fisher Scientific) and 1% penicillin–streptomycin (FBS, 15070063, Gibco). The bone marrow was cultured in MEM alpha medium with 10% FBS and 1% penicillin–streptomycin. The cells were cultured at 5% CO2 and 37°C and subcultured when the cells were fused to 80–90%. The number of BMSCs passages in subsequent experiments was <5 times. The third-generation BMSCs were added with CD34 (NB600-1071AF647, Novus), CD44 (743920, BD Pharmingen, China), CD45 (202207, Biolegend), and CD90 (205903, Biolegend) antibodies, respectively, and incubated for 30 min at room temperature under dark conditions. The expression of CD34, CD44, CD45, and CD90 was detected via flow cytometry (FACSCalibur, BD).
Extracellular vesicle extraction from BMSCs
About 1 × 108 BMSCs were cultured in MGro-500 chemically defined serum-free MSC medium (StemRD) for 48 h. BMSCs-Exo were extracted according to the procedure of the cell supernatant extracellular vesicle extraction kit (Ur52121-20t, Shanghai Yumeibo Biotechnology Co., LTD). Briefly, the supernatant of BMSCs was collected and centrifuged at 300 g for 10 min and 2000 g for 10 min at room temperature, respectively. The supernatant was filtered by a 0.22 μm aseptic membrane (LM-439, Shanghai Lianmai Bioengineering Co., LTD) and centrifuged at 100,000 g for 60 min at 4°C. DiR-Membrane EVs Labeling and Purification Kit (EXOPDiR20-1, RenGen Biosciences) was applied to prepare Dir-BMSCs-Exo. Briefly, 50 μL of BMSCs-Exo were incubated with 50 μL Dir labeling dye and reaction buffer (1:10) for 30 min. The prepared Dir-BMSCs-Exo was kept at −80°C and used in the subsequent experiments. The cell culture parameters were based on a previous report (Faezeh et al., 2023).
Nanoparticle tracking analysis
The abovementioned isolated BMSCs-Exo were used for nanoparticle tracking analysis (NTA). In brief, BMSCs-Exo were suspended in 50 μL 1 × phosphate-buffered saline (PBS) buffer and injected into a nanoparticle tracking analyzer (ZetaView PMX 110, Particle Metrix), and the particles of BMSCs-Exo were tracked and determined in terms of Browne motion and diffusion coefficient.
Transmission electron microscopy
The abovementioned isolated BMSCs-Exo were used for transmission electron microscopy (TEM) analysis. In brief, BMSCs-Exo were resuspended in 50 μL 2% paraformaldehyde. BMSCs-Exo suspension (5 μL) was added to the formvar-carbon copper network. Then, the copper mesh adsorbent BMSCs-Exo were suspended on 50 μL 1% glutaraldehyde drops and stained on 4% uranyl acetate drops. Images were captured using a TEM instrument (1230, JEOL).
In vivo SCI model establishment and treatments
Thirty SD rats were randomly selected to construct the SCI model as previously described and divided into three groups equally (Wang et al., 2018), including sham (n = 10), model (n = 10), and BMSCs-Exo groups (n = 10). After spinal exposure in SD rats, we performed laminectomy of the T9 vertebrae. For rats in the model or BMSCs-Exo groups, after full exposure of the spinal cord, we induced a moderate crush injury with a vascular clamp (2 min). Rats in the sham operation group received the same operation without vascular clamp intervention. After the operation, SD rats in the BMSCs-Exo group were intravenously injected with BMSCs-Exo (600 μg/rat) once weekly, and the equivalent dose of normal saline was injected into the sham and model groups.
Basso, Beattie, Bresnahan score
According to the previous description (Willenborg et al., 2021), the motor function of SD rats in each experimental group was scored on days 1, 3, 5, 7, 14, and 28 after SCI surgery. According to the Basso, Beattie, Bresnahan (BBB) scale, a score of 21 is considered completely normal, and a score of 0 is considered completely paralyzed. Motor function is assessed by the coordinated movement of the hips, knees, ankles, toes, and forelimbs. The investigator assessing the BBB scores was blinded to the experimental groups.
Isolation of primary spinal neurons
Primary spinal cord neurons were isolated from SD rat fetuses at 14–15 days (n = 10, Saiye Biotechnology Co., LTD) using the previously described method (Jiang et al., 2006). Briefly, the embryonic spinal canal was opened, and the embryonic spinal cord was cut into small pieces with a scalpel under sterile conditions. The tissue mass was digested through papain (YZ-1495005, Solarbio). The isolated cells were inoculated on a Greiner CELLCOAT® multiwell plate (GN662930, Sigma). After 3 days of culture in neurobasal medium (21103-049, Thermo Fisher Scientific) supplemented with 2% B27 supplement, 5 μM cytosine-β-d-arabinfuran (Sigma-Aldrich) was added to the medium to inhibit nonneuronal cell proliferation. The fresh medium was replaced every 3 days, and the number of cell passages was <3 times.
Establishment of in vitro SCI model
We stimulated primary spinal cord neurons with LPS (100 ng/mL, L8880, Solarbio) to construct SCI cell models in vitro as previously described (Li et al., 2021b; Lv et al., 2019). The primary spinal cord neurons were divided into four groups, including the control, model, BMSCs-Exo-NC, and BMSCs-Exo-inhibitor groups. For the control group, the primary spinal cord neurons were treated with nothing. For the model group, the primary spinal cord neurons were treated with 100 ng/mL LPS. For the BMSCs-Exo-NC group, the primary spinal cord neurons were treated with 100 ng/mL LPS for 30 min.
Dual-luciferase reporter assay
A dual-luciferase reporter assay was conducted to confirm that rno-miR-208a-3p binds to Cdkn1a directly. In brief, wild-type (wt) and mutant (mut) 3′-untranslated region (UTR) of Cdkn1a were cloned into the pmirGLO luciferase reporter vector (RiboBio Co., Ltd.). Next, HEK293T cells were cotransfected with wt-Cdkn1a or mut-Cdkn1a and 50 nM rno-miR-208a-3p mimics or 50 nM mimics-NC using Lipofectamine® 2000 (Thermo Fisher Scientific, Inc.) at 37°C for 48 h. In the end, a dual-luciferase reporter assay system (Promega Corporation) was used to measure the luciferase activity, and the luciferase activity was normalized to Renilla luciferase activity. Rno-miR-208a-3p mimics, inhibitor, and mimics-NC were provided by RiboBio Co., Ltd.
Enzyme-linked immunosorbent assay
The TNF-α and interleukin (IL)-1β levels were measured by enzyme-linked immunosorbent assay (ELISA) using the Rat TNF-α ELISA Kit (Abcam) and the Rat IL-1β ELISA Kit (Abcam) according to the manufacturer’s instructions. Control buffer and serum samples were added into ELISA plates containing TNF-α or IL-1β antibodies and incubated at room temperature for 2 h. After being washed with PBS, the wells were incubated with 100 μL secondary antibody (Horseradish Peroxidase; HRP labeled) for 2 h. After being washed with PBS, the wells were incubated with 100 μL Tetramethylbenzidine without light for 15 min. The absorbance was measured at 450 nm.
Real-time fluorescence quantitative PCR
Total RNA extraction and reverse transcription were performed according to the standard protocol described previously (Fang et al., 2020). The ReverTra Ace qPCR RT Kit (FSQ-101, TOYOBO) was used for reverse transcription. The POWER SYBR GREEN PCR MASTER Kit (4368708, Applied Biosystems) was used for qPCR. All primers used in this study are shown in Table 1. The relative expression was calculated with a 2−ΔΔCt value. GAPDH is used as an internal reference.
All Primers Used in the Present Study
Western blot
RIPA is used to extract proteins from cells or tissues. BCA Protein Assay Kit (Shanghai Sangong Bioengineering Co., LTD.) is used for protein quantification. The primary antibodies used were anti-TNF alpha antibody [EPR21753-109] (1:1000, ab205587, Abcam), anti-IL-1β antibody [EPR23851-127] (1:1000, ab254360, Abcam), anti-p21 antibody [EPR3993] (1:1000, ab109199, Abcam), and anti-GAPDH antibody [6C5]-loading control (1:2000, ab8245, Abcam). HRP-labeled Goat Anti-Rabbit IgG (H + L) (1:2000, A0208, Biyuntian Biotechnology) was used as the secondary antibody. Proteins were detected using an Ultra High Sensitivity ECL Kit (GLPBIO). Images were acquired using Sigmatel software v2.0, and the protein density was calculated using Image Lab software v3.0.
Hematoxylin and eosin staining
The spinal cord sections were collected and deparaffinized. After that, the tissues were stained with a hematoxylin staining solution for 10 min and then counterstained with an eosin staining solution for 1 min at room temperature.
Statistical analysis
GraphPad Prism statistical software (version 8) was used to analyze the data. The measurement data were expressed as mean ± standard deviation (error). The unpaired t-test was used to compare the normal distribution and homogeneity of variance between the two groups of data. One-way analysis of variance (ANOVA) was used to analyze multiple sets of data, and repeated measurement ANOVA was performed for data at different time points, followed by a Tukey’s posttest. p-Value <0.05 was considered statistically significant.
Results
Identification of BMSCs and its derived extracellular vesicles
First, BMSCs were isolated, and the expressions of CD34, CD44, CD45, and CD90 were detected using flow cytometry. The results showed that the expression of CD44 and CD90 was positive, whereas the expression of CD34 and CD45 was negative in the third-generation BMSCs (Fig. 1A). In addition, the NTA assay (Fig. 1B) and TEM assay (Fig. 1C) showed that BMSCs-Exo were a bilayer vesicle of 50–150 nm. The expression of CD9, CD81, and TSG101 in BMSCs-Exo was detected by the western blot (WB) assay (Fig. 1D). Furthermore, the particle (NTA)-to-protein (BCA) ratio was 1.4*10^13/µg, and the concentration of particle is 1.4*1010/mL.
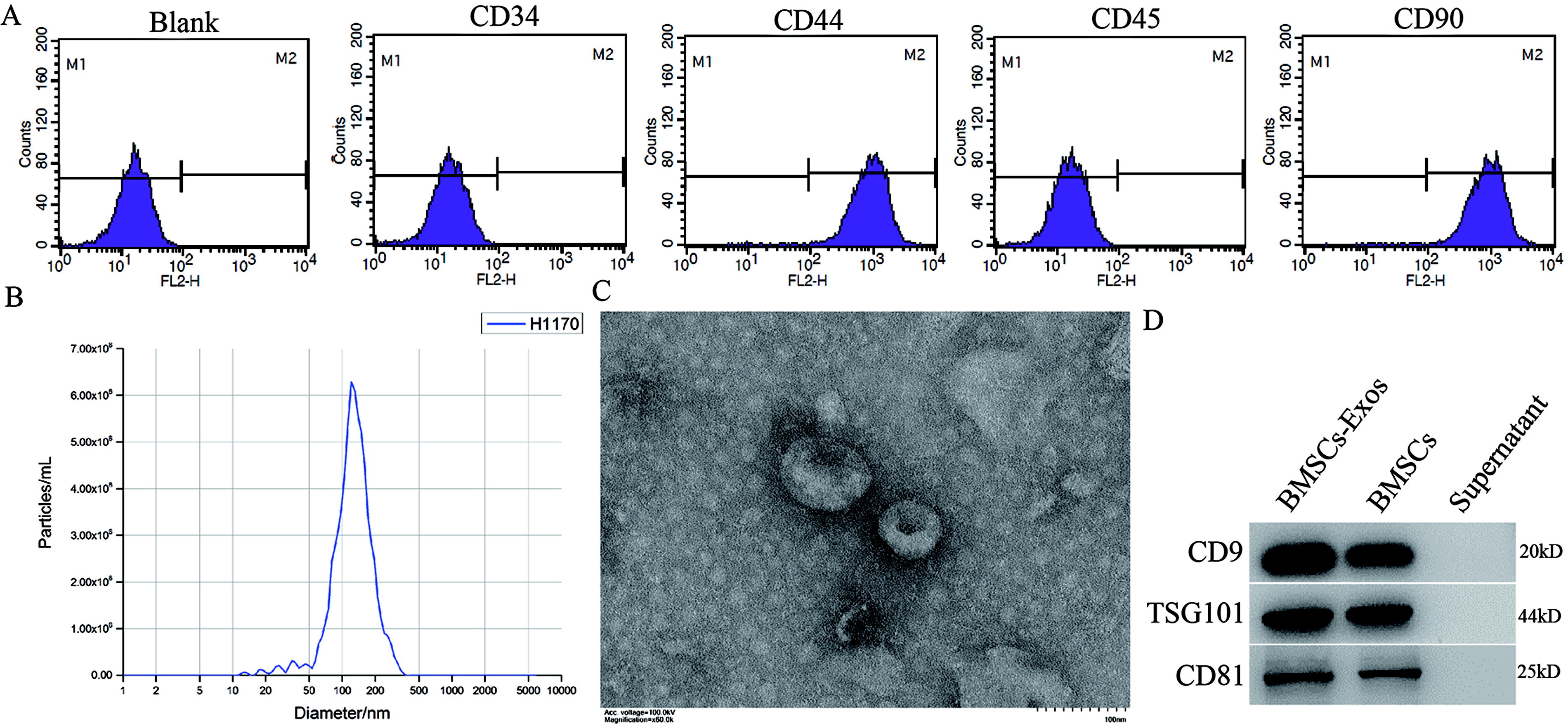
Identification of BMSCs and extracellular vesicle.
BMSCs-Exo improved SCI in vivo
In vivo imaging showed that Dir-labeled BMSCs-Exo reached the injury site of SCI rats 4 h after injection (Fig. 2A). The fluorescence intensity of the model and BMSCs-Exo groups was (5.5 ± 1.8)*106 and (1.4 ± 1.8)*1010 fluorescence units, respectively. Compared with the sham group, BBB scores in the model group were substantially lower on day 5 (3 scores), 7 (6 scores), 14 (8 scores), and 28 (18 scores). Additionally, compared with the model group, BBB scores in the BMSCs-Exo group were substantially higher on day 5 (5 scores), 7 (8 scores), 14 (14 scores), and 28 (19 scores, Fig. 2B). Spinal cord tissues of rats in each group were collected, and hematoxylin and eosin staining results showed that tissue damage was substantially improved after BMSCs-Exo treatment (Fig. 2C). In addition, BMSCs-Exo inhibited the release of SCI-induced proinflammatory cytokines TNF-α and IL-1β (Fig. 2D).
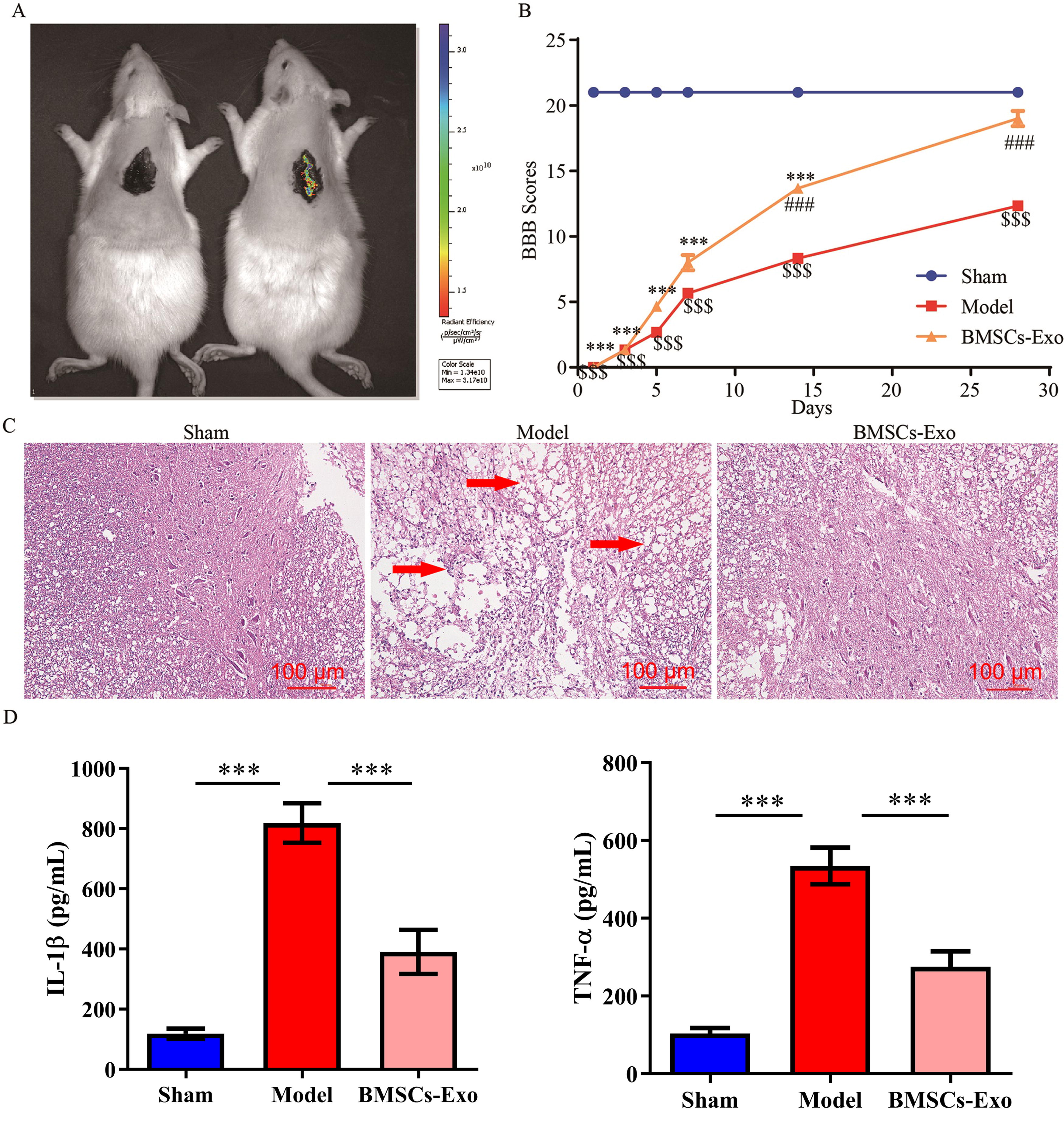
BMSCs-Exo improved SCI in rats.
The potential mechanism of BMSCs-Exo in improving SCI via RNA analysis
Compared with the sham group, 333 DEGs were found upregulated, whereas 787 DEGs were found downregulated in the model group (Fig. 3A). Compared with the model group, 341 DEGs were found upregulated, whereas 360 DEGs were found downregulated in the BMSCs-Exo group (Fig. 3B). Compared with the sham group, 71 DEGs were found upregulated in the model group, and these 71 DEGs were found downregulated in the BMSCs-Exo group compared with the model group (Fig. 3C). Among these 71 DEGs, Wnt2b, Dusp2, Timp3, Hes1, Bdnf, Tgfbr2, Il6r, Abca1, Ctsb, and Cdkn1a were the top 10 DEGs. The mRNA expression of these 10 DEGs in the sham (N = 9), model (N = 9), and BMSCs-Exo (N = 9) groups was detected via RNA analysis (Fig. 3D) and real-time fluorescence quantitative PCR (RT-PCR) assay (Fig. 3E). Cdkn1a (top 1 among the 10 DEGs) was found upregulated in the model group compared with the sham group, whereas Cdkn1a was found downregulated in the BMSCs-Exo group compared with the model group (Fig. 3D and E).

NG sequencing was used to explore the underlying mechanism of BMSCs-Exo in rats with SCI.
Rno-miR-208a-3p inhibited the expression of Cdkn1a via direct binding way
The top 10 miRNA targets of Cdkn1a were predicted via the targetscan database, and the expression of these 10 miRNAs in the sham (N = 9), model (N = 9), and BMSCs-Exo (N = 9) groups was detected via the RT-PCR assay (Fig. 4A). Especially, rno-miR-208a-3p was found downregulated in the model group compared with the sham group, whereas rno-miR-208a-3p (top 1 among the 10 miRNAs) was found upregulated in the BMSCs-Exo group compared with the model group (Fig. 4A). Compared with the sham group, the protein expression of Cdkn1a was upregulated in the model group, whereas, compared with the model group, the protein expression of Cdkn1a was downregulated in the BMSCs-Exo group (Fig. 4B). Compared with the control group, the expression of rno-miR-208a-3p was upregulated in the mimic group (Fig. 4C). Overexpression of rno-miR-208a-3p inhibited the mRNA and protein levels of Cdkn1a in the spinal cord (Fig. 4D and E). In addition, a dual-luciferase reporting assay showed that rno-miR-208a-3p bound to the 3′UTR of Cdkn1a directly (Fig. 4F).
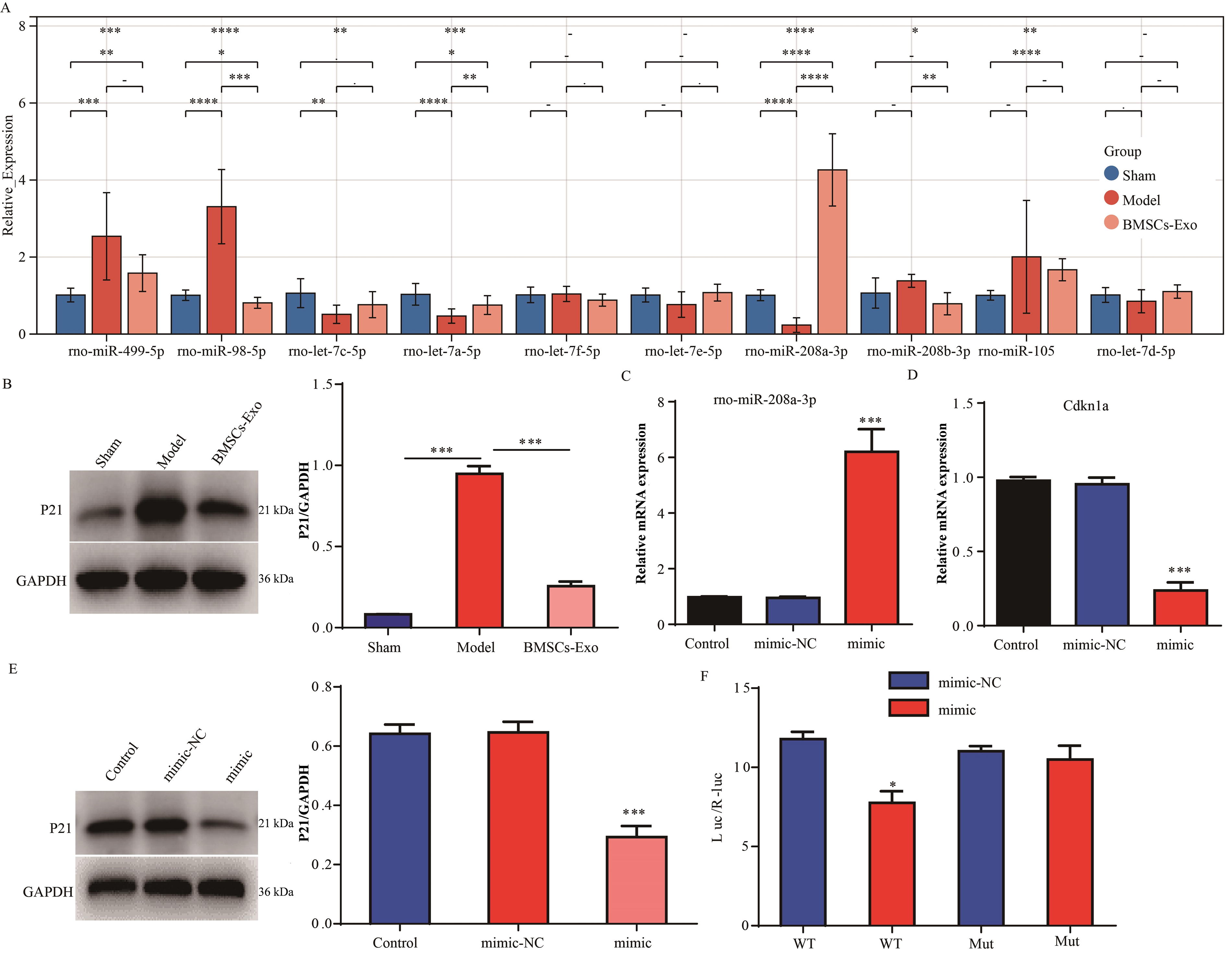
Rno-miR-208a-3p directly bound to Cdkn1a and inhibited its expression.
BMSCs-Exo-rno-miR-208a-3p improved the injury of primary spinal neurons in vitro
Primary spinal neurons were stimulated with LPS to construct the SCI cell model in vitro. Compared with the control group, the cell viability was reduced in the model group via the Cell-Counting-Kit-8 assay (Fig. 5A). Compared with the control group, cell apoptosis was enhanced in the model group via flow cytometry (Fig. 5B). Compared with the control group, the cell proportion of G1 phase was reduced, whereas the cell proportion of S and G2 phase was increased in the model group via flow cytometry (Fig. 5C). The RT-PCR assay showed that the expression of rno-miR-208a-3p was reduced, whereas the expression of Cdkn1a was increased in the model group (Fig. 5D). The WB assay showed that the protein expression of p21 was increased in the model group (Fig. 5E).
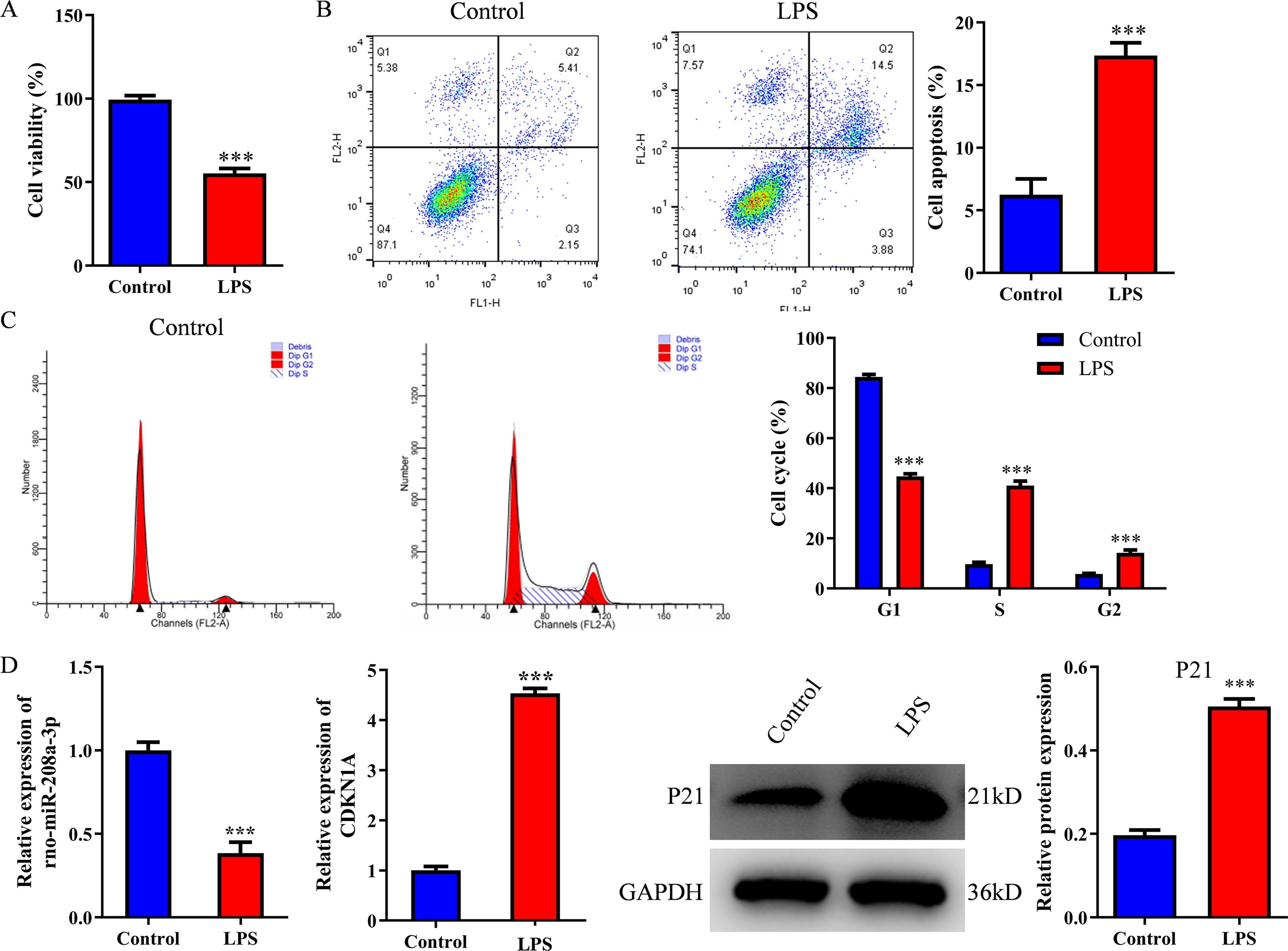
Rno-miR-208a-3p was downregulated, whereas Cdkn1a was upregulated in LPS-induced primary spinal cord cells. Primary spinal cord cells were treated with LPS (100 ng/mL).
RT-PCR was used to detect the expression of rno-miR-208a-3p in BMSCs-Exo treated with rno-miR-208a-3p NC or rno-miR-208a-3p inhibitor (Fig. 6A). In addition, RT-PCR was used to measure the expression of rno-miR-208a-3p and Cdkn1a in primary spinal neurons treated with BMSCs-Exo or/and (Fig. 6B). Compared with the model group, cell viability was increased, whereas cell apoptosis was reduced in the BMSCs-Exo-treated group. Moreover, BMSCs-Exo significantly prevented LPS-induced cell apoptosis in primary spinal neurons, whereas these phenomena were reversed by rno-miR-208a-3p inhibitor (Fig. 6C and D). Consistently, compared with the model group, the cell proportion of G1 phase was increased, whereas the cell proportion of S and G2 phase was reduced in the BMSCs-Exo group. The cell proportion of G1 phase was decreased, whereas the cell proportion of S and G2 phase was increased in the rno-miR-208a-3p inhibitor group (Fig. 6E). The WB assay showed that the expression of TNF-α and IL-1β was upregulated in the model group compared with the control group, whereas this upregulation was prevented by BMSCs-Exo. Similarly, the antiinflammation effect of BMSCs-Exo in primary spinal neurons was abolished in the presence of an rno-miR-208a-3p inhibitor (Fig. 6F and G). Taken together, the antiinflammation effect of BMSCs-Exo was dependent on rno-miR-208a-3p.
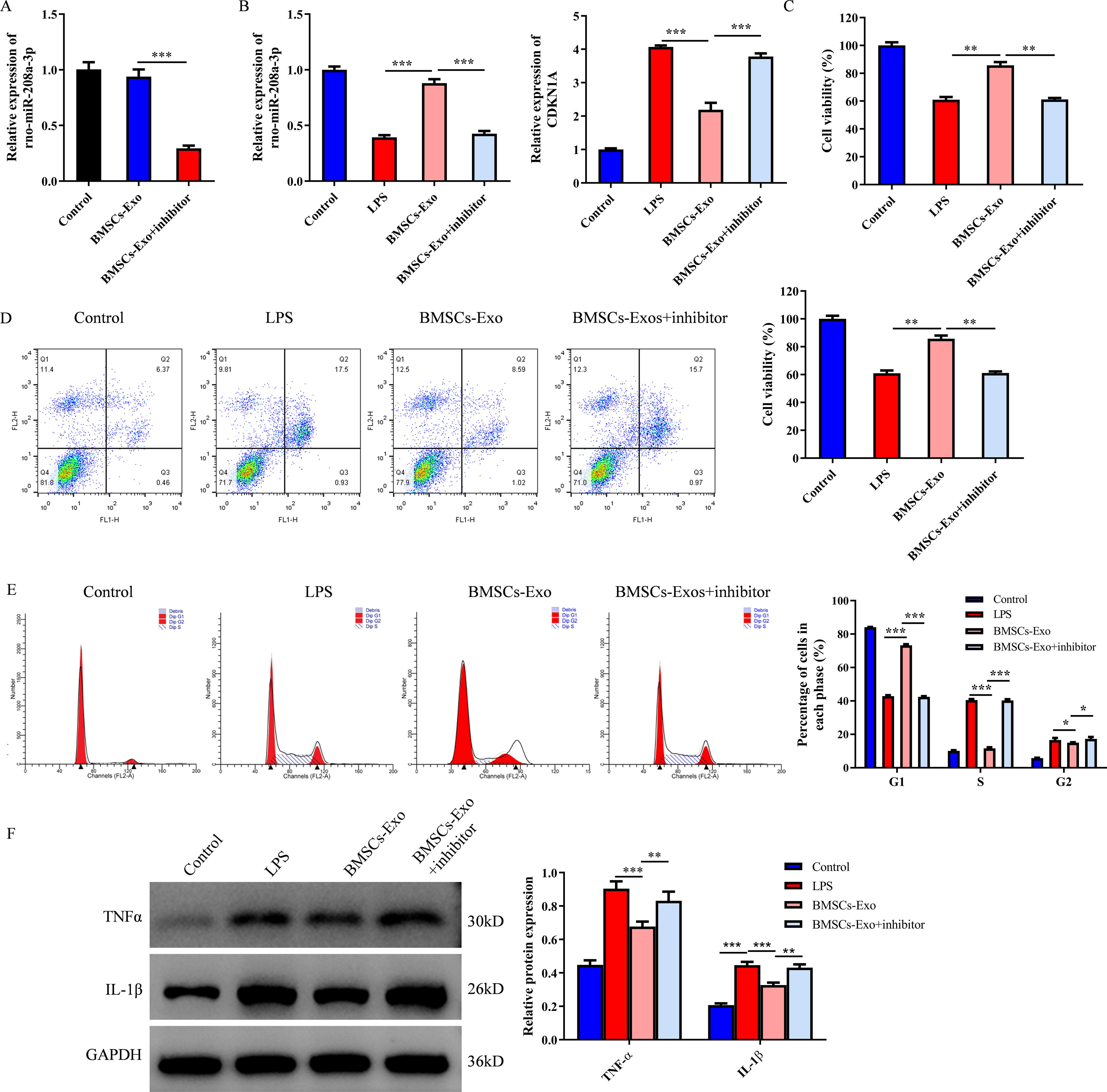
The BMSCs-Exo-carried rno-miR-208a-3p improved the injury of primary spinal neurons in vitro.
Discussion
In recent years, BMSCs-Exos were gradually regarded as promising therapeutic approaches for the treatment of SCI (Sheng et al., 2021; Watanabe et al., 2015). Hence, a more detailed understanding of the underlying mechanisms by which BMSCs-Exo improve SCI has led to the identification of new therapeutic targets. In the present study, we demonstrate that BMSCs-Exo carrying rno-miR-208a-3p could effectively stimulate the primary spinal neurons activity. Our results highlighted that BMSCs-Exo-carried rno-miR-208a-3p significantly protects rats from SCI via regulating the Cdkn1a pathway, and this finding might provide new mechanistic insights into the clinical treatment of patients with SCI.
A key finding of the current study was that BMSCs-Exo increased the BBB score and improved the pathological injury of spinal cord tissues in rats with SCI. Through second-generation sequencing, RT-PCR, and WB assays, we found that the expression of p21 was increased significantly in the spinal cord tissues of rats with SCI compared with sham rats; however, BMSCs-Exo could effectively reverse the upregulation of p21 in the spinal cord tissues of rats with SCI. It is reported that microglial extracellular vesicle miR-124-3p inhibited neuronal apoptosis and the underlying mechanism involved in the inhibition of the p53/p21/CDK1 signaling pathway (Li et al., 2021a). Therefore, we hypothesized that BMSCs-Exo might inhibit p21 expression by carrying some miRNAs.
Based on the bioinformatics analysis, RT-PCR, WB, and a dual-luciferase reporting assay, we found that rno-miR-208a-3p directly bound to p21 and inhibited its expression. In order to elucidate the effect of BMSCs-Exo-loaded rno-miR-208a-3p on neuronal cell function, a series of functional recovery experiments were conducted. Specifically, BMSCs-Exo-NC promoted cell proliferation by inhibiting apoptosis of primary spinal neurons in vitro. Nevertheless, the protective effect of BMSCs-Exo on primary spinal neurons in vitro was abolished in the presence of an rno-miR-208a-3p inhibitor. Therefore, we considered that rno-miR-208a-3p-modified BMSCs-Exo effectively promotes the activity of primary spinal cord neurons.
We also found that the expression of TNF-α and IL-1β was substantially increased in the SCI model group compared with that in the sham group. However, BMSCs-Exo treatment substantially inhibited the upregulation of TNF-α and IL-1β in rats with SCI in vivo. Although post-SCI inflammation has multiple positive effects, extensive infiltration of proinflammatory cytokines is the primary cause of neurodegeneration (Garcia et al., 2016). Consistently, BMSCs-Exo-NC inhibited TNF-α and IL-1β expressions in LPS-treated primary spinal neurons in vitro; however, these effects were abolished in the presence of an rno-miR-208a-3p inhibitor. All these results confirmed that rno-miR-208a-3p may be a vital factor for rno-miR-208a-3p-modified BMSCs-Exo, promoting the activity of primary spinal neuron cells. However, there are some challenges in translating animal model results to human treatments. First, rno-miR-208a-3p and human miR-208a-3p might play different roles in SCI disease. The function of human miR-208a-3p should be confirmed. Second, the direct target of rno-miR-208a-3p is p21, which plays a very important role in cell proliferation. Thus, the side effects of rno-miR-208a-3p should be well investigated. Third, the pharmacokinetic profile of rno-miR-208a-3p should be explored in the future.
In a word, rno-miR-208a-3p-modified BMSCs-Exo promote cell viability and decrease the release of proinflammatory cytokines in primary spinal cord neurons. The miR-208a-3p-modified BMSCs-Exo might be a promising novel strategy for the treatment of patients with SCI.
Footnotes
Authors’ Contributions
J.Y. and Y.Y. participated in performing the experiments and writing the paper. J.Y. supervised the study and revised the paper.
Data Availability
The datasets analyzed during the current study are available from the corresponding author upon reasonable request.
Disclosure Statement
The authors declare that they have no competing interests.